

Федеральное государственное образовательное учреждение
высшего профессионального образования
«Московская государственная академия
ветеринарной медицины и биотехнологии имени К. И. Скрябина»
| УДК 544.7 № госрегистрации 01200961265 Инв.№ | УТВЕРЖДАЮ Ректор ФГОУ ВПО МГАВМиБ, академик РАСХН ______________Ф.И. Василевич «___» ___________ 2009 г. |
ОТЧЕТ О НАУЧНО-ИССЛЕДОВАТЕЛЬСКОЙ РАБОТЕ
Проведение научных исследований коллективами научно-образовательных центров в области коллоидной химии и поверхностных явлений.
по теме:
РАЗРАБОТКА МЕТОДИК И СОЗДАНИЕ БИОХИМИЧЕСКИХ КОЛЛОИДНЫХ СИСТЕМ ДЛЯ ВЕТЕРИНАРНО-БИОЛОГИЧЕСКИХ И ЗООТЕХНИЧЕСКИХ НАПРАВЛЕНИЙ
(промежуточный)
Этап первый:
«Выбор направлений исследований и этап предварительных исследований по ферментным коллоидным системам»
Государственный контракт от «07» июля 2009 г. № 02.740.11.0270 в рамках ФЦП «Научные и научно-педагогические кадры инновационной России» на 2009-2013 годы
Мероприятие 1.1 Проведение научных исследований коллективами научно-образовательных центров.
| Руководитель темы | _____________________ (подпись) « __»___________2009г. (дата) | /Балакирев Н.А./ |
Москва 2009
Список основных исполнителей
| Проректор ФГОУ ВПО МГАВМиБ, академик РАСХН | ___________________ подпись, дата | Н.А. Балакирев |
| Зав. кафедрой, д.х.н., д.б.н., проф | ___________________ подпись, дата | С. Ю. Зайцев |
| Проф., д.х.н | ___________________ подпись, дата | М. С. Царькова |
| Доцент, к.б.н. | ___________________ подпись, дата | Т.В. Каштиго |
| Доцент, к.х.н. | ___________________ подпись, дата | О.С. Белоновская |
| Доцент, к.б.н. | ___________________ подпись, дата | А.А. Лисицына |
| Докторант, к.х.н. | ___________________ подпись, дата | И.Н. Староверова |
| Ст. преподаватель, к.б.н. | ___________________ подпись, дата | Т.О. Азарнова |
| Зав. уч. лабораторией, к.б.н. | ___________________ подпись, дата | И.В. Милаёва |
| Аспирант | ___________________ подпись, дата | Е. Н. Зарудная |
| Аспирант | ___________________ подпись, дата | В. В. Бондаренко |
| Аспирант | ___________________ подпись, дата | А. Н. Тимонин |
| Студент | ___________________ подпись, дата | Д.О. Соловьева |
| Лаборант | ___________________ подпись, дата | М.Н. Шапошников |
Реферат
Отчет 150 с., 6 ч., 31 рис., 10 табл., 121 источник.
Ключевые слова: биохимические коллоидные системы, иммобилизованные ферменты, супрамолекулярные ферментные комплексы, ферментные коллоидные системы, коллоидно-химические свойства, белки, полиэлектролиты.
Объекты исследования и разработки. Анализ научно-технической литературы и других материалов, относящихся к биохимическим коллоидным системам, ферментным комплексам и иммобилизованным ферментам, патентные исследования. Получение комплексов липаз с пептидами, изучение каталитической активности таких комплексов в зависимости от условий. Синтез полимерного антистатического средства для текстильных материалов и меха, исследование его термостабильности, антистатических свойств и адгезионных характеристик.
Цели работы по первому этапу
- Анализ научно-технической литературы и других материалов, относящихся к ферментным коллоидным системам (ФКС). Информационно-аналитическое обеспечение исследований ФКС.
- Патентные исследования по ФКС.
- Описание постановки и научно-техническая реализация проведения исследований ФКС.
- Изучение каталитических свойств липаз в комплексе с белками и пептидами.
- Разработка способа получения полимерного антистатического средства и изучение его коллоидно-химических свойств.
- Анализ и обобщение полученных на 1 этапе результатов НИР.
Метод и методология проведения работы включали: анализ научно-технической литературы и других материалов путём изучения печатных и электронных, отечественных и зарубежных изданий, относящихся к ФКС, патентный поиск по международной базе данных, описание постановки и технической реализации проведения исследований, разработка сетевого графика, проведение экспериментальных работ по получению комплексов ферментов (на примере липаз из разных источников) с белками и пептидами, изучение каталитической активности таких комплексов при разном соотношении полимер : липаза. Синтез полимерного антистатического средства на основе сополимеров N,N’-диметил-N,N’-диаллиламионий хлорида и метилметакрилата (ММА), отличающегося большой адгезивной способностью к текстильным материалам и образующего пленкообразные покрытия с высокими антистатическими и термическими свойствами.
В работе использованы следующие физико-химические методы исследования:
потенциометрическое титрование, тензиометрия, ЯМР-спектроскопия, термогравиметрический анализ.
Результаты работы
1. Проведён анализ научно-технической литературы и других материалов, относящихся к ФКС. Изучены методы получения ФКС, их свойства, применение в биологии, медицине, технологии живых систем.
2. Проведенные патентные исследования выявили патентную чистоту разрабатываемых в рамках данного проекта ФКС.
3. Разработана и описана постановка и техническая реализация проведения исследований по ФКС.
- Изучены каталитические свойства липаз из разных источников (из поджелудочной железы свиньи, бактерий Pseudomonas fluorescence, гриба Mucor javanicus) в комплексе с белками и пептидами.
- Разработан способ получения полимерного антистатического средства и изучены его коллоидно-химические свойства.
- Проведен анализ и обобщены полученные на 1 этапе результаты НИР.
Рекомендации по внедрению результатов НИР
Разработка методик и создание биохимических коллоидных систем с использованием современных методических и инструментальных достижений позволит решить ряд актуальных фундаментальных проблем в области коллоидной химии и поверхностных явлений, а также прикладных задач биомедицины и экологии, нано- и биотехнологии, ветеринарии и зоотехнии. Биохимические коллоидные системы на основе ферментов позволят выявить молекулярные механизмы каталитической активности и расширить ассортимент и области применения ферментных препаратов в производстве моющих средств, поверхностно- и биологически-активных соединений, вкусовых и ароматических компонентов, для аналитических и лекарственных целей в медицине и ветеринарии. Особенное значение приобретает разработка и внедрение более эффективных средств диагностики и мониторинга состояния организма животных, что является неотъемлемой частью научно-технического прогресса в развитии животноводства на современном этапе и будет реализовано в данной НИР на базе достижений коллоидной химии.
Выполнение НИР обеспечит достижение научных результатов мирового уровня в решении многих актуальных фундаментальных и прикладных проблем химии и биологии, медицины и экологии, нано- и биотехнологии, ветеринарии и зоотехнии; завершить создание комплексной системы подготовки высококвалифицированных специалистов (от бакалавров и магистров до кандидатов и докторов наук), обладающих особыми компетенциями по профилю данной НИР, что приведет к формированию эффективного и жизнеспособного научного коллектива.
Область применения полученных результатов
Результаты проекта должны быть включены в образовательную программу ФГОУ ВПО МГАВМиБ и использоваться в образовательном процессе на базе научного кадрового потенциала, лабораторного и испытательного оборудования научно-образовательного центра ФГОУ ВПО МГАВМиБ, в том числе в процессе проведения настоящих научно-технических работ.
Новые ФКС, разработанные на основе липазы, могут найти применение в биотехнологии, пищевой промышленности, производстве кормов и т.д. Разработанные способы получения антистатических средств будут использованы в текстильной промышленности. Проведенные исследования новых ФКС на основе липаз из поджелудочной железы свиньи, бактерий Pseudomonas fluorescence, гриба Mucor javanicus исключительно важны для дальнейшей работы по оптимизации каталитической активности этих ферментов с учётом физико-химических параметров и выбора полиэлектролита. Полученные на данном этапе результаты являются ключевыми для успешного выполнения всего проекта.
Прогнозные предположения о развитии объекта исследования
В процессе проведения НИР должны быть разработаны технологические параметры ФКС и методические рекомендации по их использованию для различных областей применения, в том числе: экспериментальные образцы; методики испытаний экспериментальных образцов; технологии получения биохимических коллоидных систем.
В ходе выполнения этапа НИР будут получены результаты интеллектуальной деятельности (статьи, патенты и т.д.); учебно-методические комплексы, включающие учебно-методические пособия, указания и рекомендации; примерные и рабочие учебные программы, практикумы по дисциплинам «Биохимия мембран», «Физическая и коллоидная химия», «Кинетика и термодинамика ферментативных реакций», «Энзимология», «Биоэнергетика», «Бионанотехнология» и другие.
СОДЕРЖАНИЕ
| Реферат ……………………………………………………………………………………….…… | 3 |
| Введение ……………………………..………………………………………………………….… | 10 |
| 1.1 Анализ научно-технической литературы и других материалов, относящихся к ферментным коллоидным системам (ФКС). Информационно-аналитическое обеспечение исследований ФКС | 12 |
| 1.1.1 Ферменты – коллоидные частицы ………………………………………………………..… | 12 |
| 1.1.2 Типы наночастиц ……………………………………….…………………………………... | 13 |
| 1.1.3 Супрамолекулярные комплексы ферментов …………………………………………….... | 16 |
| 1.1.4 Общие принципы иммобилизации ферментов и свойства супрамолекулярных ферментативных систем ………………………………………………………………..….……… 1.1.5 Иммобилизированные ферменты – устойчивые системы………………………………… 1.1.6 Применение иммобилизированных ферментов……………………………………………. 1.1.7 ФКС – диагностические тест-системы……………………………………………………… 1.1.8 Инкапсулирование ферментов……………………………………………………………… 1.1.9 Использование иммобилизованных ферментов в терапии………………………………... 1.1.10 Другие области применения ФКС…………………………………………………………. 1.1.11 Нанокапсулированные ферментные системы…………………………………... 1.1.12 Информационно-аналитическое обеспечение исследований ФКС……………………… | 39 51 53 61 63 66 72 75 85 |
| 1.2 Проведение патентных исследований по ФКС | 97 |
| 1.3 Описание постановки и научно-техническая реализация проведения исследований ФКС ………………………………………………………………………………………………… | 109 |
| 1.3.1 Определение активности липазы методом потенциометрического титрования………… 1.3.2 Определение динамического поверхностного натяжения (ДПН) ферментных систем…. 1.3.3 Исследование термических и термоокислительных свойств новых сополимеров и антистатических свойств обработанных ими материалов………………………………………. 1.3.4 Статистическая обработка результатов…………………………………………………….. 1.3.5 Планирование разработки с использованием сетевого графика………………………….. 1.4 Изучение каталитических свойств липаз в комплексе с белками и пептидами…...… 1.4.1 Определение каталитической активности липаз из различных источников в присутствии природного полимера – бычьего сывороточного альбумина (БСА)…………….. 1.4.2 Измерение ДПН растворов липазы из поджелудочной железы свиньи в смеси с бычьим сывороточным альбумином……………………………………………………………… 1.4.3 Измерение ДПН растворов липазы из поджелудочной железы свиньи в смеси с поли-L-глутаминовой кислотой………………………………………………………………………… 1.4.4 Измерение ДПН растворов липазы из поджелудочной железы свиньи в смеси с поли-L-лизином………………………………………………………………………………………….. 1.4.5 Измерение ДПН растворов липазы из поджелудочной железы свиньи в трехкомпонентном комплексе с поли-L-лизином и поли-L-глутаминовой кислотой…..…… | 111 112 115 119 120 126 126 127 129 130 131 |
| 1.5 Разработка способа получения полимерного антистатического средства и изучение его коллоидно-химических свойств…………………………………………………………… | 132 |
| 1.6 Анализ и обобщение полученных на 1 этапе результатов НИР …………….………... | 139 |
| Список использованных источников…………………………………………………………. | 141 |
ОБОЗНАЧЕНИЯ И СОКРАЩЕНИЯ
АЛТ Аланинаминотрансфераза
АМ Антимикробные полимеры
6-АПК 6-аминопенициллановая кислота
АСТ Аспартатаминотрансфераза
БАС Биологически активное соединение
ГГТ Гамма-глутамилтрансфераза
ГОД Глюкозооксидаза
ИЖ Ионы железа
ИМ Иммозим
ИН Инулиназа
ИП Иммобилизованная пектиназа
ИПЕ Иммобилизованная пектиназа
КФ Классификация ферментов
КК Креатинкиназа
ОПА Общая протеолитическая активность
ЛДГ Лактатдегидрогеназа
ПАА Поли(аллиламин)
ПААГ Полиакриламидный гель
ПАВ Поверхностно-активные вещества
ПАМАМ Полиамидоамин
ПВС Поливиниловый спирт
ПДДА Поли(диметилдиаллиламмоний хлорид)
ПЕ Пектиназа
ПНМК Полиэлектролитные нано- и микрокапсулы
ПР Пектиновый раствор
ПСС Поли(стиролсульфонат)
ПЭ Полиэлектролит
ПЭГ Полиэтиленгликоль
ПЭИ Поли(этиленимин)
ПЭНП Полиэтилен низкой плотности
ПФ Пиридоксальфосфат
СБ Селективные барьерные свойства
СБС Супрамолекулярная биологическая система
СМС Синтетические моющие средства
СНС Супрамолекулярные наноразмерные системы
ФКС Ферментные коллоидные системы
ЭДТА Этилендиаминтетрауксусная кислота
Введение
Создание супрамолекулярных биохимических систем (СБС) на основе таких биоорганических соединений, как белки, пептиды, липиды и их синтетических производных, является одной из наиболее интересных и активно развивающихся в настоящее время областей исследований, находящаяся «на стыке» органической и биологической химии, полимерной и коллоидной химии, физической и аналитической химии, ряда современных направлений биологических и медицинских наук. По определению одного из основоположников супрамолекулярной химии и лауреата Нобелевской премии по химии Жан-Мари Лена «супрамолекулярная химия – это химия межмолекулярных связей, изучающая ассоциацию двух и более химических частиц, а также строение и свойства подобных ассоциаций». Наиболее наглядными примерами такого рода СБС могут служить мембранные системы, широко используемые для моделирования структуры и функции биомембран, а именно: мономолекулярные слои (монослои) и пленки Ленгмюра-Блоджетт, плоские бислойные липидные мембраны (БЛМ), сферические би- и полислойные мембраны (везикулы, липосомы и аналоги). Главным недостатком указанных систем является их невысокая стабильность в отличие от природных мембран, стабилизированных электростатическими и гидрофобными взаимодействиями интегральных и периферических белков, пептидов, липидов и гликолипидов. Одним из наиболее успешных подходов к решению этой важной проблемы, развиваемых и автором с сотрудниками, является создание полимерных монослоев, БЛМ и липосом на основе липидоподобных и поверхностно-активных мономеров.
Последние достижения в области СБС связаны с зарождением и бурным развитием нанотехнологии, в том числе – биомедицинской нанотехнологии (биомеднанотехнологии), бионанотехнологии или нанобиотехнологии, которые многими авторами рассматриваются как синонимы. Одно из возможных определений бионанотехнологии (биомеднанотехнологии) может быть представлено как особая область биохимических и биомедицинских наук, занимающаяся изучением принципов создания и функционирования биологических наноразмерных систем; применением полученных знаний для усовершенствования существующих и создания принципиально новых бионаноматериалов, биотехнологических процессов и бионаноразмерных устройств; новых нанотехнологий для локальной селективной диагностики, терапии, хирургии, генной инженерии и биотехнологии. Активное развитие этой области связано с крупными достижениями в биохимии и молекулярной биологии клетки, в познании механизмов действия клеточных и субклеточных систем, а также – с развитием новых методов исследования наноразмерных биосистем, физиолого-биохимических показателей в организме животных и человека. В простейшем случае, даже отдельные сложные белки (хромопротеины, металопротеины, липопротеины, гликопротеины и т.д.), можно рассматривать как природные наносистемы, структурно и функционально оптимизированные в процессе эволюции. Тем более, это относится к их супрамолекулярным системам (агрегатам белков и т.д.), образующимся путем фундаментальных принципов самоассоциации и молекулярного узнавания.
1.1 Анализ научно-технической литературы и других материалов, относящихся к ферментным коллоидным системам (ФКС). Информационно-аналитическое обеспечение исследований ФКС.
Отв. исполнители: зав. кафедрой ФГОУ ВПО МГАВМиБ, проф. д.х.н, д.б.н. С.Ю. Зайцев; доцент, к.б.н. О.С. Белоновская.
1.1.1 Ферменты – коллоидные системы
Глобальная роль коллоидов в естествознании заключается в том, что они являются основными компонентами таких биологических образований как живые организмы. Все биополимеры организма человека и животных представляют собой коллоидные системы.
Практически любая жидкость или ткань организма человека представляет собой коллоидно-дисперсную среду. Таковыми являются, например, кровь, лимфа, молоко содержимое желудочно-кишечного тракта, желчь, спинномозговая жидкость, моча. Веществами, имеющими размеры и обладающими свойствами коллоидных частиц в живом организме, являются белки (в том числе, ферменты), полисахариды, нуклеиновые кислоты.
Ферменты — это белки, т.е. относительно лабильные полипептидные соединения природного происхождения. Молекулы ферментов содержат уникальные структуры, возникающие в результате упорядоченного свертывания полипептидной цепочки, которое происходит при синтезе фермента. В нем появляется сравнительно ограниченная область, состоящая из реакционноспособных химических групп и исполняющая каталитические функции. Геометрическое строение такой области, называемой активным центром, исключительно точно «настроено» на размеры и химические особенности молекул тех веществ, реакции которых катализирует данный фермент. Благодаря этому ферменты обладают чрезвычайно высокой каталитической активностью и специфичностью. Например, химотрипсин, сравнительно малоактивный фермент, в желудочно -кишечном тракте в реакциях гидролиза пептидов и сложных эфиров в 107 раз активнее, чем ион гидроксила.
Специфичность ферментов весьма высока, нередки случаи, когда она абсолютна. Последнее означает, что данный фермент способен катализировать превращение одного-единственного вещества из многих сотен или тысяч веществ близкой химической природы. Известны, однако, ферменты относительно слабой специфичности. Все эти особенности способствуют использованию ферментов в практике, так как в каждом конкретном случае удается выбрать фермент нужной специфичности.
Широкому применению ферментов в различных областях химии, включая ее прикладные направления, до последнего времени мешали три обстоятельства: относительно высокая стоимость ферментов; их высокая лабильность; трудность отделения от продуктов химических реакций для повторного использования.
Развитие микробиологических методов получения ферментов практически полностью решило первую проблему. Сегодня микроорганизмы стали источником производства большого числа ферментов как для научных, так и для технических целей. Можно высказать твердую уверенность, что с дальнейшим совершенствованием микробиологической технологии цена ферментов будет неуклонно понижаться. [1]
1.1.2 Типы наночастиц
«Нано» (греч. – миллиардная доля) в применении к описываемым объектам подразумевает, что их размеры находятся в пределах 1500 нм (от 109до 510-7м), что соответствует уровням биологической организации от атомарного до субклеточного. Таким образом, под определение «наночастицы», вообще-то, попадают практически любые супрамолекулярные комплексы. Однако, по уже сложившейся традиции в биологической и медицинской литературе, под наночастицами обычно подразумевают вполне конкретные молекулярные конструкции.
Наночастицы можно условно разделить на несколько классов (рис. 1).

Биологические и биогенные наночастицы. Биологический мир буквально наполнен наночастицами – это ферменты (белки с каталитической активностью), молекулы ДНК и РНК, рибосомы, клеточные везикулы, вирусы и пр. Отличительной особенностью таких объектов является их способность к агрегации и самоорганизации. Это свойство активно используется при создании искусственных конструкций, имитирующих реальные биологические структуры. Яркий пример представляют собой различные однокомпонентные и мультикомпонентные липосомы, которые способны при определенных условиях формироваться из раствора смеси липидов. Часто на практике используют и уже существующие в природе биологические наночастицы. Например, различные вирусы активно применяют для генной модификации (трансфекции) клеток. Показано, что аденовирусы с подавленной системой репликации могут быть эффективно использованы и для местной неинвазивной вакцинации через кожу (доставке антигенов к клеткам Лангерганса, присутствующим в коже) [2]. К искусственным биогенным наночастицам, предназначенным для направленной доставки, помимо липосом обычно также относят липидные нанотрубки [16], липидные наночастицы и наноэмульсии, циклические пептиды [4], хитозаны, наночастицы на основе нуклеиновых кислот [5].
Полимерные наночастицы. Полимерные материалы обладают рядом преимуществ, определяющих эффективность их применения в технологиях доставки, – биосовместимость, способность к биодеградации, функциональная совместимость. Типичными соединениями, которые представляют основу для создания полимерных наночастиц, являются полимолочная и полигликолевая кислоты, полиэтиленгликоль (ПЭГ), поликапралактон и др., а также их различные сополимеры. ПЭГ часто используют для повышения стабильности различных молекулярных переносчиков. Например, липосомы, покрытые ПЭГ («стелс-липосомы»), по сравнению с обычными, менее подвержены биодеградации, в результате чего обладают заметным пролонгированным действием [6].
Дендримеры. Дендримеры являются уникальным классом полимеров с сильно разветвлённой структурой. При этом их размер и форма могут быть очень точно заданы при химическом синтезе [7]. Дендримеры получают из мономеров, проводя последовательные конвергентную и дивергентную полимеризации (в том числе используя методы пептидного синтеза). Типичными «мономерами», используемыми в синтезе дендримеров, являются полиамидоамин (ПАМАМ) и аминокислота лизин. «Целевые» молекулы связываются с дендримерами либо путём образования комплексов с их поверхностью, либо встраиваясь глубоко между их отдельными цепями. Контролируемые размеры и свойства поверхности, а также стабильность дендримеров делают их весьма перспективными для использования в качестве переносчиков. На животных моделях показана эффективность их применения для трансдермальной доставки ряда препаратов [8].
Углеродные наночастицы. Нанотрубки и фуллерены являются одними из самых «узнаваемых» наноструктур – практически ни один популярный текст про нанотехнологии не обходится без их изображений. За открытие этой новой формы существования углерода Р. Керл, Р. Смолли и Г. Крото в 1996 г. были удостоены Нобелевской премии по химии. Эти структуры, образованные только атомами углерода, могут быть получены при помощи вольтовой дуги, лазерной абляцией (выжиганием), химическим осаждением из газовой фазы, а также в процессе горения. Сегодня в промышленных масштабах фуллерены получают термическим распылением углеродсодержащей сажи в атмосфере инертного газа при пониженном давлении в присутствии катализатора. Нанотрубки обладают повышенным сродством к липидным структурам; при этом они способны образовывать стабильные комплексы с пептидами и ДНК-олигонуклеотидами [9, 10], и даже инкапсулировать эти молекулы [11, 12]. Это определяет их применение в области создания эффективных систем доставки вакцин и генетического материала [13].
Неорганические наночастицы. К этому классу обычно относят наноструктуры, полученные на основании оксида кремния, а также различных металлов (золото, серебро, платина). При этом часто такая наночастица имеет кремниевое ядро и внешнюю оболочку, сформированную атомами металла. Использование металлов позволяет создавать переносчики, обладающие рядом уникальных свойств. Так, их активность (и в частности, высвобождение терапевтического агента) может быть модулирована термическим воздействием (инфракрасное излучение), а также изменением магнитного поля [13]. При этом показано, что металлические наночастицы могут эффективно проникать вглубь эпидермиса [14].
Использование описанных выше наночастиц в медицине позволит не только эффективно доставлять биологически активные молекулы сквозь различные барьеры организма, которые они не способны преодолевать самостоятельно (кожный, гематоэнцефалический), но и существенно изменять характер действия препарата. Например, трансдермальная доставка, по сравнению с доставкой через кровяное русло, позволяет избежать нежелательных побочных эффектов, снизить эффективную дозу препарата за счет существенного повышения его локальной концентрации. Кроме того, было показано, что у терапевтических молекул, доставляемых в организм с помощью наночастиц, меняется фармакокинетика. Если для препаратов, попадающих в организм перорально или в результате инъекции, увеличение концентрации во времени описывается характерной кинетической кривой первого порядка (концентрация экспоненциально увеличивается во времени), то в случае использования наночастиц наблюдается идеальная временная зависимость нулевого порядка (равномерное увеличение концентрации препарата во времени) [13]. Это позволяет более точно планировать дозировки препарата и пролонгировать его действие.
1.1.3 Супрамолекулярные комплексы ферментов
Хорошо известно, что существование организма животного неразрывно связано с множеством биохимических реакций клеточного метаболизма. Эти реакции протекают в присутствии специфических белков – ферментов, наделенных каталитическими свойствами [14–16]. Имеющаяся обширная и доступная литература по ферментам позволяет не останавливаться на определении классов и всех характеристик этих специфических белков [14–16] (в списке литературы к данной диссертации цитируются только некоторые издания, необходимые для этой работы). В данном разделе литературного обзора основное внимание будет уделено структурно-функциональной характеристике только некоторых ферментов (наиболее важных для нашего исследования) из их огромного множества, существующего в природе. На сегодня выделено более 3500 ферментов, для нескольких сотен из которых уже определена трехмерная пространственная структура [14–16].
Наиболее полной и широко известной является классификация ферментов (КФ) по типу катализируемой биохимической реакции [16]. Эта «функциональная» классификация, принятая в Москве на Международном биохимическом съезде в 1961 году, выделяет 6 классов ферментов: оксидоредуктазы; трансферазы; гидролазы; лиазы; изомеразы; лигазы (синтетазы) [14–16]. В последнее время активно развивается классификация ферментов по уровням структурной организации. На взгляд автора, наиболее рациональным и удачным для «структурной» классификации ферментов является использование принципов и подходов, развитых недавно в супрамолекулярной и биоорганической химии [15, 17].
В современной биохимической литературе это иллюстрируется таблицей 1, переведенной из монографии П. Фриедриха (P. Friedrich) [18].

Начиная с работ С.И. Афонского [20], представления о полисубъединичных ферментах и ферментных комплексах как супрамолекулярных системах начинают использоваться и в нашей биохимической литературе. Многие ферменты, определяемые в плазме крови и широко применяемые для диагностики физиолого-биохимического статуса человека и животных, можно отнести к супрамолекулярным ферментным системам, поскольку in vivo: 1) они образуют достаточно сложные комплексы с субстратами и модификаторами различных типов; 2) они состоят из нескольких полипептидных цепей (субъединиц), которые, комбинируясь различными способами, образуют четвертичную структуру фермента; 3) они имеют четко определенный набор изоферментных форм. Последняя характеристика имеет уникальное значение для клинической диагностики патологических состояний определенных органов и тканей как человека, так и животных.
Принято считать, что изоферменты – это группа ферментов, которая катализирует один и тот же тип реакции, но имеет различные физико-химические характеристики [16]. Например, изоферменты различается по величине и распределению электрического заряда (что позволяет выполнить их электрофоретическое разделение), термостабильности, оптимальной величине рН, отношению к ингибиторам, действию на «вторичные» субстраты [16]. Очень важно, что каждый изофермент (точнее – каждая изоферментная форма данного фермента) или их заданный набор локализован в определенных органах и тканях животных, причем все изоферменты имеют единое каталитическое действие на основной субстрат. Установлены изоферментные формы большого числа дегидрогеназ, трансаминаз, оксидаз, фосфатаз, фосфорилаз [14–16]. Ниже будут описаны только сравнительно хорошо изученные изоферментые формы лактатдегидрогеназы и креатинкиназы, а также ряд других ферментов, важных для биохимических исследований и клинической диагностики.
Лактатдегидрогеназа
Лактатдегидрогеназа (ЛДГ) – КФ. 1.1.1.27. - различных органов и тканей человека или животных (рис. 2-4) всегда состоит из 4 полипептидных субъединиц одного или двух типов: М («мышечный» тип субъдиниц) или Н («сердечный» тип субъединиц) [16]. Поэтому лактатдегидрогеназу с полным основанием можно называть супрамолекулярной ферментной системой, которая имеет 4 активных центра, включающих обязательно (в случае ЛДГ человека и животных) по молекуле кофермента – никотинамидадениндинуклеотида (НАД+ или НАДН). ЛДГ, выделенная в кристаллическом виде из мышечной ткани позвоночных, имеет молекулярную массу тетрамера порядка 140000.
Показано, что полипептидная цепь Н-субъединицы ЛДГ1, выделенной из сердечной мышцы человека, состоит из 332 аминокислот с каталитическим центром Asp166 - His193 - Arg169 [16], тогда как полипептидная цепь М-субъединицы ЛДГ5, выделенной из скелетных мышц человека, состоит из 331 аминокислоты, но с каталитическим центром Asp165 - His192 - Arg168.
 |  |
| Рисунок 2 – Схематическое изображение М-субъединиц ЛДГ, выделенной из скелетных мышц свиньи (справа) и Н-субъединицы ЛДГ, выделенной из сердечной мышцы свиньи (слева), в комплексе с 2 молекулами НАДН: коды 9ldb (PDB-EBI) и 5ldh (PDB-EBI), соответственно. | |
Полипептидная цепь М-субъединицы ЛДГ, выделенной из скелетных мышц свиней (sus scrofa domestica), состоит из 331 аминокислоты (рис. 2 и 3), тогда как полипептидная цепь Н-субъединицы ЛДГ, выделенной из сердечной мышцы свиньи, состоит из 334 аминокислот. Эти различия не влияют на природу и положение аминокислот каталитического центра ЛДГ свиней – в обоих случаях эта последовательность Asp168 - His195 - Arg171 (рис. 4 и 5). В доступной литературе не удалось найти структуры для изоферментов ЛДГ собак, но только отдельных видов мышей и рыб, у которых Н- и М-субъединицы ЛДГ практически не отличаются (330 или 331 аминокислота с их одинаковой последовательностью в каталитических центрах : Asp168 - His195 - Arg171.
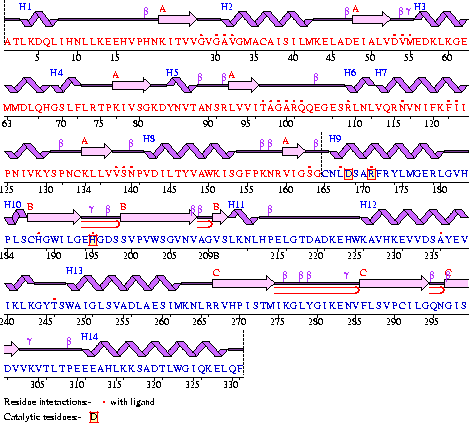 |
| Рисунок 3 – Аминокислотная последовательность М-субъединицы ЛДГ5, выделенной из мышц свиньи (sus scrofa domestica): код 9ldb (PDB-EBI). |
 |
| Рисунок 4 – Строение активного центра М-субъединицы ЛДГ, выделенной из мышц свиньи (sus scrofa domestica) в комплексе с коферментом НАДН: код 9ldb (PDB-EBI). |
 |  |
| Рисунок 5 – Структура кофермента НАДН (слева) и его локализация (справа) в М-субъединицах ЛДГ5, выделенная из мышц свиньи код 9ldb (PDB-EBI). | |
Лактатдегидрогеназа – фермент класса оксидоредуктаз (L-лактат:NAD-оксидоредуктаза) обратимо катализирующий окисление молочной кислоты (L–лактата) в пировиноградную кислоту (пируват), где в качестве промежуточного акцептора водорода требуется кофермент – никотинамидадениндинуклеотид: окисленная форма (НАД+) и восстановленная форма (НАДН+Н+).

В большинстве животных тканей равновесие катализируемой ЛДГ реакции сильно сдвинуто в сторону образования молочной кислоты; требует присутствия НАДН+Н+ и протекает в анаэробных условиях. Исключение составляют раковые клетки: в них образуется большое количество молочной кислоты в аэробных условиях. Фермент встречается во всех органах и тканях, но наибольшая активность ЛДГ обнаружена в почках, сердце, скелетной мускулатуре и печени. ЛДГ содержится также и в эритроцитах, поэтому сыворотка, используемая для анализа, должна быть лишена даже следов гемолиза. Повышение общей активности в сыворотке отмечается при инфаркте миокарда, лейкозах, тромбоцитопении, повреждениях печени вирусной, токсической и травматической природы, опухолях различной локализации, заболеваниях почек, гемолизе эритроцитов [16].
В тканях животных выявлено 5 изоферментных форм ЛДГ, которые являются тетрамерами, причем каждый орган имеет свой «изоферментный спектр» ЛДГ. Так, тетрамер Н4 или ЛДГ1 – преимущественно локализован в сердечной мышце; тетрамер М4 или ЛДГ5 - локализован как в печени, так и в поперечно-полосатой мышечной ткани; другие три изофермента – гибриды: Н3M1 или ЛДГ2 ; Н2M2 или ЛДГ3 ; Н1M3 или ЛДГ4 – преимущественно локализованы в ретикулоэндотелиальной системе, легких и почках, соответственно. Интересно, что изофермент Н4 помимо реакции «пировиноградная кислота-молочная кислота» может катализировать реакцию «кетоглутаровая кислота-гидроксимасляная кислота», но легко ингибируется пируватом и поэтому особенно полезен для такого «высокоаэробного органа» как сердце. Хотя изофермент М4 эффективен лишь в реакции «пировиноградная кислота-молочная кислота», но он не ингибируется пируватом и поэтому особенно полезен для скелетных мышц, где может создаваться большая нагрузка, требующая мощных процессов анаэробного гликолиза. Обычно активность изофермента Н3М1 в крови человека и животных незначительно выше, чем других изоферментов. Поэтому увеличение активности М4 свидетельствует о патологии печени, а в случае инфаркта миокарда активность изоферментов Н4 и M1Н3 становится значительно выше активности других изоферментов. Указанные различия в активности изоферментов в крови имеют важное значение для клинической диагностики патологических состояний определенных органов и тканей как человека, так и животных.
Креатинкиназа
Креатинкиназа (КК) – КФ. 2.7.3.2. - гетерогенный фермент с молекулярной массой порядка 86000-89000 (рис. 6-11). Комплекс КК состоит из двух субъединиц 2 разных типов (В и М), обладающих молекулярной массой 44500 и 43000 соответственно [16]. В приложение 2 приведено строение полипептидных цепей креатинкиназы (типа ММ) из мышц человека, состоящих из 365 аминокислотных остатков. Каталитический центр состоит из 5 аминокислотных остатков Arg132 - Arg320 - Glu232 - Arg292 - Arg236.
Креатинкиназа катализирует как прямую реакцию переноса фосфорильного остатка с АТФ на креатин, так и обратную реакцию переноса фосфорильного остатка с креатинфосфата на АДФ:
АТФ + Креатин АДФ + Креатинфосфат.
Скорость прямой реакции максимальна при рН 9,0; обратной – при рН 6-8. В организме равновесие сдвинуто в сторону обратной реакции. Активность фермента повышается под влиянием ионов магния, марганца, кальция, глутатиона и цистеина, а ионы цинка, меди и ртути, напротив, оказывают ингибирующее воздействие [16].
Креатинкиназа (типа КК-ВВ) из мозга цыпленка (Gallus gallus) состоит из двух В-субъединиц (рис. 6): код 1qh4 (PDB-EBI). Полипептидная цепь В-субъединицы креатинкиназы (типа КК-ВВ) из мозга цыпленка состоти из 380 аминокислотных остатков. Хотя эта последовательность отличается, но порядок аминокислотных остатков в калитическом центре этой В-субъединицы Arg132 - Arg320 - Glu232 - Arg292 - Arg236 полностью совпадает с таковой для М-субъединицы креатинкиназы (типа ММ) из мышц человека код 1i0e (PDB-EBI).
Безусловно «супрамолекулярным» является строение ассиметрического комплекса и биологически активного октамера (рис. 7) митохондриальной креатинкиназы (КК-мит.) из сердечной мышцы цыпленка (Gallus gallus): код 1crk (PDB-EBI). Полипептидная цепь субъединицы КК-мит. из сердечной мышцы цыпленка состоит из 380 аминокислотных остатков, как и для приведенной выше В-субъединицы типа КК-ВВ из мозга цыпленка. Однако порядок аминокислотных остатков в калитическом центре этой митохондриальной субъединицы Arg231(A) - Arg287(A) - Arg127(A) - Arg315(A) - Glu227(A) существенно отличается от приведенной выше В-субъединицы креатинкиназы (типа КК-ВВ) из мозга цыпленка.
Необычным является строение ассиметрического комплекса и биологически активного димера креатинкиназы из сердечной мышцы кролика (Oryctolagus cuniculus) (рис. 8). Полипептидная цепь субъединицы креатинкиназы из сердечной мышцы кролика состоит из 365 аминокислотных остатков. Как эта последовательность, так и порядок аминокислотных остатков в калитическом центре этой субъединицы Arg132 - Arg320 - Glu232 - Arg292 - Arg236 полностью совпадает с таковой для М-субъединицы креатинкиназы (типа ММ) из мышц человека.
 |  |
| Рисунок 6 – Схематическое изображение ассиметрического комплекса (слева) и биологически активного димера (справа) креатинкиназы (типа КК-ВВ) из мозга цыпленка (Gallus gallus): код 1qh4 (PDB-EBI). | |
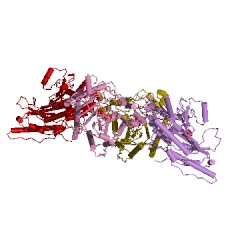 |  |
| Рисунок 7 – Схематическое изображение ассиметрического комплекса (слева) и биологически активного октамера (справа) митохондриальной креатин-киназы (типа КК-мит) из сердечной мышцы цыпленка: код 1crk (PDB-EBI). | |
 |  |
| Рисунок 8 – Схематическое изображение ассиметрического комплекса (слева) и биологически активного димера (справа) креатинкиназы из сердечной мышцы кролика (Oryctolagus cuniculus. Rabbit): код 2crk (PDB-EBI). | |
Одним из объяснений этого феномена может служить наличие внутри каждой группы изоферментов нескольких изоформ, отличающихся по своим физико-химическим свойствам. Эти факты усложняют и делают поистине «супрамолекулярной» ставшую уже классической теорию о том, что «комбинация двух субъединиц в димерную структуру приводит к образованию трех изоферментов, называемых по входящим в их состав субъединицам КК-ММ, КК-ВВ и КК-МВ изоэнзимами, и локализованных в скелетных мышцах, клетках мозга и сердечной мышце, соответственно» [16]. Кроме того, в тканях миокарда содержится и ММ-изофермент КК. В сыворотке крови здорового человека имеет место следующее соотношение изоферментов КК: активность КК -ММ составляет 94-96 %, КК-МВ - 4-6 %, изофермент КК-ВВ присутствует в следовых количествах.
Согласно современным представлениям, в энергетическом обеспечении функции миофибрилл и саркоплазматического ретикулума сердечной мышцы ведущее место занимает креатинкиназный механизм, который в значительной мере определяет скорость и характер внутриклеточного перераспределения энергии и утилизацию ее субклеточными структурами. Поэтому активность креатинкиназы в сыворотке крови повышается при остром инфаркте миокарда. При этом активность креатинкиназы увеличивается раньше, чем изменяется активность других ферментов. Однако значительное повышение активности креатинкиназы в сыворотке крови может наблюдаться также при повреждении скелетной мускулатуры и при самых различных нарушениях центральной нервной системы. Считается, что повышение общей активности КК при ряде заболеваний центральной нервной системы связано с увеличением проницаемости клеточных мембран скелетных мышц и сердца, а также понижением скорости инактивации и выделения фермента из кровотока. На основании всех известных данных можно сделать вывод, что наибольшее распространение в клинической лабораторной практике приобрело определение общей КК и ее МВ-составляющей, причем определение активности МВ-изофермента КК имеет особо важное значение при диагностике инфаркта миокарда и мониторинге постинфарктного состояния.
Аспартатаминотрансфераза и аланинаминотрансфераза
Считается оптимальным рассматривать совместно ферменты КФ 2.6.1.1. или аспартатаминотрансферазу (АСТ) и КФ 2.6.1.2. или аланинамино-трансферазу (АЛТ), которые имеют много общего в строении и свойствах, а также учитывая недостаток детальных структурных данных по АЛТ в отличие от АСТ. Как АЛТ, так и АСТ состоят из двух полипептидных субъединиц одинакового типа с ММ порядка 45000, т.е. ММ каждого фермента составляет порядка 90000 [16]. Субъединицы имеют «богатую» вторичную структуру с двумя выраженными доменами, один из которых необходим для связывания кофермента (пиридоксальфосфата). По последним данным, полученным методом рентгеновской дифракции кристаллов митохондриального фермента с разрешением 0,28 нм, считается, что оба активных центра димера находятся на границе между субъединицами и состоят из аминокислотных остатков обеих субъединиц. Для АСТ показано, что остаток Arg292 в обычном ферменте в основном «отвечает» за специфичность к субстрату. Мутантный по данному остатку фермент (замена на Asp-292 приводит к инверсии зарядового баланса) становится активным в реакции трансаминирования аргинина, а не аспартата [16].
АСТ из митохондрий сердечной мышцы цыпленка (gallus gallus): код 1map (PDB-EBI) состоит из двух одинаковых субъединиц (рис. 9). Поли-пептидная цепь субъединицы АСТ из митохондрий сердечной мышцы цыпленка состоит из 401 аминокислотного остатка. Считается, что кофермент ПФ всегда связывается с Lys258 (рис. 10) и порядок амино-кислотных остатков в каталитическом центре этой субъединицы Trp140 - Asp222 - Lys258 достаточно консервативен для всех АСТ. Всего в активном центре субъединицы АСТ насчитывается 19 аминокислотных остатков.
Надо обязательно учитывать, что в этом активном центре находятся также еще 4 аминокислотных остатка из другой субъединицы АСТ. Интересно, что АСТ из цитоплазмы клеток сердечной мышцы цыпленка (gallus gallus): код 2cst (рис. 11) состоит из двух одинаковых субъединиц, но состоящих из 411 аминокислотных остатков каждая. Всего в активном центре субъединицы (обозначенной А) этой АСТ насчитывается 19 аминокислотных остатков (с рядом отличий) и еще 4 аминокислотных остатка из другой субъединицы (В) этой же АСТ. Важно, что порядок и положение аминокислотных остатков в каталитическом центре субъединицы этой АСТ неизменен Trp140(A) - Asp222(A) - Lys258(A), несмотря на увеличение (на 10 единиц) общего числа аминокислотных остатков для АСТ из цитоплазмы клеток сердечной мышцы цыпленка, по сравнению с АСТ из митохондрий этой же мышцы.
АСТ катализирует обратимый перенос аминогруппы с аспарагиновой кислоты на -кетоглутаровую кислоту с образованием оксалоацетата:
 |
Наибольшее содержание АСТ обнаружено в сердечной мышце, в печени, скелетной мускулатуре, головном мозге, почках, семенниках. Так, активность АСТ в сердечной мышце почти в 10000 раз выше, чем в сыворотке крови. Поскольку даже в эритроцитах активность АСТ в 10 раз выше, чем в сыворотке, то исследуемая сыворотка крови не должна иметь следов гемолиза. Увеличение активности АСТ в сыворотке крови происходит при инфаркте миокарда (в 4-5 раз) и сохраняется 3-5 дней. Умеренное увеличение активности АСТ обнаруживается также у больных с метастазами в печень, циррозом печени, при прогрессивной мышечной дистрофии, а также при некрозе или повреждении печеночных клеток любой этиологии, включая холестатическую и обтурационную желтуху, острый и хронический гепатит, причем в большинстве этих случаев уровень активности АЛТ выше уровня активности АСТ [16].
 |  | |
| Рисунок 9 – Изображение одной субъединицы (слева) и димера (справа) АСТ из митохондрий сердечной мышцы цыпленка (gallus gallus): 1map (PDB-EBI). | ||
  |  | |
| Рисунок 10 – Структура кофермента ПФ и субстрата (слева) и их локализация (справа) в субъединице аспартатаминотрансферазы из митохондрий сердечной мышцы цыпленка (gallus gallus): код 1map (PDB-EBI) | ||
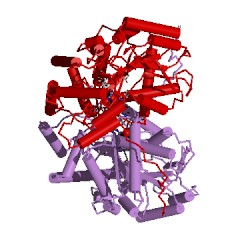 | 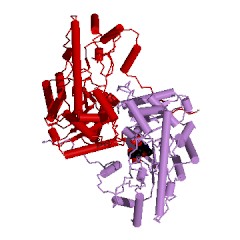 |
| Рисунок 11 – Изображение димеров АСТ, полученных из цитоплазмы клеток сердечной мышцы цыпленка (gallus gallus): код 2cst (слева) и свиньи (Sus scrofa): код 1ajs (справа) | |
 |
Аланинаминотрансфераза катализирует обратимый перенос аминогрупп с L-аланина на -кетоглутаровую кислоту с образованием пирувата и L-глутаминовой кислоты:
Наиболее высокая активность АЛТ обнаруживается в печени, поджелудочной железе, сердце, и скелетной мускулатуре. Повышение активности АЛТ является наиболее специфичным признаком заболеваний печени, особенно острых, что имеет большое диагностическое значение. Как правило, оно возникает за 1-4 недели до развития клинических симптомов и за 7-10 дней до обнаружения максимального уровня билирубина в крови, более чем в 5-10 раз превышая норму. Длительное увеличение активности АЛТ или повышение активности в поздние сроки заболевания может означать начало массивного печеночного некроза. Повышение активности АЛТ в сыворотке крови отмечается при гепатитах различной этиологии, механической желтухе, циррозе печени, введении гепатотоксических препаратов, инфаркте миокарда. В норме соотношение активностей АСТ/АЛТ (коэффициент де Ритиса) для человека равно 1,33±0,42. У больных инфекционным гепатитом происходит снижение коэффициента, а при остром инфаркте миокарда резко возрастает [16].
Гамма-глутамилтрансфераза
Фермент КФ. 2.3.2.2. или -глутамилтрансфераза (ГГТ) – мембрано-связанный белок сложного строения: димер ГГТ состоит из субъединиц А и В, содержащих разное число аминокислот (рис. 12). Таких димеров может быть несколько в «мембранно-трасферазном комплексе», который с полным основание можно считать «ферментной» СБС (рис. 12). К сожалению, детали структур ГГТ животных в доступной литературе отсутствуют, поэтому здесь приведены данные для бактерий. Димер ГГТ, выделенный из бактерий (Escherichia coli. Strain: k-12.), состоит из субъединиц, содержащих 359 (А) и 109 (В) аминокислотных остатков. Несмотря на то, что В-субъединица этой ГГТ почти в 3,5 раза короче А-субъединицы, но в ней взаимодействует с лигандом или субстратом в 3 раза больше амино-кислотных остатков (15), чем в А-субъединице (6), что следует из строения активного центра ГГТ (рис. 13).
ГГТ является ключевым из 6 ферментов -глутамильного цикла, служащего для переноса ряда аминокислот через плазматические мембраны. Данный фермент присутствует в высоких концентрациях в печени, почках и поджелудочной железе. Его активность в плазме является чувствительным индикатором заболеваний гепатобилиарной системы, хотя он и не дает возможности разграничить холестатический и гепатоцеллюлярный типы патологии. При обструкции желчных путей увеличение активности ГГТ в плазме может предшествовать подъему активности щелочной фосфатазы. Активность ГГТ в плазме возрастает при отсутствии заболеваний печени у пациентов, принимающих антиконвульсанты (аналогичное действие может оказывать рифампицин при лечении туберкулеза). В указанных выше случаях повышение активности ГГТ в плазме происходит не вследствие повреждения клеток, а в результате усиления внутриклеточного синтеза фермента и освобождения его повышенного количества в циркуляцию в процессе нормального метаболизма клеток. Повышение активности ГГТ плазмы связано также с холестазом (превышение верхнего предела нормы более чем в 10 раз); гепатитом (острый и хронический); циррозом (без холестаза); панкреатитом (превышение верхнего предела нормы в 5-10 раз); многими состояниями, отражающими «вторичные воздействия» на печень: с лекарствами, вызывающими индукцию фермента; с сердечной недостаточ-ностью (превышение верхнего предела нормы менее чем в 5 раз) [16].
 |  |
| Рисунок 12 – Изображение асимметрического комплекса – тетрамера (слева) и димера (справа) ГГТ, выделенной из бактерий (Escherichia coli. Strain: k-12.): код 2dbw (PDB-EBI) | |
 |
| Рисунок 13 – Строение активного центра для субъединицы ГГТ из бактерий (Escherichia coli. Strain: k-12.): код 2dbw (PDB-EBI) |
Все эти ферменты и их изоформы в крови животных составляют характерный набор – «ферментный профиль», который изменяется при патологических изменениях тканей, что активно используют в клинической биохимической диагностике.
Липолитические ферменты (липазы из поджелудочной железы животных, бактерий и грибов)
Различные типы липаз традиционно относятся к липолитическим ферментам (это прежде всего гидролазы эфиров карбоновых кислот) и играют важную роль в обмене липидов у животных [20–25]. Наиболее важной для данной работы являлась панкреатическая липаза или триацилглицероллипаза (КФ 3.1.1.3) как гидролаза сложных эфиров глицерина и жирных кислот или, по-другому, ацилгидролаза различных триацилглицеридов. Панкреатическая липаза преимущественно вырабатывается поджелудочной железой животных и принимает участие в расщеплении нейтральных жиров, поступающих в кишечник с пищей. Панкреатическая липаза – один из активно исследуемых липолитических ферментов, поэтому ее часто используют в качестве модели для других липаз (как животного, так и растительного или микробного происхождения).
Использование идеологии супрамолекулярных систем особенно полезно при структурно-функциональном исследовании комплексов молекул субстрата и димеров или октамеров панкреатической липазы в комплексе с колипазой (рис. 14), выделенных из поджелудочной железы свиньи (Sus scrofa): код 1eth (PDB-EBI). На рис. 15 и 16 приведены полипептидные последовательности для субъединиц липазы и колипазы свиньи (Sus scrofa), состоящих из 448 и 87 аминокислотных остатков, соответственно. Для правильного структурного представления этой достаточно сложной супрамолекулярной системы необходимо добавить 1071 молекулу воды, хотя бы по одному катиону Са2+ и одной молекуле субстрата (рис. 17) на каждый комплекс липазы с колипазой в димере или октамере (рис. 14 и 18). Строение каталитического центра липазы: Ser153, Asp80 и Gly77, отличается от «субстрат-связывающего» центра. Предполагается, что комплекс липазы с колипазой образуется 16 и 12 аминокислотными остатками соответствующих белков с использованием 8 водородных связей и от 96 до 100 «других контактов», хотя «критическими» для такой супрамолекулярной организации организация этих комплексов являются только 4 аминокислотных остатка с каждой стороны и 15 «других типов» контактов.
Гидролиз триглицеридов в желудочно-кишечном тракте животных осуществляется панкреатическими липазами, которые продуцируются у млекопитающих в ацинарных клетках поджелудочной железы и накапливаются в зимогенных гранулах, содержащих набор пищеварительных ферментов или их проферментов. Из этих гранул липаза высвобождается в специальный проток, по которому панкреатический сок поступает к месту своего назначения — в тонкий кишечник. Данных, свидетельствующих о наличии профермента (зимогена) липазы, не имеется и нет никаких причин думать, что таковой существует. В отличие от протеиназ или фосфолипаз, панкреатическая липаза обладает особой специфичностью, которая заключается в том, что липаза действует на свой субстрат — триглицерид только в том случае, если он находится в эмульгированном состоянии. Такое эмульгирование липидов эффективно осуществляется желчью при участии солей желчных кислот (в печени синтезируются холевая, дезоксихолевая, гликохолевая и таурохолевая кислоты), что увеличивает скорость расщепления триглицеридов панкреатической липазой. Адсорбируясь и уменьшая поверхностное натяжение жировых капелек, желчные кислоты раздробляют их и превращают жиры и масла в тонкую эмульсию, диаметр частиц которой не превышает 0,5 мкм. Эмульгирование жира приводит к увеличению поверхности соприкосновения триглицерида с водным раствором липазы, т. е. облегчает ферментативный гидролиз [16, 24].
 | 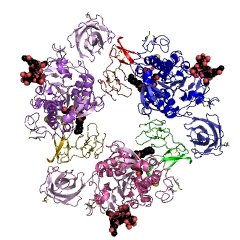 |
| Рисунок 14 – Схематическое изображение супрамолекулярных систем молекул субстрата и димера (слева) и октамера (справа) комплексов липазы из поджелудочной железы свиньи (Sus scrofa) и колипазы : код 1eth (PDB-EBI) | |
 |
| Рисунок 15 – Полипептидная последовательность для липазы из поджелудочной железы свиньи (Sus scrofa): код 1eth (PDB-EBI). |
 |
| Рисунок 16 – Полипептидная последовательность для колипазы из поджелудочной железы свиньи (Sus scrofa): код 1eth (PDB-EBI). |
 | |||
| Рисунок 17 – Строение активного центра с включенным аналогом субстрата для липазы из поджелудочной железы свиньи (Sus scrofa): код 1eth (PDB-EBI). | |||
  |
| ||
Рисунок 18 – Структура гидрофобного аналога субстрата (слева) и локализация двух таких остатков (справа) в димере липазы из поджелудочной железы свиньи (Sus scrofa): код 1eth (PDB-EBI). | |||

Панкреатическая липаза свиньи является гликопротеидом, имеющим молекулярную массу порядка 50 000 и оптимум рН 8-9 [24]. Данный фермент расщепляет триглицериды, находящиеся в эмульгированном состоянии, но может гидролизовать также ди- и моноглицериды [24].
 |