

Влияние моделирования эффектов микрогравитации на цитоскелет и экспрессию генов у мезенхимальных стромальных клеток-предшественников костного мозга человека in vitro
На правах рукописи
ГЕРШОВИЧ
Павел Михайлович
Влияние моделирования эффектов микрогравитации на цитоскелет и экспрессию генов у мезенхимальных стромальных клеток-предшественников костного мозга человека in vitro
14.03.08 – авиационная, космическая и морская медицина
03.03.04 - клеточная биология, цитология, гистология
А В Т О Р Е Ф Е Р А Т
диссертации на соискание ученой степени
кандидата биологических наук
Москва – 2010
Работа выполнена в Учреждении Российской академии наук Государственном научном центре Российской Федерации – Институте медико-биологических проблем РАН
Научный руководитель:
доктор медицинских наук, профессор Буравкова Людмила Борисовна
Официальные оппоненты:
доктор медицинских наук, профессор Оганов Виктор Сумбатович доктор биологических наук, профессор Киселев Сергей Львович
Ведущая организация: Учреждение Российской академии наук
Институт молекулярной генетики РАН
Защита диссертации состоится «16» сентября 2010 г. в 1000 часов на заседании диссертационного совета Д 002.111.01 в Учреждении Российской академии наук Государственном научном центре Российской Федерации – Институте медико-биологических проблем РАН (ГНЦ РФ – ИМБП РАН) по адресу: 123007, г. Москва, Хорошевское шоссе, д. 76 а.
С диссертацией можно ознакомиться в библиотеке ГНЦ РФ – ИМБП РАН
Автореферат разослан « »______________2010 г.
Ученый секретарь диссертационного совета,
доктор биологических наук М.А. Левинских
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы. Гравитация является фундаментальным механическим фактором, определяющим особенности эволюции и структурно-функциональной организации живых организмов на Земле. Известно, что в условиях микрогравитации происходят атрофические перестройки костной ткани, выраженность которых во многом зависит от расположения кости по отношению к вектору гравитации [Ступаков Г.П., Воложин А.И., 1989; Газенко О.Г., Григорьев А.И., Егоров А.Д., 1997; Оганов В.С., Богомолов B.B., 2009; Vico L. et al., 2001]. Локальная потеря массы костной ткани в условиях дефицита механической нагрузки или микрогравитации позволяет предположить, что рецепция механического сигнала (или его отсутствия) может осуществляться, в том числе и на клеточном уровне. Обусловленное микрогравитацией изменение способности костной ткани к самовозобновлению указывает на то, что в этот процесс могут быть вовлечены стволовые клетки и остеогенные клетки-предшественники. Не исключено, что пусковым механизмом могут служить различные изменения в структурах цитоскелета клеток остеогенной природы. В ряде недавних работ было показано участие определенных структур цитоскелета стволовых клеток в выборе пути их дифференцировки [Sordella R. et al., 2003; McBeath R. et al., 2005]. Цитоскелет живой клетки наряду с мембранными компонентами является одной из ведущих базовых структур клетки, обеспечивающих ее морфологическую целостность и функциональную активность. Ремоделирование актинового цитоскелета происходит под влиянием факторов различной природы: факторов роста, межклеточной адгезии и адгезии клеток к субстрату, осуществляемой, главным образом, интегриновыми рецепторами, а также при действии механических сигналов [Юдинцева Н.М. и др., 2008; Buravkova L.B. et al., 2001; Bershadsky A.D. et al., 2003; Crawford-Young S.J., 2006]. Существуют исследования, свидетельствующие об участии актинового цитоскелета и ассоциированных с ним внеклеточных и внутриклеточных белков в процессах проведения сигнала и регуляции экспрессии генов [Пинаев Г.П., 2009; Aplin A.E., et al., 1998; Janmey P.A., 1998]. Кроме того, показано, что в ходе остеогенной дифференцировки мезенхимальных клеток-предшественников организация актинового цитоскелета изменяется [Rodrguez J.P. et al., 2004]. Однако, по-прежнему, остается неясной роль цитоскелета, как потенциально гравитационно-чувствительной структуры клеток, в механизмах их адаптации к микрогравитации, а также в том, какое значение могут иметь перестройки цитоскелета или изменения в экспрессии генов компонентов цитоскелета для нормальной функциональной активности клеток-предшественников. Мультипотентные мезенхимальные стромальные или стволовые клетки (ММСК) являются популяцией клеток-предшественников стромы костного мозга [Pittenger M.F. et al., 1999; Conget P.A., Minguell J.J., 1999; Caplan A.I., Bruder S.P., 2001; Bianco P. et al., 2001]. Дифференцировочный потенциал, а также другие особенности функционирования этих клеток могут быть сравнительно легко изучены в культуре, в том числе и при оценке влияния различных физических факторов [Altman G.H. et al., 2002; Sordella R.et al., 2003; McBeath R. et al., 2005]. Перспективность всестороннего изучения биологии ММСК, а также их клинического использования определяется, прежде всего, их способностью восстанавливать дефекты и повреждения соединительных тканей (костной, хрящевой и мышечной). Точные молекулярные механизмы гравитационной чувствительности ММСК и более коммитированных остеогенных клеток-предшественников остаются не до конца ясными, что обусловлено сложностью объекта исследования, многоуровневой системой регуляции их дифференцировочного потенциала и функциональной активности. В частности, показано, что моделирование эффектов микрогравитации приводит к ингибированию остеогенной дифференцировки ММСК и преостеобластов и к активации программы адипогенной дифференцировки ММСК [Zayzafoon M. et al., 2004; Meyers V.E. et al., 2004; 2005; Pardo S.J. et al., 2005; Dai Z.Q. et al., 2007; Patel M.J. et al., 2007; Pan Z. et al., 2008; Gershovich J.G. et al., 2008; Capulli M. et al., 2009; Huang Y. et al., 2009], что выражается в подавлении уровня экспрессии определенных генов-маркеров остеогенного коммитирования, и связывается с перестройками актинового цитоскелета клеток и изменением уровня экспрессии ассоциированных с цитоскелетом механочувствительных адгезивных рецепторов клетки (интегринов). В целом, исследования, посвященные изучению гравитационной чувствительности ММСК in vitro, по-прежнему, весьма немногочисленны. Таким образом, изучение состояния цитоскелета и молекулярно-генетических свойств ММСК в условиях моделирования эффектов микрогравитации является актуальным вопросом, решение которого позволит приблизиться к более глубокому пониманию процессов, происходящих с клетками-предшественниками в условиях реальной микрогравитации.
Цель работы: изучение особенностей организации цитоскелета, ассоциированных с ним элементов и молекулярно-генетическая характеристика мультипотентных мезенхимальных стромальных клеток (ММСК) костного мозга человека в условиях моделирования эффектов микрогравитации.
Задачи исследования:
1) изучить динамику реорганизации актинового и тубулинового цитоскелета культивируемых ММСК на различных этапах наземного моделирования эффектов микрогравитации с помощью RPM;
2) исследовать адгезивные свойства ММСК и экспрессию некоторых рецепторов адгезии при моделировании эффектов микрогравитации;
3) оценить уровень экспрессии генов основных белков цитоскелета и ассоциированных с ним элементов на различных этапах наземного моделирования эффектов микрогравитации с помощью RPM;
4) провести профилирование экспрессии 84 генов – «маркеров стволовых клеток человека» с помощью наборов Stem Cell RT Profiler™ PCR Array на различных этапах наземного моделирования эффектов микрогравитации с помощью RPM;
5) провести анализ уровня экспрессии ключевых маркеров остеогенной дифференцировки ММСК при длительном моделировании эффектов микрогравитации.
Научная новизна.
• Впервые показано, что моделирование эффектов микрогравитации с помощью RPM приводит к реорганизации актинового цитоскелета ММСК костного мозга человека уже после нескольких минут воздействия, которая является обратимой при более продолжительной экспозиции (120 часов) и при статической ре-адаптации в течение 24 часов, и потере ими периферических винкулиновых сайтов фокальной адгезии, не влияя на организацию тубулинового цитоскелета клеток.
• Впервые выявлена динамика изменения уровня экспрессии генов цитоскелета в ММСК. В частности, показано, что экспозиция на RPM вызывает транзиторное изменение уровня экспрессии структурных и регуляторных генов, связанных с актиновым цитоскелетом, наиболее выраженное на этапе 48 часов, которое частично компенсируется на конечных сроках экспозиции (120 часов) и полностью при ре-адаптации клеток в статических условиях в течение 24 часов, что может отражать процесс адаптации цитоскелета к моделированию эффектов микрогравитации.
• Впервые установлено, что моделирование эффектов микрогравитации (120 ч) приводит к достоверному изменению паттерна экспрессии генов - «маркеров стволовых клеток» (Stem Cell RT Profiler™ PCR Array, SABiosciences) в ММСК костного мозга человека. При этом, моделирование эффектов микрогравитации вызывает как активацию, так и угнетение экспрессии 54 различных генов, кодирующих белки, принимающие участие во внутриклеточной сигнализации, клеточной адгезии, регуляции пролиферации и дифференцировки стволовых клеток.
• Показано, что коммитирование ММСК к остеогенезу в условиях длительного моделирования эффектов микрогравитации (10-20 сут) сопровождается снижением уровня экспрессии генов, кодирующих основные маркеры остеогенной дифференцировки клеток, в том числе и ключевого мастер-транскрипционного фактора, контролирующего программу остеогенной дифференцировки ММСК – RUNX2.
Теоретическая и практическая значимость работы.
Обнаруженные структурные и молекулярно-генетические изменения в культурах ММСК свидетельствуют о наличии гравитационно-зависимых внутриклеточных механизмов, которые обуславливают как ранние, так и поздние ответные реакции клеток-предшественников на моделирование эффектов микрогравитации. Полученные приоритетные данные существенно дополняют имеющиеся теоретические представления о механизмах восприимчивости клеток-предшественников взрослого организма к изменению параметров гравитационной среды, по крайней мере, в условиях in vitro и о роли актинового цитоскелета, как базовой структуры клеток, в том числе и стволовых, в качестве гравитационно-сенсорной клеточной структуры. Результаты работы подтверждают возможность использования мезенхимальных стромальных клеток костного мозга человека в качестве удобной экспериментальной модели для изучения роли гравитации и микрогравитации в процессах функционирования и дифференцировки клеток-предшественников взрослого организма, а также для исследования механизмов клеточной гравитационной чувствительности. Полученные в работе результаты являются основой для разработки проекта технического задания для проведения космического эксперимента с целью дальнейшего изучения морфофункциональных и молекулярно-генетических свойств ММСК в условиях микрогравитации.
Положения, выносимые на защиту:
1. Актиновый цитоскелет и ассоциированные с ним элементы представляют собой гравитационно-чувствительные структуры клетки, которые наиболее быстро реагируют на постоянное изменение положения ММСК в пространстве, что подтверждается наблюдаемой транзиторной реорганизацией актинового цитоскелета, и изменением в уровне экспрессии связанных ним структурных и регуляторных генов, начиная с самых ранних сроков экспозиции на RPM (30 мин - 48 ч). Восстановление структуры актинового цитоскелета ММСК на более поздних этапах (120 ч) сопровождается активацией адгезии клеток и частичным восстановлением уровня экспрессии цитоскелетных генов, что может быть важным звеном в механизме адаптации цитоскелета клеток-предшественников к условиям моделирования эффектов микрогравитации.
2. Под влиянием моделирования эффектов микрогравитации в ММСК человека происходит как угнетение, так и активация экспрессии ряда генов, отвечающих за поддержание свойств мультипотентности и дифференцировки стволовых клеток различных типов. В отличие от генов, связанных с актиновым цитоскелетом, паттерн экспрессии генов, кодирующих маркеры «стволовых клеток» в ММСК человека, наиболее значительно изменяется после 120 часов экспозиции в условиях моделирования эффектов микрогравитации с помощью RPM, что отражает временные различия между механизмами изменения экспрессии структурных и регуляторных генов, ответственных за различные типы внутриклеточных процессов в стволовых клетках.
3. Подавление процесса индуцированной остеогенной дифференцировки ММСК в условиях моделирования эффектов микрогравитации с помощью RPM может быть обусловлено как угнетением экспрессии генов, кодирующих ключевые маркеры остеогенной дифференцировки клеток, а именно, RUNX2, ALPL и OMD, так и активацией экспрессии анти-остеогенных и про-адипогенных факторов, таких как PPAR.
Апробация работы. Основные результаты и положения диссертации были доложены и обсуждены на: 7-й – 8-й Конференциях молодых учёных и специалистов, аспирантов и студентов, посвящённых дню Космонавтики. Москва, 2008, 2009; 11-й, 12-й, 14-й международной Пущинской школе-конференции молодых ученых «Биология-наука XXI века». Пущино, 2007, 2008, 2010; International symposium «Biological motility»: Achievements and Perspectives. Pushchino. 2008; 29th Annual International Physiology Meeting. Angers. France. 2008; 17th Humans in Space Symposium. Moscow. 2009; French-Russian-Belarussian Conference «Neurovascular impairment induced by environmental conditions: molecular, cellular and functional approach». Angers. France. 2010.
По теме диссертации опубликовано 14 печатных работ, в том числе 4 статьи в журналах, рекомендованных ВАК РФ.
Диссертация апробирована на заседании секции Ученого совета ГНЦ РФ – ИМБП РАН “Космическая физиология и биология” 15.06.2010 г.
Связь работы с научными программами. Работа выполнена при поддержке гранта РФФИ №08-04-01607-а, контракта с Роснауки № 02.518.11.7059 и программы Фундаментальных исследований ГНЦ РФ - ИМБП РАН.
Структура и объем диссертации. Диссертация состоит из введения, обзора литературы, описания материалов и методов исследования, результатов исследования и их обсуждения, заключения, выводов и списка литературы. Текст диссертации изложен на 150 страницах, содержит 18 рисунков и 8 таблиц. Список литературы состоит из 307 цитируемых источников, из которых 34 - на русском и 273 - на иностранном языке.
ОСНОВНОЕ СОДЕРЖАНИЕ РАБОТЫ
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Культивирование ММСК. Объектом исследования являлись культуры мезенхимальных стромальных клеток костного мозга человека, любезно предоставленные д.б.н. Романовым Ю.А. (Научно-практическая лаборатория стволовых клеток человека НИИ РКНПК Росмедтехнологий). В качестве среды для поддержания роста ММСК использовали среду DMEM-LG с концентрацией глюкозы 1 г/л (Gibco, США), содержащую 2 мМ глутамина (MP Biomedicals, США), 1 мМ пирувата натрия, 12,5 мМ HEPES-буфера (Gibco или Sigma, США), 100 ед/мл пенициллина и 100 г/мл стрептомицина (Биолот, Россия), 10% эмбриональной телячьей сыворотки (Gibco, США). Замену среды проводили каждые 3-4 дня. Культивирование в стандартных условиях проводили при 37оС в СО2-инкубаторе (Sanyo, Япония).
Моделирование эффектов микрогравитации. Для моделирования эффектов микрогравитации использовали Устройство для рандомизации положения - Random Positioning Machine (dRPM, Dutch Space, Leiden, The Netherlands). В процессе работы RPM происходит рандомизация положения объекта относительно вектора гравитации за счет разнонаправленного вращения двух взаимоперпендикулярных рамок, к которым крепится платформа с экспериментальными образцами, за счет чего имитируется эффект снижения влияния силы тяжести на культуру клеток [van Loon J.J.W.A., 2007]. В результате испытаний был выработан оптимальный набор предварительно устанавливаемых параметров вращения симуляции микрогравитации (100 радиан в мин для внешней рамки и 80 для внутренней, при активной функции смены направления вращения).
Индукция и детекция остеогенной и адипогенной дифференцировки ММСК костного мозга человека. Индукцию остеогенной и адипогенной дифференцировки ММСК проводили по общепринятым методикам [Jaiswal N. et al., 1997; Janderova L. et al., 2003]. Эффективность дифференцировки оценивали в случае остеогенеза - по активации экспрессии щелочной фосфатазы и накоплению минерализованных компонентов матрикса; в случае адипогенеза - по накоплению липидных капель в цитоплазме клеток и образованию кластеров адипоцитов.
Иммунофенотипическая характеристика клеток проводилась методом проточной цитофлуориметрии с помощью моноклональных антител (Beckman Coulter, Франция или Biolegend, США), меченых FITC (CD34, CD11b, CD90, CD44, HLA A,B,C, CD49b, CD54) и PE (CD45, CD29, CD73, CD105, CD106), на приборе Epics XL, Beckman Coulter, США.
Флуоресцентные исследования. Изменения в структуре белков цитоскелета ММСК: F-актина, -тубулина и винкулина оценивали с помощью метода флуоресцентной микроскопии на микроскопе Axiovert 25 (Carl Zeiss, Германия), оснащенном сопряженной с ним цифровой черно-белой видеокамерой AxioCam HRm. После различных сроков экспозиции на RPM (30 минут, 6 часов, 24 часа, 48 часов, 120 часов и 24+24 часа ре-адаптации в статических условиях) клетки отмывали от среды, после чего фиксировали ледяным метанолом в течение 5 минут с последующей отмывкой. Для предотвращения неспецифического связывания антител, перед окрашиванием клетки инкубировали при комнатной температуре в течение 1 часа в 1% растворе БСА. Для визуализации структуры фибриллярного актина использовали стандартный краситель TRITC-фаллоидин в концентрации 50 мкг/мл (Sigma, США). Структуру микротрубочек цитоскелета оценивали путем окрашивания клеток в течение 1 часа антителами против -тубулина (Sigma, США), коньюгированными с Cy3 в разведении 1:200, соответственно. Клетки инкубировали с антителами к винкулину (Chemicon, США), в разведении 1:100 в течение 2 часов. После инкубации клетки отмывали от избытка первичных антител 3 раза PBS и инкубировали 1 час с вторичными антителами, коньюгированными с FITC (Jackson Immunoresearch, США).
Анализ уровня экспрессии генов.
Выделение тотальной РНК, обратная транскрипция. Выделение тотальной РНК производили с помощью набора RNeasy PLUS mini kit (Qiagen, США). После этапа культивирования клетки снимали с подложки, отмывали, питательная среда удалялась центрифугированием, а полученные образцы консервировались при -35 оС с реактивом RNAlater (Qiagen, США). Согласно инструкции производителя, клетки (250 – 500 тыс) после удаления стабилизирующего реагента RNAlater центрифугированием (5 минут при 1000 об/мин) инкубировали на шейкере в лизирующем буфере RTL (поставляемым в наборе) в течение 10 минут при комнатной температуре. Далее клеточный лизат переносили на колонки Qiashredder (Qiagen, США) и подвергали центрифугированию при 14000 об/мин в течение 2 минут, после чего к гомогенату клеток добавляли равное количество 70% этанола. Полученный раствор переносили на сорбционные колонки (поставляется в комплекте) и центрифугировали 30 сек при 10000 об/мин, супернатант удалялся. Далее производилась последовательная отмывка сорбционных колонок буферами RW1 (700 мкл) и RPE (500 мкл) с последующим центрифугированием 30 сек при 10000 об/мин. РНК смывали с колонок 50 мкл деионизированной воды. Полученные образцы РНК замораживали и хранили при -35 оС. Реакцию обратной транскрипции и получение кДНК на основе ранее выделенной РНК производили с помощью набора QuantiTect Reverse Transcription Kit (Qiagen, США), согласно инструкции производителя.
ПЦР с детекцией продуктов в режиме реального времени. Для определения уровня экспрессии генов с помощью ПЦР с детекцией продуктов в режиме реального времени с помощью прибора MX3000P (Stratagene, США) применялся набор реактивов для проведения ПЦР-РВ в присутствии красителя SYBR Green 1, содержащий рекомбинантную ДНК-полимеразу HotStarTaq, смесь дезоксирибонуклеотидов и флуоресцентный краситель SYBR Green (Синтол, Россия). Количество кДНК на одну реакцию составляло 0,2 – 0,4 мкг. Конечный объем реакции составлял 25 мкл. Для определения уровня экспрессии исследуемых генов применялись оптимизированные для метода Real-Time PCR пары праймеров QuantiTect Primer Assay (Qiagen, США) для каждого исследуемого гена: GAPDH – кат. № QT01192646, TBP - кат. № QT00000721, RHOА – кат. № QT00044723, ROCK - кат. № QT00034972, CFL – кат. № QT00201348, VCL – кат. № QT00078302, TUBB – кат. № QT00089775, ACTB – кат. № QT00095431, ACTG – кат. № QT00996415, ACTA – кат. № QT00199815, RUNX2 – кат. № QT00020517, PPARG – кат. № QT00029841, BGLAP – кат. № QT0022771, COL1A1 – кат. № QT00037793, ALPL – кат. № QT00012957, OMD – кат. № QT0015953.
В качестве хаускиппинг-генов для определения относительного значения изменения уровня экспрессии изучаемых генов, были использованы гены глицеральдегид-3-фосфат дегидрогеназы (GAPDH) и TATA-домен связывающий протеин (TBP), так как их значение остается высококонсервативным для клеток костного мозга при различных условиях [Vandesompele J. et al. 2002]. При анализе данных разница в значении СТ 3.5 единицы соответствовала изменению уровня экспрессии в 10 раз, согласно указаниям производителя прибора. Специфичность проводимой реакции определялась с помощью анализа кривой плавления продуктов амплификации в диапазоне от 55 оС до 95оС с шагом 1 градус. Количество ПЦР-реакций при анализе уровня экспрессии каждого из исследуемых генов составляло не менее 15 - 20 повторов в рамках одного эксперимента. Профилирование экспрессии генов - маркеров стволовых клеток человека с помощью наборов Stem Cell RT Profiler™ PCR Array. Система анализа экспрессии Stem Cell RT Profiler™ PCR Array (SABioscienses, США) позволяет изучать экспрессию 84 генов клеток человека, вовлеченных в идентификацию, рост и дифференцировку стволовых клеток, с помощью метода ПЦР с детекцией продуктов в режиме реального времени. В набор включены: гены, специфичные для стволовых клеток человека, гены, кодирующие белки, участвующие во внутриклеточной сигнализации и регуляции дифференцировки. Специфичные праймеры исследуемых генов и контроли качества реакции иммобилизованы в ячейках 96-ти луночных планшетов, адаптированных для использования с прибором MX3000P (Stratagene, США). Для проведения реакции ПЦР использовались наборы реагентов RT2-Real Time SYBR Green / ROX PCR master mix. Конечный объем реакционной смеси, согласно инструкции производителя, составлял 25 мкл. Количество кДНК в пересчете на одну реакцию – 0,2 – 0,4 мкг. Реакционная смесь переносилась в планшеты с иммобилизованными и лиофилизированными праймерами. После чего проводилась ПЦР с детекцией продуктов в режиме реального времени. Для нормализации результатов исследования использовались 5 хаускиппинг-генов: B2M, HPRT1, RPL13A, GAPDH, ACTB, входящие в состав набора.
Статистическая обработка данных. Статистическую обработку результатов проводили с использованием методов вариационной статистики программы “Excel” для Windows XP. Статистическая обработка данных ПЦР с детекцией продуктов в режиме реального времени осуществлялась с помощью программы REST 2009 v. 2.0.12, основные алгоритмы которой изложены в работе [Pfaffl M. et al., 2002]. Обработка данных профилирования экспрессии генов - маркеров стволовых клеток человека с помощью наборов Stem Cell RT Profiler™ PCR Array осуществлялась с помощью программного обеспечения, доступного на сайте производителя: http://www.sabiosciences.com/pcr/arrayanalysis.php. Межгрупповые отличия считали достоверными при p0,05.
ОСНОВНЫЕ РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ И ИХ ОБСУЖДЕНИЕ
Проведенная морфологическая, иммунофенотипическая и функциональная характеристика клеток костного мозга человека, основанная на общепринятых критериях оценки [Тепляшин А.С. и др., 2005; Паюшина О.В. и др., 2006; Pittenger M.F. et al., 1999; Conget P.A., Minguell J.J., 1999; Zuk P.A. et al., 2001; Majumdar M.K. et al., 2003; Wagner W. et al., 2005; Dominici M. et al., 2006], позволяла заключить, что они могут быть отнесены к ММСК, так как имеют типичную морфологию; на протяжении ряда пассажей экспрессируют характерные поверхностные антигены (CD90, CD105, CD73, CD29, CD44); не экспрессируют маркеры гематогенного ряда (CD11b, CD34, CD45); способны к дифференцировке в клетки мезенхимальной природы (остеобласты и адипоциты), сохраняя эту способность при длительном субкультивировании, что давало возможность использовать данные клетки в дальнейших экспериментах (Рис.1-2.).

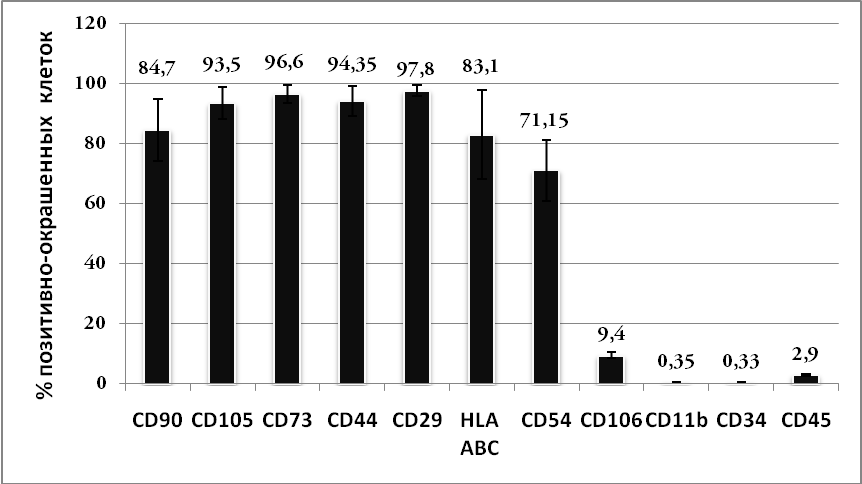
Рис. 1. Иммунофенотипическая характеристика ММСК костного мозга человека. А – иммунофенотип ММСК 2 пассажа, Б – иммунофенотип ММСК 7 пассажа. (M±m).
    | Рис. 2. Дифференцировка ММСК человека в остеогенном и адипогенном направлении. A – единичные клетки, экспрессирующие щелочную фосфатазу в культуре интактных ММСК, Б – активность ЩФ в ходе 10-суточной остеодифференцировки ММСК, В – отложения солей кальция в ходе 20-суточной остеодифференцировки ММСК, Ув. 100x. Г – накопление и слияние в цитоплазме клеток липидных капель в ходе 20-суточной адиподифференцировки ММСК Ув. 400x. |
Влияние моделирования эффектов микрогравитации на структуры цитоскелета ММСК и их способность к адгезии.
Среди основных структур цитоскелета клетки наибольший интерес исследователей привлекает его актиновая составляющая, как наиболее лабильная и быстро реагирующая на воздействия различного характера. Окрашивание фибриллярных актиновых волокон с помощью TRITC-фаллоидина позволило нам выявить изменения в их организации уже после 30 минут пребывания культуры ММСК в условиях моделирования эффектов микрогравитации с помощью RPM. Если в обеих контрольных группах клетки имели характерную пространственную исчерченность актиновых волокон, с хорошо организованными стресс-фибриллами, то в опытной группе, наряду с нормальными, появлялись отдельные клетки с аморфным распределением актина во внутриклеточном пространстве (Рис.3.). Зачастую, подобные изменения структуры F-актина выражались в его перераспределении от краев к центру клетки. При этом в пространстве клетки отчетливо выявлялись отдельные шарообразные «глобулы» F-актина, а сами клетки теряли актиновые стресс-фибриллы. После 6-часовой экспозиции клеток на RPM количество клеток с подобными нарушениями в организации актинового цитоскелета возрастало (Рис.3.). Анализ микрофотографий показал, что после 24 часов моделирования эффектов микрогравитации с помощью RPM изменения в структурной организации F-актинового цитоскелета были более отчетливыми и выражались уже в его перераспределении от центра к краям клетки, в результате чего в перинуклеарном пространстве клетки возникали характерные «пустоты». Интересно, что в это же время в культуре ММСК наблюдалось появление открепленных клеток (Рис.4.). Аналогичные эффекты отмечались и после 48 часов воздействия (Рис.3.), при этом количество открепившихся клеток во флаконах возрастало (Рис.4.). После 120 часов пребывания культуры ММСК в условиях моделированной микрогравитации клетки частично или полностью восстанавливали архитектонику актинового цитоскелета (Рис.3.). Кроме того, было выявлено, что на данном этапе культивирования открепление клеток прекращалось (Рис.4.). Такая же картина наблюдалась и после 24-часовой ре-адаптации клеток в статических условиях после того, как они в течение 24 часов подвергались моделированиию эффектов микрогравитации на RPM (Рис.3.). Подобное восстановление исходной микроархитектоники актинового цитоскелета при достаточно продолжительном пребывании клеток в условиях моделированной микрогравитации, по-видимому, отражает процесс адаптации клеток-предшественников к измененным гравитационным параметрам их среды обитания. Подтверждением этому предположению служит наблюдаемый факт аналогичного восстановления структуры актинового цитоскелета при возвращении клеток в стандартные статические условия, а также прекращение процесса открепления клеток в культуре.
Белок фокальных контактов винкулин связан с актином в сайтах фокальной адгезии и участвует в процессах адгезии клеток к субстрату [Miyamoto et al., 1995; Aplin et al., 1998]. Этот белок ко-локализован с кластерами интегринов в тех участках, где последние прикрепляются к внеклеточному матриксу. Выявление фокальных контактов у клеток-предшественников с помощью иммуноспецифичных антител против винкулина показало, что их структура также реагирует на изменение гравитационных условий. Характерным проявлением влияния моделирования эффектов микрогравитации в данном случае было постепенное перераспределение сайтов фокальной адгезии от равномерно покрывающих всю поверхность, до расположенных преимущественно в перинуклеарном пространстве клетки (Рис.5.). Этот процесс начинался уже через 6 часов после начала экспериментального воздействия и достигал максимума на конечных экспозициях (48 и 120 часов). В течение всего периода наблюдения у ММСК происходила прогрессивная потеря фокальных контактов на периферических участках клеток. Таким образом, флуоресцентная микроскопия позволила нам также выявить модификацию структуры винкулиновых сайтов адгезии у ММСК. Динамика этих изменений вполне согласуется с описанным выше процессом ремоделирования и восстановления структуры актиновых волокон у ММСК в течение пребывания клеток в условиях моделированной микрогравитации.
Используя флуоресцентную микроскопию, нам не удалось обнаружить выраженных нарушений в структуре микротрубочек цитоскелета, окрашенных специфичными антителами к -тубулину, ни на одном из исследуемых сроков экспозиции ММСК на RPM (Рис.6.). Полученные результаты могут, с одной стороны, свидетельствовать о том, что тубулиновый компартмент цитоскелета является наименее чувствительной к гравитации структурой цитоскелета клетки у клеток-предшественников. Однако нельзя исключать и того, что выбранные нами временные рамки исследования недостаточны для детекции потенциально возможных изменений.
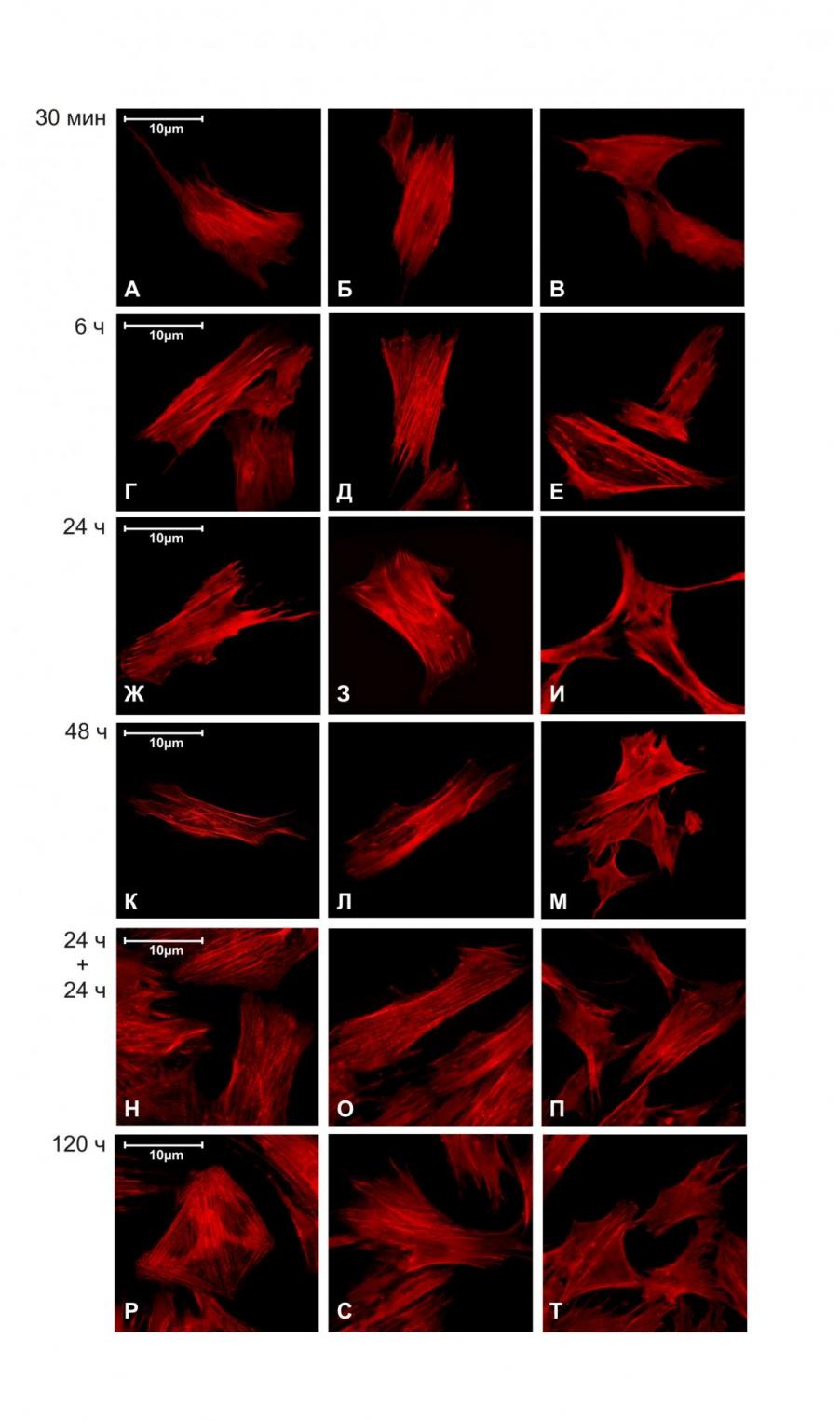

Рис.4. Влияние моделирования эффектов микрогравитации на адгезию ММСК костного мозга человека. Стрелками указаны открепившиеся при моделировании эффектов микрогравитации клетки. Ув. х100.
Поскольку после 120 часов экспозиции ММСК на RPM адгезия клеток восстанавливалась, было интересно определить, за счет каких рецепторов адгезии этот процесс может быть реализован. Известно, что ММСК экспрессируют на своей мембране широкий спектр рецепторов адгезии [Pittenger M.F. et al., 1999; Conget P.A., Minguell J.J., 1999; Zuk P.A. et al., 2001; Majumdar M.K. et al., 2003], вовлеченных в различные аспекты жизнедеятельности стволовых клеток, среди которых интегрины привлекают наибольший интерес в качестве гравитационно-сенсорных рецепторов. Проведенные исследования показали, что количество клеток, экспрессирующих CD29, который является основным -интегрином ММСК, опосредующим адгезию клеток к значительному количеству белков внеклеточного матрикса, оставалось неизменным после 120 часов экспозиции на RPM. В это же время, происходила значительная активация экспрессии CD49b (2-интегрин, рецептор коллагена), в результате чего количество клеток, экспрессирующих CD49b, возрастало в 1,5 – 2 раза (Рис.7.), что согласуется с данными [Meyers V.E. et al., 2004].
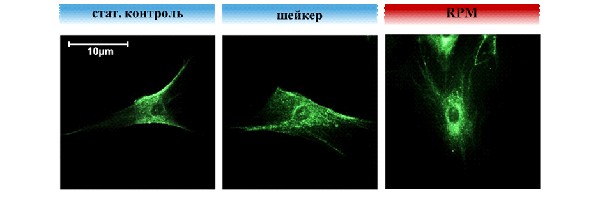

Рис.5. Пространственная организация сайтов фокальной адгезии, образованных винкулином, у ММСК после 120 часов культивирования в условиях моделирования эффектов микрогравитации с помощью RPM.


Рис.6. Пространственная организация -тубулинового цитоскелета ММСК на различных этапах экспозиции в условиях моделирования эффектов микрогравитации с помощью RPM.


Рис.7. Экспрессия рецепторов адгезии на поверхности ММСК после 120 ч экспозиции на RPM. А - влияние моделирования микрогравитации на экспрессию CD29 (1-интегрина), Б – влияние моделирования микрогравитации на экспрессию CD49b (2-интегрина). (M±m). Различия достоверны по критерию Манна-Уитни (*р<0,05). St – статический контроль, sh – шейкер.
Влияние моделирования эффектов микрогравитации на экспрессию избранных генов цитоскелета ММСК (с помощью полимеразной цепной реакции с детекцией продуктов в режиме реального времени).
Обнаруженные в нашей работе обратимые перестройки актинового цитоскелета ММСК позволили сделать предположение о том, что экспрессия некоторых генов, кодирующих белки цитоскелета, также может изменяться. Поэтому, следующим этапом данного исследования являлось изучение влияния моделированной микрогравитации на относительный уровень экспрессии ассоциированных с цитоскелетом структурных и регуляторных генов, в основном связанных с актиновым цитоскелетом: -актина (ACTA1), -актина (ACTB), -актина (ACTG1), винкулина (VCL), малой ГТФазы RhoA (RhoA), Rho-киназы (ROCK1), кофилина (CFL1) и -тубулина (TUBB). После 30 минут и 6 часов экспозиции клеток на RPM достоверных изменений в уровне экспрессии исследуемых генов обнаружено не было. Однако после 24, 48 и 120 часов выявились разнонаправленные изменения уровня экспрессии исследуемых генов относительно контроля. Наиболее выраженные изменения в уровне экспрессии ассоциированных с цитоскелетом генов наблюдались после 48 часов воздействия. Необходимо отметить, что клетки, помещенные на 24-часовую ре-адаптацию в статическое состояние, после 24-часовой экспозиции на RPM, характеризовались отсутствием достоверных различий в уровне экспрессии исследуемых генов по отношению к уровню в контрольной группе (Рис.8.). Анализ динамики изменения уровня экспрессии отдельных генов (Рис.8.) позволяет сделать вывод о том, что уровень экспрессии гена -актина начинает достоверно возрастать уже после 24 часов на RPM, а его максимальное значение наблюдается после 48 часового периода воздействия. На этом же сроке экспозиции на RPM уровень экспрессии -актина немного снижается, а малой ГТФ-азы RhoA, кофилина и -тубулина напротив повышается. После 120 часов моделированной микрогравитации сохраняется повышенным относительный уровень экспрессии -тубулина, кроме того наблюдается снижение относительного уровня экспрессии гена винкулина. Из приведенных данных видно, что достоверное изменение наибольшего числа исследуемых генов происходит после 48 часов воздействия (Рис.8.). Тем не менее, очевидно, что большинство из этих генов возвращаются на уровень экспрессии, сходный со статическим контролем, после 120-часовой экспозиции на RPM, причем, то же самое происходит и при 24-часовой ре-адаптации в статических условиях, что согласуется с наблюдаемыми структурными перестройками цитоскелета на соответствующих сроках экспозиции.

Рис.8. Влияние моделирования эффектов микрогравитации на относительный уровень экспрессии структурных и регуляторных генов белков цитоскелета ММСК.
А - изменение относительного уровня экспрессии отдельных генов цитоскелета в условиях моделирования эффектов микрогравитации (на сроках от 30 минут до 120 часов и 24 часовой реадаптации в статических условиях после 24 воздействия). Б – динамика изменения относительного уровня экспрессии генов цитоскелета при изучении влияния различных сроков экспозиции в dRPM (DutchSpace, Нидерланды). (M±SD).* p< 0,05; ** p< 0,01.
Влияние моделирования эффектов микрогравитации на экспрессию генов «стволовых маркеров» в ММСК (с помощью полимеразной цепной реакции с детекцией продуктов в режиме реального времени).
Так как в целом ряде работ было продемонстрировано, что ММСК экспрессируют некоторые маркеры эмбриональных стволовых клеток, наряду с генами, характерными для производных других зародышевых листков [Tremain N. et al., 2001; Silva W.A. Jr. et al., 2003; Pedemonte E. et al., 2007; Ulloa-Montoya F. et al., 2007], мы считали необходимым определить, как может меняться уникальный транскрипционный профиль ММСК на различных этапах экспозиции на RPM. «Stem Cell RT Profiler™ PCR Array» клеток человека (SABiosciences) позволяет изучать экспрессию 84 генов, вовлеченных в идентификацию, рост, самоподдержание и дифференцировку различных типов стволовых клеток человека. Производителем предлагается достаточно условная классификация данных генов, в зависимости от того, в какой клеточный процесс каждый из них вовлечен, и маркером какого клеточного типа он является (Табл.3). В результате проведенных исследований было установлено, что после 48 часов экспозиции культур ММСК на RPM уровень экспрессии только 9 генов изменяется. При этом, уровень экспрессии 3 генов возрастает (CCNA2, CD44, CDC2) и 6 снижается (ACTC1, ALDH2, CDH2, FZD1, ISL1, MME). Более выраженные изменения уровня экспрессии генов были обнаружены после 120 часов моделирования эффектов микрогравитации с помощью RPM. Среди 84 исследуемых генов, у 30 уровень экспрессии достоверно возрастал не менее чем в 5 раз (Табл.1), и у 24 - достоверно снижался (Табл. 2). Гены, уровень экспрессии которых увеличивался в наибольшей степени (не менее чем в 10 раз), обычно включали регуляторы клеточного цикла и пролиферации (CCND2, CCND1, CDС2, CDC42, HSPA9, FGF2, NOTCH2, NUMB), адгезии и межклеточного взаимодействия (CD44, CDH2, CTNNA1, CXCL12, GJA1, MME, NCAM1), внутриклеточной сигнализации и регуляции транскрипции (ADAR, APC, FZD1, NOTCH2, NUMB, BMP1, BTRC) (Табл.1). Гены, уровень экспрессии которых уменьшался в наибольшей степени (более чем в 10 раз), включали такие важные регуляторы, так называемого, самоподдержания стволовых клеток, а также маркеры коммитирования и клеточной дифференцировки как, ALPI, GDF2, GDF3, WNT-1, MYOD1, BMP-3, DHH, DLL3, FGF4, PDX1, NEUROG2, SOX-2, COL9A1, COL2A1, Т (Табл.2).
Таким образом, в ходе исследования было впервые установлено, что длительное моделирование эффектов микрогравитации с помощью RPM приводит к изменению паттерна экспрессии генов – «маркеров стволовых клеток человека» (Рис. 9.). При этом была выявлена интересная закономерность: при положительной регуляции преимущественно изменялась экспрессия генов, вовлеченных в процессы клеточной адгезии, пролиферации и внутриклеточной сигнализации, тогда как в случае отрицательной регуляции экспрессии - в основном гены, участвующие в клеточной дифференцировке. Кроме того, в отличие от генов цитоскелета, наибольшее изменение экспрессии генов «стволовых маркеров» наблюдалось после 120, а не 48 часов экспозиции. Интересно, что некоторые из исследуемых генов являются представителями антагонистических сигнальных путей (notch и wnt), и в условиях моделирования эффектов микрогравитации наблюдается преимущественная активация экспрессии первых и преимущественное подавление экспрессии вторых, что актуализирует необходимость дальнейшего изучения белковой активности компонентов данных сигнальных путей.
Таблица 1. Влияние моделирования эффектов микрогравитации (120ч) на экспрессию генов «стволовых маркеров» (положительная регуляция).
| Refseq | Ген | Регуляция (в разах) | Название продукта | Значение р |
| NM_001111 | ADAR | 200,96 | Adenosine deaminase, RNA-specific | 0,05 |
| NM_001135 | ACAN | 6,38 | Aggrecan | 0,0002 |
| NM_000690 | ALDH2 | 5,29 | Aldehyde dehydrogenase 2 family (mitochondrial) | 0,001 |
| NM_000038 | APC | 24,72 | Adenomatous polyposis coli | 0,0002 |
| NM_006129 | BMP1 | 10,51 | Bone morphogenetic protein 1 | 0,00001 |
| NM_033637 | BTRC | 12,59 | Beta-transducin repeat containing | 0,00005 |
| NM_053056 | CCND1 | 16,5 | Cyclin D1 | 0,002 |
| NM_001759 | CCND2 | 17,89 | Cyclin D2 | 0,002 |
| NM_000610 | CD44 | 25,47 | CD44 molecule (Indian blood group) | 0,0003 |
| NM_001786 | CDC2 | 19,71 | Cell division cycle 2, G1 to S and G2 to M | 0,00003 |
| NM_001791 | CDC42 | 10,44 | Cell division cycle 42 (GTP binding protein, 25kDa) | 0,00008 |
| NM_004360 | CDH1 | 6,25 | Cadherin 1, type 1, E-cadherin (epithelial) | 0,00003 |
| NM_001792 | CDH2 | 13,31 | Cadherin 2, type 1, N-cadherin (neuronal) | 0,0007 |
| NM_000088 | COL1A1 | 5,33 | Collagen, type I, alpha 1 | 0,0001 |
| NM_001903 | CTNNA1 | 23,66 | Catenin (cadherin-associated protein), alpha 1, 102 kDa | 0,001 |
| NM_000609 | CXCL12 | 18,91 | Stromal cell-derived factor 1 | 0,0007 |
| NM_001429 | EP300 | 7,02 | E1A binding protein p300 | 0,0008 |
| NM_002006 | FGF2 | 23,77 | Fibroblast growth factor 2 (basic) | 0,00001 |
| NM_015850 | FGFR1 | 7,65 | Fibroblast growth factor receptor 1 | 0,00001 |
| NM_003505 | FZD1 | 13,87 | Frizzled homolog 1 (Drosophila) | 0,00001 |
| NM_001527 | HDAC2 | 13,59 | Histone deacetylase 2 | 0,002 |
| NM_004134 | HSPA9 | 19,8 | Heat shock 70kDa protein 9 (mortalin) | 0,001 |
| NM_000902 | MME | 43,64 | Membrane metallo-endopeptidase | 0,000002 |
| NM_032188 | MYST1 | 5,18 | MYST histone acetyltransferase 1 | 0,0005 |
| NM_000615 | NCAM1 | 13,97 | Neural cell adhesion molecule 1 | 0,000002 |
| NM_017617 | NOTCH1 | 6,99 | Notch homolog 1, translocation-associated (Drosophila) | 0,0005 |
| NM_024408 | NOTCH2 | 25,53 | Notch homolog 2 (Drosophila) | 0,0005 |
| NM_003744 | NUMB | 12,08 | Numb homolog (Drosophila) | 0,0001 |
| NM_006086 | TUBB3 | 5,49 | Tubulin, beta 3 | 0,002 |
| NM_000165 | GJA1 | 39,97 | Gap junction protein, alpha 1, 43kDa | 0,00009 |
Таблица 2. Влияние моделирования эффектов микрогравитации (120ч) на экспрессию генов «стволовых маркеров» (отрицательная регуляция).
| Refseq | Ген | Регуляция (в разах) | Название продукта | Значение р |
| NM_001631 | ALPI | -398,70 | Alkaline phosphatase, intestinal | 0,00001 |
| NM_001201 | BMP3 | -161,18 | Bone morphogenetic protein 3 | 0,00001 |
| NM_000732 | CD3D | -9,14 | CD3d molecule, delta (CD3-TCR complex) | 0,0004 |
| NM_001768 | CD8A | -145,6 | CD8a molecule | 0,00001 |
| NM_004931 | CD8B | -21,64 | CD8b molecule | 0,00001 |
| NM_001844 | COL2A1 | -278,69 | Collagen, type II, alpha 1 | 0,00001 |
| NM_001851 | COL9A1 | -257,04 | Collagen, type IX, alpha 1 | 0,00001 |
| NM_021044 | DHH | -176,78 | Desert hedgehog homolog (Drosophila) | 0,00001 |
| NM_005618 | DLL1 | -19,19 | Delta-like 1 (Drosophila) | 0,0003 |
| NM_016941 | DLL3 | -315,72 | Delta-like 3 (Drosophila) | 0,0001 |
| NM_002007 | FGF4 | -372,86 | Fibroblast growth factor 4 | 0,00001 |
| NM_021784 | FOXA2 | -23,47 | Forkhead box A2 | 0,0001 |
| NM_016204 | GDF2 | -35,16 | Growth differentiation factor 2 | 0,00001 |
| NM_020634 | GDF3 | -27,97 | Growth differentiation factor 3 | 0,0001 |
| NM_000166 | GJB1 | -42,59 | Gap junction protein, beta 1, 32kDa | 0,0001 |
| NM_004004 | GJB2 | -5,39 | Gap junction protein, beta 2, 26kDa | 0,0001 |
| NM_000209 | PDX1 | -482,99 | Pancreatic and duodenal homeobox 1 | 0,00001 |
| NM_002202 | ISL1 | -15,77 | ISL LIM homeobox 1 | 0,00001 |
| NM_002275 | KRT15 | -13,57 | Keratin 15 | 0,0001 |
| NM_002478 | MYOD1 | -231,66 | Myogenic differentiation 1 | 0,00001 |
| NM_024019 | NEUROG2 | -157,86 | Neurogenin 2 | 0,00001 |
| NM_003106 | SOX2 | -888,87 | SRY (sex determining region Y)-box 2 | 0,00001 |
| NM_003181 | T | -27,52 | T, brachyury homolog (mouse) | 0,00001 |
| NM_005430 | WNT1 | -62,21 | Wingless-type MMTV integration site family, member 1 | 0,00001 |
Таблица. 3. Гены, входящие в состав Stem Cell RT Profiler™ PCR Array клеток человека, и их условная классификация (переведено с английского по данным сайта http://www.sabiosciences.com/rt_pcr_product/HTML/PAHS-405A.html).
| Регуляторы клеточного цикла | APC, AXIN1, CCNA2, CCND1, CCND2, CCNE1, CDC2, CDC42, EP300, FGF1, FGF2, FGF3, FGF4, MYC, NOTCH2, PARD6A, RB1 |
| Хромосомные Модуляторы | GCN5L2, HDAC2, MYST1, MYST2, RB1, TERT |
| Гены, регулирующие ассиметричное и симметричное деление клеток | DHH, NOTCH1, NOTCH2, NUMB, PARD6A |
| Маркеры самоподдержания | HSPA9, MYST1, MYST2, NEUROG2, SOX1, SOX2 |
| Цитокины и факторы роста | BMP1, BMP2, BMP3, CXCL12, FGF1, FGF2, FGF3, FGF4, GDF2, GDF3, IGF1 |
| Гены, регулирующие межклеточные взаимодействия | DHH, DLL1, GJA1, GJB1, GJB2, JAG1 |
| Молекулы клеточной адгезии | APC, BGLAP, CD4, CD44, CDH1, CDH2, COL9A1, CTNNA1, CXCL12, NCAM1 |
| Метаболические маркеры | ABCG2, ALDH1A1, ALDH2, FGFR |
| Маркеры клеточной дифференцировки: | |
| Маркеры ЭСК | ACTC1, ASCL2, FOXA2, PDX1 (IPF1), ISL1, KRT15, MSX1, MYOD1, T |
| Маркеры ГСК | CD3D, CD4, CD8A, CD8B, MME |
| Маркеры клеток мезенхимальной природы | ACAN (AGC1), ALPI, BGLAP, COL1A1, COL2A1, COL9A1, PPARG |
| Маркеры нервных клеток | CD44, NCAM1, OPRS1, S100B, TUBB3 |
| Сигнальные пути, необходимые для поддержания «стволовости»: | |
| Notch Pathway | DLL1, DLL3, DTX1, DTX2, DVL1, EP300, GCN5L2, HDAC2, JAG1, NOTCH1, NOTCH2, NUMB |
| Wnt Pathway | ADAR, APC, AXIN1, BTRC, CCND1, FRAT1, FZD1, MYC, PPARD, WNT1 |
 | 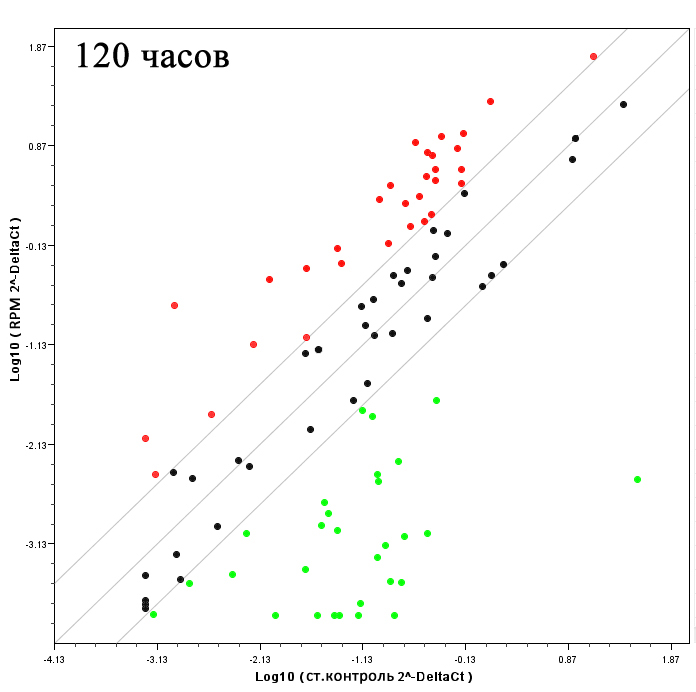 | Рис.9. Паттерн экспрессии генов – «стволовых маркеров» при моделировании эффектов микрогравитации. Красные точки – положительная регуляция экспрессии, зеленые – отрицательная регуляция, черные – отсутствие изменений в уровне экспрессии. |
Влияние моделирования эффектов микрогравитации на экспрессию основных маркеров остеогенной дифференцировки ММСК (с помощью полимеразной цепной реакции с детекцией продуктов в режиме реального времени).
Ранее было показано, что моделирование эффектов микрогравитации приводит к ингибированию остеогенной дифференцировки ММСК и преостеобластов [Zayzafoon M. et al., 2004; Meyers V.E. et al., 2004; 2005; Pardo S.J. et al., 2005; Dai Z.Q. et al., 2007; Patel M.J. et al., 2007; Gershovich J.G. et al., 2008; Pan Z. et al., 2008; Capulli M. et al., 2009; Huang Y. et al., 2009]. Некоторые авторы связывают данные изменения с нарушениями в актиновом цитоскелете клеток и активностью малых ГТФ-аз семейства Rho [Zayzafoon М. et al., 2004; Meyers V.E. et al., 2004; 2005]. В связи с этим, в данной работе мы исследовали экспрессию ключевых транскрипционных факторов (RUNX2, PPAR), регулирующих соответственно остеогенный и адипогенный дифференцировочный потенциал ММСК, а также основных маркеров их остеогенной дифференцировки ( ALPL, COL1A, BGLAP, OMD). Исследования показали, что после 10 суток индукции остеогенной дифференцировки ММСК в условиях моделирования эффектов микрогравитации с помощью RPM экспрессия ключевого мастер-транскрипционного фактора остеогенной дифференцировки ММСК - RUNX2 достоверно снижается по сравнению с клетками контрольной группы. Помимо подавления уровня экспрессии RUNX2 в ММСК наблюдается отрицательная регуляция экспрессии генов щелочной фосфатазы (ALPL) и белка адгезии остеобластов - остеомодулина (OMD) (Рис.10.). Интересно, что после 20 суток экспозиции ММСК на RPM в остеогенной среде экспрессия RUNX2 была по-прежнему снижена, тогда как экспрессия про-адипогенного транскрипционного фактора – PPAR - достоверно повышалась. На данном этапе в коммитированных к остеогенной дифференцировке клетках наблюдалась пониженная экспрессия OMD, тогда как экспрессия ALPL уже не отличалась от уровня в контрольной группе, кроме того на данном этапе достоверно активировалась экспрессия гена цепи коллагена 1 типа - COL1A (Рис.10.). Таким образом, в результате моделирования эффектов микрогравитации с помощью RPM происходит угнетение экспрессии генов, контролирующих процесс остеогенного коммитирования клеток. На начальных этапах моделирования эффектов микрогравитации подавление уровня экспрессии остеогенных генов-маркеров может происходить за счет снижения экспрессии ключевого транскрипционного фактора, регулирующего процесс остеогенной дифференцировки - Runx2, тогда как на поздних этапах ингибирование остеогенной дифференцировки клеток может быть реализовано посредством активации экспрессии такого про-адипогенного фактора транскрипции, как PPAR, известного тем, что он функционирует как доминантный негативный регулятор остеогенеза [Kang S. et al., 2007; Shockley K.R. et al., 2007]. Показанное нами возрастание уровня экспрессии PPAR согласуется с данными других исследователей [Zayzafoon М. et al., 2004; Meyers V.E. et al., 2005; Pan Z. et al., 2008; Huang Y. et al., 2009], в работах которых было установлено, что наряду с перестройками цитоскелета, это может быть основным фактором активации программы адипогенеза условиях моделированной микрогравитации или гипокинезии. Механизмы снижения экспрессии Runx2 могут быть обусловлены перестройками структуры актинового цитоскелета ММСК на начальных этапах моделирования эффектов микрогравитации, а также могут быть ассоциированы с ингибированием путей сигнальной трансдукции, связанных с сигнальными молекулами wnt и -катенином, поскольку показано, что под стимулирующим влиянием -катенина транскрипция Runx2 активируется [Gaur T. et al., 2005]. Ремоделирование актинового цитоскелета ММСК, наблюдающееся в ходе их остеогенной дифференцировки, свидетельствует о том, что при фенотипической трансформации клеток проявление экспрессии ряда характерных генов может зависеть от возможности нормальных перестроек актинового цитоскелета клеток, так как применение таких ингибиторов полимеризации актина, как цитохолазин D, приводит к уменьшению уровня экспрессии маркеров остеогенеза [Rodrguez J.P. et al., 2004].
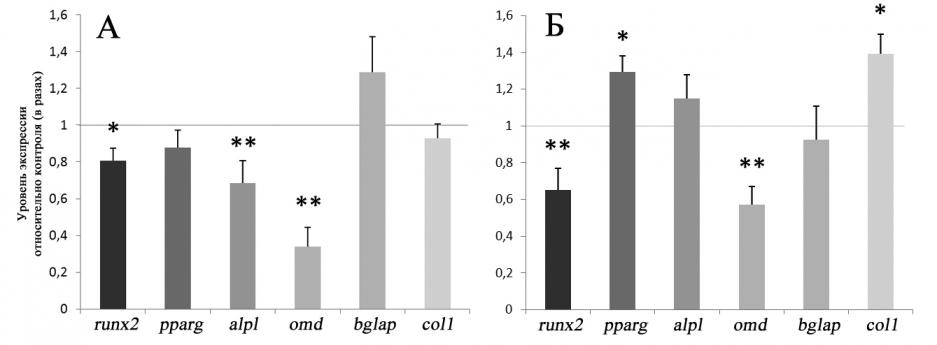
Рис.10. Влияние моделирования эффектов микрогравитации на относительный уровень экспрессии генов - маркеров остеогенного коммитирования ММСК.
А – Экспозиция ММСК в остеогенной среде на RPM в течение 10 сут. Б – Экспозиция ММСК в остеогенной среде на RPM в течение 20 сут. Уровень экспрессии генов в контрольной группе принят за единицу. (M±SD). * p< 0,01; ** p< 0,001.
ЗАКЛЮЧЕНИЕ
Проведенные исследования позволили установить, что мультипотентные мезенхимальные стромальные клетки костного мозга человека, принадлежащие к компартменту клеток-предшественников взрослого организма, представляют собой клеточную популяцию, которая восприимчива к изменению гравитации, по крайней мере, в условиях in vitro. При моделировании эффектов микрогравитации с помощью одной из современных наземных моделей – dRPM, в культурах ММСК имеют место обратимые перестройки актинового цитоскелета и происходит изменение адгезивных свойств клеток, которое сопровождается перераспределением фокальных контактов, образованных винкулином, и активацией экспрессии некоторых поверхностных рецепторов адгезии, в частности 2-интегрина. На фоне выявленных морфофункциональных и физиологических ответных реакций в культурах ММСК наблюдается изменение уровня экспрессии генов структурных и регуляторных белков актинового цитоскелета (АСТА1, ACTG, RHOA, CFL) клеток, уже на самых ранних этапах экспозиции на RPM, начиная с 24 часов, которое достигает максимума после 48 часов. Тем не менее, эти изменения являются временными и частично нивелируются на более поздних этапах воздействия (120 часов), а также быстро возвращаются на исходный уровень при ре-адаптации клеток в статических условиях. На более поздних этапах моделирования эффектов микрогравитации (120 часов), ответные реакции ММСК уже выражаются в изменении паттерна экспрессии генов – «маркеров стволовых клеток человека». В данном случае меняется экспрессия 54 генов, кодирующих целый спектр белков, вовлеченных в идентификацию, рост, самоподдержание и дифференцировку различных типов стволовых клеток человека. Моделирование эффектов микрогравитации приводит к активации или угнетению экспрессии разных генов, принимающих участие во внутриклеточной сигнализации, клеточной адгезии, регуляции пролиферации и дифференцировки клеток. И, наконец, коммитирование ММСК к остеогенезу в условиях длительного моделирования эффектов микрогравитации сопровождается подавлением экспрессии основных маркеров остеогенной дифференцировки клеток (ALPL, OMD), в том числе и ключевого мастер-транскрипционного фактора, контролирующего программу остеогенной дифференцировки ММСК – RUNX2. Необходимость дальнейшего изучения влияния реальной микрогравитации на клетки-предшественники очевидна в связи выполняемой ими функцией тканевой репарации, особенно учитывая данные об ингибирующем влиянии моделирования эффектов микрогравитации на остеогенный потенциал культивируемых ММСК, что представляет интерес не только в фундаментальном, но также в практическом плане. Работы в этом направлении позволят расширить фундаментальные представления о механизмах гравитационной и механической чувствительности клеток-предшественников взрослого организма и их возможной вовлеченности в развитие локальных клеточных реакций, развивающихся в костной системе в условиях микрогравитации, что, в свою очередь, будет способствовать разработке новых подходов к лечению и средствам профилактики состояний, вызванных уменьшением гравитационной нагрузки на костную ткань.
ВЫВОДЫ
- Моделирование эффектов микрогравитации с помощью RPM приводит к реорганизации актинового цитоскелета ММСК костного мозга человека (30 мин - 48 часов), которая является обратимой при более продолжительной экспозиции (120 часов) и при статической ре-адаптации в течение 24 часов, не оказывая влияния на организацию тубулинового цитоскелета.
- На начальных этапах моделирования эффектов микрогравитации (24 - 48 часов) ухудшается способность ММСК к адгезии. При длительной экспозиции ММСК на RPM (120 часов) адгезия клеток восстанавливается, этот процесс сопровождается перинуклеарным перераспределением винкулиновых сайтов фокальной адгезии на фоне увеличения доли клеток, несущих рецептор адгезии - 2-интегрин и возрастания уровня экспрессии генов, вовлеченных в клеточную адгезию (CD44, NCAM-1, СDH1, CDH2).
- Моделирование эффектов микрогравитации (24, 48, 120 часов) вызывает в ММСК транзиторное изменение уровня экспрессии генов, кодирующих белки актинового цитоскелета и ассоциированных с ним элементов (ACTA1, ACTG, RHOA, CFL1, VCL), которое частично компенсируется на конечных этапах (120 часов) и полностью при ре-адаптации клеток в статических условиях в течение 24 часов.
- При моделировании эффектов микрогравитации происходит достоверное изменение паттерна экспрессии «маркеров стволовых клеток» в ММСК человека: после 120 часов экспозиции экспрессия 54 генов из 84 исследуемых достоверно изменяется не менее чем в 5 раз, при этом уровень экспрессии 30 генов, включающих регуляторы клеточной пролиферации, адгезии и внутриклеточной сигнализации повышается, а 24 генов, большинство из которых контролирует клеточную дифференцировку – снижается.
- В условиях длительного моделирования эффектов микрогравитации в культурах, индуцированных к остеогенной дифференцировке ММСК достоверно подавляется экспрессия генов-маркеров остеогенеза – RUNX2, OMD (10, 20 сут), ALPL (10 сут) и активируется экспрессия анти-остеогенного гена - PPAR (20 сут).
- Обнаруженные структурные и молекулярно-генетические изменения свидетельствуют о наличии гравитационно-зависимых регуляторных механизмов, опосредующих как ранние, так и поздние ответные реакции клеток-предшественников на моделирование эффектов микрогравитации.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО МАТЕРИАЛАМ ДИССЕРТАЦИИ
Статьи:
- Gershovich P.M., Gershovich J.G., Buravkova L.B. Simulated microgravity alters actin cytoskeleton and integrin-mediated focal adhesions of cultured human mesenchymal stromal cells. // Jornal of Gravitational Physiology. 2008. Vol.15. №.1 Р. 203-204.
- Гершович П.М., Гершович Ю.Г., Буравкова Л.Б. Цитоскелет и адгезия культивируемых стромальных клеток-предшественников костного мозга человека при моделировании эффектов микрогравитации. // Цитология. 2009. Том.51. №11. С. 896-904.
- Гершович Ю.Г., Гершович П.М., Буравкова Л.Б. Влияние клиностатирования на культивируемые мезенхимальные стромальные клетки костного мозга человека. // Технологии живых систем. 2009. №2. С. 3-9.
- Буравкова Л.Б., Гершович П.М., Гершович Ю.Г., Григорьев А.И. Механизмы гравитационной чувствительности остеогенных клеток-предшественников. // ACTA NATURAE. 2010. Том. 2. №1 (4). С. 30-39.
- Гершович П.М., Гершович Ю.Г., Буравкова Л.Б. Роль мультипотентных мезенхимальных стромальных клеток костного мозга в адаптации клеток остеогенного дифферона к микрогравитации. // Российский физиологический журнал имени Сеченова. 2010. Том 96. №.4. С. 406-418.
Тезисы докладов:
- Гершович П.М., Гершович Ю.Г., Буравкова Л.Б. // Влияние клиностатирования на актиновый цитоскелет и адгезию мезенхимальных стромальных клеток-предшественников in vitro. Тезисы докладов. 11я международная Пущинская школа-конференция молодых ученых «Биология-наука XXI века». 2007. С.7.
- Гершович П.М., Гершович Ю.Г. // Применение метода ПЦР в реальном времени для оценки влияния 3D-клиностатирования на экспрессию некоторых генов цитоскелета мезенхимальных стромальных клеток-предшественников человека in vitro. Сборник тезисов. 7я Конференция молодых учёных и специалистов, аспирантов и студентов, посвящённая дню Космонавтики. Москва. 2008. С. 16-17.
- Гершович Ю.Г., Гершович П.М., Буравкова Л.Б. // Гравирецепция в культуре мезенхимальных клеток-предшественников. Тезисы докладов. 12я международная Пущинская школа-конференция молодых ученых «Биология-наука XXI века». 2008.С.172.
- Гершович П.М. // Реорганизация актинового цитоскелета и уровень экспрессии цитоскелетных генов в условиях моделированной микрогравитации. Сборник тезисов. 8я Конференция молодых учёных и специалистов, аспирантов и студентов, посвящённая дню Космонавтики. Москва. 2009. С. 16-17.
- Гершович П.М., Гершович Ю.Г., Жамбалова А.П., Буравкова Л.Б. // Экспрессия генов стволовых маркеров в культуре костномозговых стромальных клеток человека в условиях моделированной микрогравитации. Тезисы докладов. 14я международная Пущинская школа-конференция молодых ученых «Биология-наука XXI века». 2010. С. 203.
- Gershovich P.M., Gershovich J.G., Buravkova L.B. // The effects of 3D-clinorotation on cytoskeleton structures of mesenchymal stem cells derived from human bone marrow. Book of abstracts. International symposium «Biological motility»: Achievements and Perspectives. Pushchino. Part 2. 2008. P.218.
- Gershovich P.M., Gershovich J.G., Buravkova L.B. // Simulated microgravity alters actin cytoskeleton and integrin-mediated focal adhesions of cultured human mesenchymal stromal cells. Book of abstracts. 29th Annual International Physiology Meeting. Angers. France. 2008. P.143-144.
- Gershovich P.M., Gershovich J.G., Buravkova L.B. // Simulated microgravity and the pattern of actin and associated proteins gene expression in cultured human mesenchymal stromal cells. Book of abstracts. 17th Humans in Space Symposium. Moscow. 2009. P. 46.
- Gershovich P.M., Gershovich J.G., Buravkova L.B. // The role of cytoskeleton in functioning of cultured MSC during simulated microgravity. Book of abstracts. French-Russian-Belarusian Conference «Neurovascular impairment induced by environmental conditions: molecular, cellular and functional approach». Angers. France. 2010.