

Влияние интенсивной мышечной нагрузки на морфофункциональное состояние репродуктивных органов с а мок крыс
На правах рукописи

Самойленко Валерий Юрьевич
Влияние интенсивной мышечной нагрузки на
морфофункциональное состояние репродуктивных
органов самок крыс
03.03.01 – физиология
Автореферат
диссертации на соискание ученой степени
кандидата биологических наук
Майкоп – 2012
Работа выполнена на кафедре анатомии, физиологии и гигиены человека
ФГБОУ ВПО «Ставропольский государственный университет»
| Научный руководитель: | Беляев Николай Георгиевич доктор биологических наук, доцент |
| Официальные оппоненты: | Боташева Татьяна Леонидовна доктор медицинских наук, профессор ФГБУ «Ростовский научно-исследовательский институт акушерства и педиатрии» Министерства здравоохранения и социального развития Российской Федерации, главный научный сотрудник. Артемьева Надежда Константиновна доктор биологических наук, профессор, ФГБОУ ВПО "Кубанский государственный университет физической культуры, спорта и туризма", заведующий кафедрой биохимии и естественнонаучных дисциплин. |
| Ведущая организация: | ФГБОУ ВПО «Астраханский государственный университет» |
Защита состоится «30» мая 2012 г. в 16.00 часов на заседании диссертационного совета Д 212.001.07 в Адыгейском государственном университете по адресу: 385000, Республика Адыгея, г. Майкоп, ул. Пионерская, 260, конференц-зал научной библиотеки АГУ.
С диссертацией можно ознакомиться в научной библиотеке Адыгейского государственного университета, с авторефератом на сайте ВАК при Министерстве образования и науки РФ http: //www.vak.ed.gov.ru и на сайте Адыгейского государственного университета http: //www.adygnet.ru
Автореферат разослан «28» апреля 2012 г.
![]()
Ученый секретарь диссертационного совета
кандидат биологических наук, доцент Н.Н. Хасанова
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность исследования. Вопрос о влиянии мышечных нагрузок на женский организм продолжает привлекать внимание исследователей (Ягунов С.А., 1969; Соболева Т.С., 1996; Артемьева Н.К., 2000, 2008; Сазыкина Е.И. с соавт., 2001; Ниаури Д.А. с соавт., 2003; Зырянова Е.А., 2006; Литисевич Л.В., 2006; Юрчук О.А., 2007; Зырянова Е.А., 2008; Бердичевская Е.М., 2009; Гавердовский Ю.К., 2009; Кудря О.Н., 2011; Александрова Л.И. с соавт., 2011). Но при этом приходится констатировать явно недостаточный уровень знаний об особенностях адаптации женского организма к интенсивным, часто экстремальным, тренировочным и соревновательным нагрузкам, характерным практически для всех видов спорта (Граевская Н.Д., 1987; Зелинский О.О., 2005; Карковцева И.А., 2007; Дерех Э.К., 2008; Зырянова Е.А., 2008; Юров С.В., 2010; Панков Г.А., 2010).
Знания о функциональных резервах организма обычной женщины не вполне приемлемы к оценке функциональных возможностей спортсменок, демонстрирующих высокие спортивные результаты. Эти обстоятельства обусловливают необходимость проведения исследований в рамках проблематики морфологических и физиологических особенностей систем женского организма, которые определяют специфику их реакции на мышечные нагрузки.
В аспекте данной проблемы особый интерес представляет исследование влияния спортивной деятельности на женскую репродуктивную систему. Прежде всего, это определяется достаточно большим количеством данных, указывающих на высокую частоту нарушений в репродуктивной системе (Похоленчук Ю.Т., Свечникова Н.В., 1987; Раисова А.Т.,1990; Серова Л.В., 2003; Беляев Н.Г. и соавт., 2007, 2011; Зырянова Е.А.,2008; Писков С.И., 2008; Barbieri R. L.,1990; Bonen A.F., Haynes F.V.,1991), и отсутствием четкого понимания механизмов, посредством которых спортивная деятельность приводит к изменениям в функционировании отдельных звеньев данной системы.
В частности, вопросы развития гиперандрогении до конца не изучены, а имеющиеся данные в ряде случаев противоречивы. Так, ряд исследователей (Телунц А.В., 2001; Чернуха Г.Е., 2004) связывают гиперандрогению с гиперфункцией надпочечников, как следствие адаптации к интенсивным мышечным нагрузкам и высокому эмоциональному накалу спортивных соревнований. Другие (Сологуб В.В., 1989; Соболев Д.В., 1998; Липовка Л.В., 2004; Калинина Н.А., 2004) придерживаются мнения, что в спортивных секциях в процессе многолетних тренировок происходит постепенный отсев девочек феминного и концентрация маскулинного типов, тогда как маскулинизация является широко распространенным в популяции заболеванием - андрогинный синдром (Сологуб В.В., 1989; Соболев Д.В., 1998; Липовка Л.В., 2004). При данном заболевании отмечается блокирование синтеза кортизола в надпочечниках при одновременном повышении аналогов половых гормонов (Зырянова Е.А., Марова Е.И., Смоленский А.В., 2008). Подобное перераспределение стероидных гормонов дает женщинам преимущество в адаптации к физическим нагрузкам и возможность в последующем достигать высоких результатов в спорте.
Отсутствие единого мнения о причинах столь высокой частоты встречаемости гиперандрогении среди спортсменок в очередной раз подчеркивает актуальность подобных исследований. Немаловажным фактором является и то, что гиперандрогения регистрируется у спортсменок в различных формах ее проявления в 5 раз чаще, чем в популяции. В то время как гиперандрогения является одним из основных факторов, с которым связывают не только запаздывание полового созревания и различные формы нарушения менструальных функций, но и женское бесплодие (Сазыкина Е.Н. с соавт., 2001; Ниаури Д.А. с соавт, 2003;Чернухина О.В., 2003; Липовка Л.В., 2003, 2004; Зырянова Е.А., 2006; Литисевич Л.В., 2006).
Подтверждением актуальности диссертационного исследования является финансовая поддержка РГНФ (проект №07-06-18020с) выполненной работы. Изложенное выше и легло в основу данного диссертационного исследования, определив основные цели и задачи работы.
В аспекте рассматриваемой проблемы приемлемым является исследование влияния интенсивных мышечных нагрузок на женскую репродуктивную систему в экспериментальной модели с использованием самок крыс.
Грамотная экстраполяция результатов, полученных на самках крыс, позволит выстроить концепцию влияния интенсивных мышечных нагрузок на отдельные звенья репродуктивной системы и будет содействовать разработке практических мероприятий в физиологии женского спорта.
Цель исследования: выявить влияние интенсивной мышечной нагрузки на морфофункциональное состояние репродуктивных органов самок крыс.
Задачи исследования:
- оценить влияние интенсивных мышечных нагрузок на морфометрические показатели матки, маточных труб и морфофункциональное состояние яичников самок крыс;
- выявить роль яичников в гиперандрогенации организма самок крыс, адаптированного к выполнению мышечных нагрузок;
- изучить динамику тестостерона и эстрадиола самок крыс в условиях систематического выполнения интенсивных мышечных нагрузок.
Научная новизна.
С позиции комплексного подхода впервые проведена оценка гормонального статуса самок крыс в условиях их адаптации к интенсивной и продолжительной мышечной нагрузке. Осуществлены морфометрия матки, маточных труб и гистологические исследования яичников, миометрия и эндометрия матки с раннего пубертата до периода половой зрелости в условиях адаптации к мышечным нагрузкам. Установлена зависимость динамики концентрации гормонов от объема выполняемых мышечных нагрузок.
Обнаружено отставание в темпах увеличения массы матки, маточных труб и их линейных размеров у самок крыс, принуждавшихся к выполнению интенсивных мышечных нагрузок, по сравнению с животными контрольной группы. Данное отставание в массе и линейных размерах органов репродуктивной системы обусловлено сформировавшейся гиперандрогенией и гиперкортицизмом как результат адаптивных морфофункциональных изменений в организме под влиянием интенсивных мышечных нагрузок.
Выявлено, что по мере роста физической работоспособности самок крыс в репродуктивном периоде онтогенеза уменьшается количество яйцеклеток, проходящих полный цикл развития, и увеличивается процент атрезирующих фолликулов. Установленные морфофункциональные изменения в яичниках являются одним из факторов, обеспечивающих гиперандрогению женского организма в процессе увеличения объема тренировочных нагрузок и роста физической работоспособности.
Положения, выносимые на защиту.
1. Процесс адаптации самок крыс к интенсивным мышечным нагрузкам сопряжен с замедлением темпов увеличения массы матки, маточных труб и их линейных размеров.
- Гиперандрогения и гипертрофия пучковой и сетчатой зон надпочечников являются основополагающими факторами негативных изменений в органах женской репродуктивной системы, вызванными систематическим выполнением интенсивных мышечных нагрузок.
- По мере роста физической работоспособности самок крыс уменьшается количество яйцеклеток, проходящих полный цикл развития, и увеличивается процент атрезирующих фолликулов.
Теоретическая значимость. Результаты, полученные в процессе диссертационного исследования, являются дополнительным вкладом в теорию неспецифической адаптации женского организма к экстремальным факторам, к которым без сомнения можно отнести интенсивные мышечные нагрузки.
Материалы диссертационного исследования расширяют имеющиеся представления о морфофункциональных особенностях адаптации женского организма к интенсивным мышечным нагрузкам.
Определение гормонального статуса и данные морфометрической оценки матки и маточных труб самок крыс позволяют глубже понять механизмы адаптивных изменений в органах женской репродуктивной системы, которые в последующем могут явиться предпосылкой для развития патологий.
Результаты гистологических исследований яичников самок крыс дополняют имеющиеся данные об изменениях, протекающих в женском организме и обусловленных синдромом гиперандрогении.
Практическая значимость работы. Результаты, полученные при определении гормонального статуса организма, данные морфометрических, гистологических исследований матки, маточных труб и яичников самок крыс могут быть учтены при разработке научно-методических основ тренировочного процесса в женском спорте, а также использованы при ранней диагностике нарушений репродуктивной функции, обусловленной гиперандрогенией.
Апробация работы. Основные положения диссертации были доложены и обсуждены на межрегиональных научно-методических конференциях (Ставрополь, 2003-2009 гг.); на X Международной научно - практической конференции. (Ростов-на-Дону, 2007); Международной научно-практической конференции «Научное обоснование физического воспитания, спортивной тренировки и подготовка кадров по физической культуре и спорту» (Минск, 2009г.); VIII Всероссийской конференции «Нейроэндокринология – 2010» (Санкт-Петербург, 2010).
По теме исследования опубликовано 9 научных трудов, в т.ч. 2 статьи в рецензируемых журналах, определенных в Перечне ВАК при Министерстве образования и науки РФ.
Внедрение результатов Основные теоретические положения, результаты экспериментальных исследований настоящей работы используются в учебном процессе при чтении базовых дисциплин физиологического и морфологического блока в ФГБОУ ВПО «Ставропольский государственный университет» (355009, Россия, Ставропольский край, г. Ставрополь, ул. Пушкина, 1а), Ставропольском филиале Московского государственного гуманитарного университета имени М.А. Шолохова (355000, Россия, Ставропольский край, г. Ставрополь, ул. Доваторцев, 66г).
Теоретические положения и выводы настоящего исследования используются при чтении курса лекций по дисциплинам «Экологическая физиология», «Человек и его здоровье» и «Физиология человека» для студентов факультета физической культуры Ставропольского государственного университета.
ОРГАНИЗАЦИЯ И МЕТОДЫ ИССЛЕДОВАНИЯ
В исследованиях были использованы животные - самки крыс линии Вистар.
В соответствии с задачами исследования животные с возраста 2 месяцев принуждались к выполнению тренировочных нагрузок на протяжении 9 недель. Для моделирования мышечных нагрузок использовался тредбан. В используемой методике тренировок мы выделяли IV этапа тренировочных циклов: I – подготовительный (1-4 недели); II – разгрузочный (5-6 недели); III – цикл интенсивных нагрузок (7-8 недели); IV – цикл активного отдыха (9-я неделя).
На 2, 4, 6:, 8 и 9-й неделях тренировок осуществляли взятие крови для проведения биохимических анализов, органов (сердца, матки, надпочечников) для определения их массы, яичников и матки для гистологических исследований.
В крови проводили подсчет эритроцитов и определение гемоглобина автоматическим гематологическим анализатором Hema-Screen 18 (Гема-Скрин 18). Оценку гормонального статуса самок крыс осуществляли по концентрации в плазме крови 11- оксикортикостероидов (11-ОКС), эстрадиола и тестостерона. Определение концентрации половых гормонов в плазме крови осуществляли методом иммуноферментативного анализа с использованием наборов (ИФА) «Rat Elisa Assay Kit». Содержание 11-ОКС определяли флуорометрическим методом (Усватова И.Я., Панков Ю.А.,1976). Взвешивание сердца, матки и надпочечников с точностью до 0,01 мг осуществляли на электронных весах «ВЛТЭ-150».
При проведении гистологических исследований яичники фиксировали в 8% забуференном растворе формалина. Резку осуществляли на микротоме «ТЕХНОМ МЗП-1003» при помощи замораживающего устройства «ТЕХНОМ ОМТ-0228». Дальнейшая обработка гистологических препаратов осуществлялась в соответствии с рекомендациями /Автандилова Г.Г., 1990/.
При микроморфометрии использовали стандартный окулярмикрометр MOB - 1-16. Микрофотографирование осуществлялось при помощи комплекса, состоящего из микроскопа «Биомед-2» и цифровой фотокамеры «ДСМ-300».
Учитывая, что гормональный статус самок определяется фазой эстрального цикла, по достижению животными возраста 90 дней кровь для анализа брали в стадии Dioestus. Определение фазы эстрального цикла самок крыс осуществляляли методом влагалищного мазка (Кабак Я.М., 1968). Обработку полученных результатов исследования осуществляляли с использованием пакета прикладных компьютерных программ SPSS (версия 14) с применением критериев Стьюдента и Пирсона.
При статистической обработке результатов использовали пакет программ STATISTICA for Windows 5.0 Stat-Soft, Inc., включавший расчет средних значений и их ошибок с определением достоверности различий по t-критерию Стьюдента. Построение графических изображений проводилось с использованием стандартного пакета программ Microsoft Exel 7.0, таблиц – с помощью Microsoft Word 2000.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
Морфофункциональные изменения в организме самок крыс в процессе адаптации к физическим нагрузкам. Тренировка животных осуществлялась на протяжении 9 недель в соответствии с модифицированной методикой, описанной в разделе «Материалы и методы исследования». Животные экспериментальной группы ежедневно помещались в тредбан и осуществляли бег в соответствии с таблицей тренировки животных. Животные контрольной группы оставались в клетках.
По мере проведения эксперимента время бега экспериментальных животных постоянно увеличивалось, что свидетельствовало о росте работоспособности и выносливости.
Так, до начала эксперимента животные способны были выполнять бег по движущейся ленте в течение 2,7±0,30 часа. К моменту окончания эксперимента максимальная продолжительность бега для животных контрольной группы соответствовала 3,04±0,28 часа, а для самок экспериментальной группы – 4,2±0,21 часа, т.е. работоспособность животных в процессе тренировок возросла на 55%. Повышение работоспособности обусловлено определенными адаптивными изменениями в системах организма. В частности, отмечено увеличение количества эритроцитов.
Возрастание количества эритроцитов регистрировалось уже на 4-ой неделе тренировок и сохранялось на высоком уровне, по сравнению с животными контрольной группы, в последующие сроки эксперимента (табл. 1). К моменту окончания тренировочных циклов количество эритроцитов в крови самок крыс экспериментальной группы соответствовало 5,8±0,32 млн/мм. У животных контрольной группы - 4,6±0,45 млн/мм, т.е. тренировка животных обеспечивала повышение количества эритроцитов на 35%. Повышение количества эритроцитов у животных контрольной группы за аналогичный временной период составило 14%.
Достоверно более высокие величины отмечены и в содержании гемоглобина (табл.1).
В данном случае уровень гемоглобина в крови животных на 6-ой неделе эксперимента соответствовал - 14,0±0,60 гр%, на 9-ой неделе - 15,6±0,45 гр% при 8,6±0,30 гр% на 6-ой неделе и 9,1±0,32 гр% на 9-ой неделе соответственно у животных контрольной группы.
Таблица 1
Количество (М±m) эритроцитов и уровень гемоглобина в крови самок крыс
| Недели тренировок | Кол-во эритроцитов, млн/мм3 Контрольная группа | Кол-во эритроцитов, млн/мм3 Экспериментальная группа | Р2 | Содержание гемоглобина, гр% Контрольная группа | Содержание гемоглобина, гр% Экспериментальная группа | Р2 |
| I | 4,3±0,42 | 4,3±0,42 | - | 8,7±0,60 | 8,7±0,60 | - |
| 4 Р1 | 4,2±0,36 >0,5 | 4,6±0,60 >0,1 | >0,5 | 8,8±0.60 >0,5 | 8,9±0,40 >0,5 | >0.5 |
| 6 Р1 | 4,5±0,36 >0,1 | 5,6±0,60 >0,05 | >0.1 | 8,6±0,30 <0,05 | 14,0±0,60 <0,05 | <0.05 |
| 9 Р1 | 4,6±0,45 >0,05 | 5,8±0,32 >0,05 | <0.01 | 9,1±0,32 <0,05 | 15,6±0,45 <0,05 | <0.01 |
Примечание: Р1 – достоверность различий по сравнению с данными на начало эксперимента; I- данные на начало эксперимента; n - во всех случаях равно 7; Р2 – достоверность различий по сравнению с контрольной группой.
Увеличение количества эритроцитов не сопровождалось значительными изменениями реологических свойств крови, так как в данном случае регистрировалось и увеличение объема крови при незначительном изменении соотношения плазмы и форменных элементов крови. Т. е. происходили положительные адаптивные изменения в системе крови.
Таблица 2
Показатели (М±m) динамики массы сердца крыс в течение 9 недель тренировочного цикла
| Условия эксперимента | Масса сердца, мг/100 г | |||
| І | Через 4 недели | Через 6 недель | Через 9 недель | |
| Контрольная группа Р1 | 240±0,021 | 240±0,019 >0.5 | 230±0,017 >0.5 | 270±0,019 <0.01 |
| Экспериментальная группа Р1 | 240±0,021 | 260±0,020 >0.5 | 380±0,019 <0.01 | 580±0,040 <0.05 |
| Р2 | - | <0.01 | <0.01 | <0.05 |
Примечание: Р1 – достоверность различий по сравнению с данными на начало эксперимента; Р2 – достоверность различий по сравнению с контрольной группой. I- данные на начало эксперимента;
Положительные изменения были отмечены и в массе сердца (табл.2). Так, у самок экспериментальной группы незначительный прирост массы сердца в перерасчете на 100 грамм массы тела был отмечен уже через 4 недели тренировок. Высоко достоверное преобладание массы сердца животных экспериментальной группы регистрировалось с 6-ой недели систематического выполнения мышечных нагрузок и возрастало к моменту окончания эксперимента.
Высокая физическая работоспособность самок крыс и изменения в ряде вегетативных систем организма позволяют нам говорить о достижении определённого уровня тренированности животных.
Динамика 11-оксикортикостероидов и половых гормонов самок крыс в процессе тренировочных циклов. Наряду с морфологическими и функциональными изменениями в вегетативной системе организма, процесс адаптации связан и с изменением нейроэндокринного статуса организма самок крыс, принуждавшихся к выполнению мышечных нагрузок.
При исследовании гормонального статуса животных, начиная с возраста 90 дней, кровь для исследования брали в фазу Dioestrus. При этом необходимо отметить, что у самок экспериментальной группы в течение всего эксперимента не регистрировалось чёткой ритмики овариально-менструального цикла.
Учитывая, что гормонам коркового слоя надпочечников принадлежит ведущая роль в становлении адаптивных реакций (Розен В. Б., 1994; Hoekinds G., 1998) и то, что через всю гипоталамо-гипофизарно - адренокортикальную систему реализуется опосредованное влияние факторов окружающей среды на репродуктивную систему (Филаретов А. А., 1992; Viau V., 2002), осуществлялось определение уровня 11-ОКС в крови животных.
Таблица 3
Показатели (М±m) динамики концентрации 11-ОКС мкг/л в течение 9 недель тренировок
| Недели эксперимента | Условия эксперимента | Р2 | |
| Контрольная группа | Экспериментальная группа | ||
| I | 210,6±9,3 | 210,6±9,3 | - |
| 2 неделя Р1 | 215,3±7,4 >0,5 | 250,2±6,4 <0,001 | <0,001 |
| 4 неделя Р1 | 208,6±12,1 >0,5 | 270,6±8,1 <0,001 | <0,001 |
| 6 неделя Р1 | 197,0±9,8 >0,5 | 280,3±7,4 <0,001 | <0,001 |
| 8 неделя Р1 | 203,6±11,3 >0,5 | 269,2±8,7 <0,001 | <0,001 |
| 9 неделя Р1 | 205,2±7,1 >0,5 | 263,8±6,9 <0,001 | <0,001 |
Примечание: I - данные на начало эксперимента: n - во всех случаях равно 7; Р1 - достоверность различий по сравнению с данными на начало эксперимента; Р2 - достоверность различий по сравнению с контролем.
В результате проведенных исследований было установлено повышение концентрации 11-ОКС в крови самок экспериментальной группы (табл.3). В конкретных величинах концентрация 11-ОКС в плазме животных повысилась с 210,6±9,3 мкг/л до 263,8±6,9 мкг/л к моменту окончания эксперимента. При этом необходимо отметить фазовые изменения в динамике уровня 11-ОКС. В частности, пик повышения концентрации определяемого гормона регистрировался через 6 недель тренировок с последующим снижением на 8-ой и 9-ой неделях.
Снижение концентрации 11-ОКС на последующих этапах эксперимента мы связываем с повышением функциональных возможностей организма. Развитие тренированности ведет к уменьшению выраженности кортикальной реакции.
Повышение базального уровня 11-ОКС является следствием увеличения функциональных возможностей железы как результат структурно-приспособительной перестройки. Общим выражением подобных перестроек является возрастание массы железы.
В наших исследованиях масса надпочечников самок крыс (табл.4) в течение 9 недель эксперимента повысилась с 7,9±0,42 мг/100г до 13,0±0,50 мг/100г массы тела.
В контрольной группе колебания массы надпочечников в течение всего эксперимента не превышали 2%.
При динамической работе большой мощности гипертрофия надпочечников происходит за счёт прироста массы пучковой и сетчатой зоны. В то время как скоростная тренировка вызывает у крыс увеличение массы надпочечников за счёт мозгового слоя и клубочковой зоны (Виру А.А., Кырге П.К., 1983)
Таблица 4
Показатели (М±m) динамики изменения массы надпочечников в процессе тренировочного цикла
| Недели тренировок | Надпочечники, мг/100г | ||
| Контрольная группа | Экспериментальная группа | Р2 | |
| I | 7,9±0,42 | 7,9±0,42 | - |
| 4 Р1 | 8,0±0,11 >0,5 | 8,7±0,13 >0,1 | <0,001 |
| 6 Р1 | 7,9±0,81 >0,5 | 12,4±0,70 <0,001 | <0,001 |
| 9 Р1 | 8,0±0,39 >0,5 | 13,0±0,50 <0,001 | <0,001 |
Примечание: Р1 - достоверность различий по сравнению с контролем; Р2 - достоверность различий по сравнению с данными на начало эксперимента; I- данные на начало эксперимента.
Несомненно, гормоны пучковой и сетчатой зон надпочечников наиболее значимы для становления адаптивных реакций. В то же время отклонение от нормы в балансе кортикостероидов существенно влияет на половое развитие женского организма (Палади Г.А., Марку Г.А., Мукуца Э. В.,1978; Козлов В.М. и соавт.,1990; Тинников А.А., 1990; Подзолкова Н.М., 2006).
Так, высокая концентрация кортикостероидов как результат адаптивной гипертрофии надпочечников оказывает ингибирующее влияние на процесс полового созревания женской особи (Палади Т.А. и соавт., 1978; Тинников А.А., 1990). Влияние кортикостероидов на репродуктивную систему осуществляется через центральные и периферические звенья гипофизарно-яичниковой оси.
Необходимо отметить, что точкой приложения действия кортикостероидов является генетических аппарат клетки (Юдаев Н.А. и соавт., 1976). Поэтому нарушение баланса кортикостероидов может явиться причиной возникновения нарушений в репродуктивной системе не только на ранних этапах онтогенеза, но и в постнатальный период (Шишкина Г.Т., Дыгало Н.Н., 1993; Тарасенко Л.В. и соавт., 1995; Willig R., 1985; Hochberg Z. Et al, 1985; Rohde W., 1998).
Учитывая важную роль в развитии и функционировании женской репродуктивной системы, корковый слой надпочечников получил название «третьей гонады» (Кация Г.В., Тодуа Т.Н., 1991; Botella-Lustia J., 1954).
Таким образом, повышение концентрации кортикостероидов в процессе адаптации к интенсивным мышечным нагрузкам, возможно, и явилось одной из причин нерегистрируемой цикличности в функционировании половой системы самок крыс экспериментальной группы. Реакцию выключения половых циклов у самок следует рассматривать как особый вид адаптации к длительно действующему стрессирующему фактору.
С гормональной функцией коры надпочечников связано и секретирование половых гормонов. В наших исследованиях наряду с понижением концентрации эстрадиола регистрировалось достоверное повышение уровня тестостерона в крови животных экспериментальной группы.
Необходимо отметить, что в крови животных как контрольной, так и экспериментальной группы регистрировались фазовые изменения уровня половых гормонов (рис.1, 2). При этом колебания концентрации эстрадиола и тестостерона у животных контрольной группы определялись стадией полового развития.

Рис.1. Динамика концентрации тестостерона в течение 8 недель тренировок, нмоль/л. Примечание: I - данные на начало эксперимента; * - достоверность различий по сравнению с данными контрольной группы
В частности, высокие концентрации гормонов регистрировались в конце пубертатного периода (4-ая неделя эксперимента, возраст животных 84 дня) с последующим снижением на 8-ой и 9-ой неделях эксперимента - период репродуктивной зрелости (табл.5).
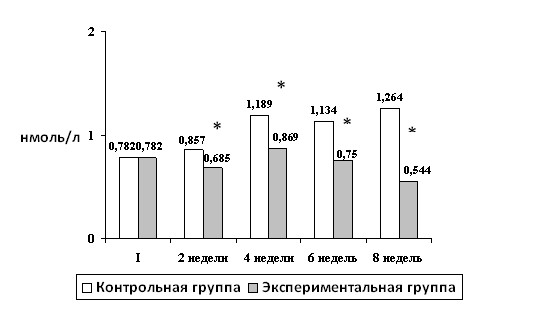
Рис.2. Динамика концентрации эстрадиола в течение 8 недель тренировок, нмоль/л. Примечание: I - данные на начало эксперимента; * - достоверность различий по сравнению с данными контрольной группы
Таблица 5
Показатели (М±m) динамики концентрации тестостерона и эстрадиола в течение 9 недель тренировок
| Недели эксперимента | Тестостерон, нмоль/л | Р2 | Эстрадиол, нмоль/л | Р2 | ||
| Контрольная группа | Эксперименталь- ная группа | Контрольная группа | Эксперименталь- ная группа | |||
| I | 3,1±0,09 | 3,1±0,09 | - | 0,782±0,023 | 0,782±0,023 | - |
| 2 Р1 | 3,3±0,12 >0,1 | 2,9±0,25 >0,1 | >0,1 | 0,857±0,031 >0,1 | 0,685±0,054 >0,1 | <0.02 |
| 4 Р1 | 5,2±0.08 <0,001 | 6,4±0,17 <0,001 | <0,01 | 1,189±0,075 <0.001 | 0,869±0,043 >0,1 | <0,001 |
| 6 Р1 | 4,7±0,22 <0,001 | 6,8±0,31 <0,001 | <0,001 | 1,134±0,046 <0,001 | 0,750±0,042 >0,1 | <0,001 |
| 8 Р1 | 3,7±0,29 <0,001 | 5,0±0,08 <0,001 | <0,01 | 1,264±0,085 <0,001 | 0,544±0,033 <0,001 | <0,001 |
| 9 Р1 | 3,8±0,34 <0,001 | 7,4±0,15 <0,001 | <0,001 | 1,089±0,094 <0,001 | 0,893±0,059 >0,1 | <0,001 |
Примечание: I - данные на начало эксперимента: n - во всех случаях равно 7; Р1 - достоверность различий по сравнению с данными на начало эксперимента; Р2 – достоверность различий по сравнению с контролем.
У животных экспериментальной группы прослеживается зависимость в динамике содержания гормонов от объема выполняемых нагрузок. Так, снижение концентрации определяемых гормонов регистрировалось в пике интенсификации тренировочного процесса (8-ая неделя) и повышение в период активного отдыха (9-ая неделя).
К этим факторам необходимо отнести гиперкортицизм и гиперандрогению как результат адаптивной гипертрофии надпочечников. Гиперандрогения у спортсменок - важная проблема женского спорта. Вопрос о ее причинах до сих пор дискутируется. В условиях продолжительного действия стресс-фактора гиперандрогения может быть обусловлена структурными и функциональными изменениями в яичниках.
Адаптация организма к интенсивным мышечным нагрузкам во многом обеспечивается изменением гормонального статуса. Прежде всего - повышением концентрации кортикостероидов и изменениями соотношения половых гормонов. Преимущественное насыщение организма мужскими или женскими половыми гормонами во многом определяет конечный биологический эффект. Андрогены, являясь антагонистами эстрогенов, опосредованно подавляют фолликулостимулирующую функцию гипофиза и непосредственно действуют на органы женской репродуктивной системы (Крупко-Большова А.Ю., 1990). При этом сформировавшиеся структурные гормональные перестройки могут сохранятся и после прекращения тренировок (Солодков А.С., 1990), а развивающиеся в следствие этого дисфункции репродуктивной системы могут оставаться и даже прогрессировать.
Морфофункциональная характеристика полового аппарата самок крыс в условиях адаптации к интенсивным мышечным нагрузкам. С целью установления особенностей становления органов репродуктивной системы в условиях гиперандрогении и гиперкортицизма, а также выявления возможно иных, помимо гипертрофии надпочечников, механизмов гиперандрогении осуществлялись морфометрические и гистологические исследования яичников, матки и маточных труб самок крыс.
Результаты морфологического исследования полового аппарата самок, подвергавшихся тренировочному циклу, позволили выявить ряд существенных изменений линейных показателей и показателей массы матки и маточных труб.
Прежде всего, существенное отставание в темпах увеличения их линейных размеров и массы прослеживалось у животных экспериментальной группы (рис. 3).
Так, линейные размеры исследуемых органов самок крыс контрольной группы в течение первых 4-х недель эксперимента возросли с 3,1±0,2 см до 5,2±0,4 см, т.е. прибавка составила более 67%.
У самок крыс, систематически выполнявших мышечные нагрузки, прибавка линейных размеров маточных труб за аналогичный временной период составила 6,4%. В конкретных величинах линейные размеры исследуемого органа за 4 недели эксперимента увеличились с 3,1±0,2 см до 3,3 ± 0,2 см. Прибавка составила всего 0,2 см. У животных контрольной группы за аналогичный временной период прибавка линейных размеров маточных труб составила 2,1см.
Линейные размеры исследуемых органов на 6-ой неделе эксперимента так же достоверно различались и составляли в контрольной группе 5,5 ±0,2 см, а в экспериментальной группе - 3,5±0,4 см.
Рис.3. Длина полового аппарата самок крыс в процессе тренировочного цикла, см.
Примечание: I - данные на начало эксперимента; 4-9 - недели эксперимента. * - достоверность различий по сравнению с данными контрольной группы
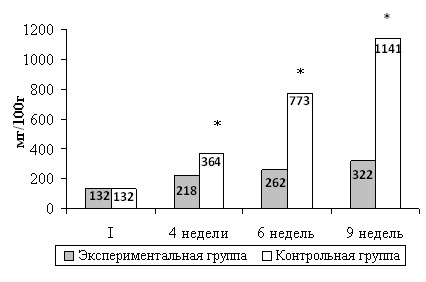
Рис.4. Масса матки самок крыс в процессе тренировочного цикла.
Примечание: I - данные на начало эксперимента; 4-9 - недели эксперимента. * - достоверность различий по сравнению с данными контрольной группы
К моменту окончания исследования линейные размеры маточных труб самок крыс контрольной группы на 37% превосходили линейные размеры маточных труб самок крыс экспериментальной группы и соответствовали 6,3±0,3 см и 4,0±0,2 см соответственно. Данные отличия были высоко достоверны (Р<0,001).
Аналогичная картина прослеживалась и в отношении массы матки (рис. 4). Так, через 4 недели эксперимента масса исследуемого органа у контрольных животных составляла 364±0,08 мг\100г, а у животных, принуждавшихся к регулярному выполнению мышечных нагрузок, 218±0,03 мг\100г; через 6 недель масса исследуемого органа у контрольных животных соответствовала 773±0,02 мг\100г, а у самок крыс экспериментальной группы - 262±0,03 мг\100г соответственно.
По окончанию эксперимента масса матки самок крыс контрольной группы более чем в 3,5 раза превосходила массу матки самок экспериментальной группы.
Визуально матка и маточные трубы животных экспериментальной группы тоньше, окружающая жировая ткань практически отсутствует (рис.5, 6).
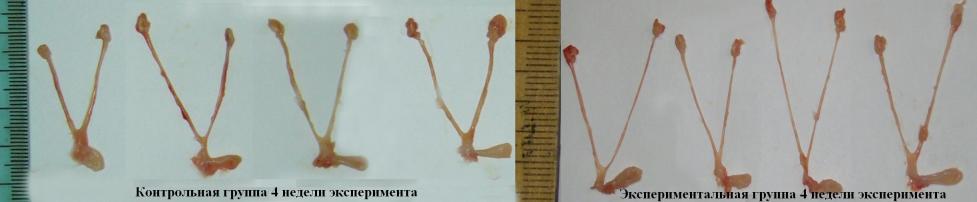
Рис.5. Общий вид репродуктивных органов у крыс контрольной и экспериментальной группы. Половой аппарат крысы (4 недели мышечной нагрузки)

Рис. 6. Общий вид репродуктивных органов у крыс контрольной и экспериментальной группы. Половой аппарат крысы (9 недель мышечной нагрузки)
Таким образом, можно заключить, что общая тенденция изменений в половом аппарате самок, принуждавшихся к выполнению интенсивных мышечных нагрузок, носит характер гипоплазии или дигенезии.
При гистологических исследованиях матки были выявлены существенные нарушения в структуре внутреннего и среднего слоёв матки.
Так, в миометрии матки регистрируются очаги склеротизации и гомогенизации мышц, т.е. явление некроза, характерное для эндометриоза. В основе лежит не только нарушение гормонального баланса, но и значительное снижение кровоснабжения гладкомышечной ткани матки - отрицательный перекрестный адаптивный эффект.
В эндометрии матки также регистрируются очаги некроза, частичное ороговение эпителия. Железы находятся в состоянии вакуольной дистрофии. Подобные изменения связаны с отсутствием желтых тел в яичниках, определяющих секреторную активность эпителиальных желез.
Обнаруженные структурные изменения в матке (гипоплазия, дистрофия миометрия - баллонная дистрофия) в последующем могут явиться причиной снижения возможности оплодотворённой яйцеклетки к имплантации, нарушения обменных процессов в организме плода, его гипотрофии, гипоплазии и невынашивания беременности.
Изменения в матке и яичниках, возможно, связаны с отрицательным адаптивным перекрестным эффектом. Так, в органах, непосредственно не задействованных в адаптации к конкретному возмущающему фактору, отмечается снижение кровоснабжения и, как следствие, недостаточное поступление энергетического и пластического материала. В конечном итоге, это может явиться причиной нарушения структуры тканей в данных органах.
Столь значительные морфофункциональные изменения в органах репродуктивной системы и гормональном статусе самок, возможно, связаны и с возрастом животных на момент начала тренировок. Самки крыс принуждались к выполнению мышечных нагрузок в «критический» период развития их репродуктивной системы (9 недель). Данный возрастной период характеризуется формированием овуляторной функции яичников, становлением качественной и количественной секреции половых гормонов. Нейроэндокринная система в этот период функционирует со значительным напряжением и особенно чувствительна к влиянию стресс-факторов, к которым с полной уверенностью можно отнести интенсивные мышечные нагрузки.
Гиперандрогения у спортсменок - важная проблема женского спорта. Вопрос о ее причинах до сих пор дискутируется. В условиях продолжительного действия стресс-фактора гиперандрогения может быть обусловлена структурными и функциональными изменениями в яичниках.
Так, у животных контрольной группы в возрасте 90 дней (4-я неделя эксперимента) в яичниках обнаруживаются желтые тела и яйцеклетки, находящиеся на разных стадиях созревания. Т.е. представленные результаты гистологических исследований свидетельствуют о полноценном функционировании репродуктивной системы самок крыс контрольной группы.
Наиболее ярко это проявляется в возрасте 130 дней (9-я неделя эксперимента). В данном случае регистрируется большое количество желтых тел, свидетельствующих о прохождении большим количеством яйцеклеток полного цикла развития (рис.7).
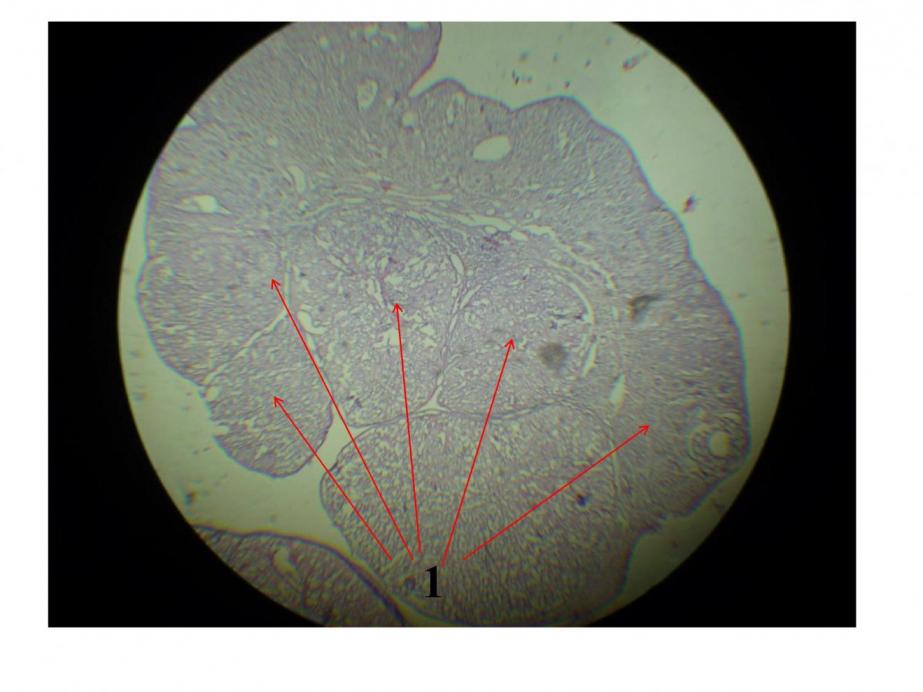
Рис.7. Яичник самок крыс контрольной группы животных (1-жёлтые тела) (9 недель эксперимента). Окраска гематоксилин - эозином. Увеличение 200.
У животных экспериментальной группы регистрируется незначительное количество желтых тел на 4-й неделе эксперимента и полное отсутствие их в момент окончания тренировочных циклов (рис.9). Но при этом в яичниках самок крыс, систематически выполнявших мышечные нагрузки, отмечается высокий процент атрезирующих вторичных и третичных фолликулов.
В атрезирующих фолликулах фолликулярные клетки способны перерабатывать андрогены в эстрогены только на начальных стадиях атрезии, а на стадии атретического тела и скопления интерстициальных клеток такая способность теряется, но сохраняется возможность секретировать тестостерон. Соответственно, высокий процент атрезирующих фолликулов может являться дополнительным фактором, обеспечивающим высокий уровень тестостерона в крови животных, принуждавшихся к выполнению мышечных нагрузок.

Рис.9. Яичник самок крыс экспериментальной группы животных (атрезия фолликулов с видом гиалиновых тел) (9 недель тренировок). Окраска гематоксилин - эозином. Увеличение 600.
Высокий процент атрезирующих фолликулов в яичниках животных экспериментальной группы следует расценивать как одну из форм адаптивных реакций, направленных на удовлетворение возросшей потребности организма в анаболических стероидах. Прежде всего, это связано с тем, что тестостерон яичников также, как и тестостерон надпочечников, способствует маскулинизации организма самок и, соответственно, лучшей адаптации к мышечным нагрузкам.
В итоге, можно заключить, что в основе гиперандрогении при мышечных нагрузках, с одной стороны, лежит повышение массы надпочечников и, как следствие, их секреторных возможностей, с другой - высокий процент атрезирующих фолликулов.
Таким образом, полученные данные демонстрируют то, что интенсивная мышечная нагрузка воздействует угнетающе на репродуктивную систему самок крыс и эти изменения могут быть связаны с отрицательным перекрестным эффектом - гиперкортицизмом и изменением соотношения эстрогенов и тестостерона.
ВЫВОДЫ
1. Адаптация самок крыс к интенсивным мышечным нагрузкам сопряжена с отставанием более чем в 3,5 раза в темпах увеличения массы матки, маточных труб, их линейных размеров на фоне гипертрофии сердечной мышцы при повышении количества эритроцитов, концентрации гемоглобина, что характерно для повышения выносливости и работоспособности организма.
2. Отставание в темпах увеличения массы полового аппарата у самок экспериментальной группы сопровождалась эозинофилией эндометрия, отёком и частичным склерозированием миометрия, замещением мышечной ткани соединительной и атрезией фолликулов в яичниках.
3. В процессе адаптации самок крыс к мышечным нагрузкам формируются механизмы, обеспечивающие длительное поддержание высокой концентрации тестостерона и других кортикостероидов наряду со снижением уровня женских половых гормонов, как необходимого условия активации энергетических и пластических процессов. Выраженность гормональных перестроек определяется продолжительностью и интенсивностью выполняемых мышечных нагрузок.
4. Становление адаптивных механизмов в организме животных сопряжено с гипертрофией надпочечников и повышением их секреторной активности за счёт пучковой и сетчатой зоны. В динамике секреции 11-оксикортикостероидов регистрируются фазовые изменения. Повышение на начальных этапах формирования адаптивных механизмов и уменьшение выраженности кортикальной реакции по мере развития тренированности.
5. Адаптация к большим по объёму и интенсивности динамическим нагрузкам проявляется в росте мышечной ткани, который наиболее эффективно регулируется мужскими половыми гормонами - тестостероном, в условиях отсутствия клеток Лейдинга в женском организме идёт интенсификация работы сетчатой зоны коры надпочечников, повышая уровень аналогов мужских половых гормонов у самок крыс.
6. Выявленные морфофункциональные изменения в яичниках самок крыс - увеличение количества атрезирующих фолликулов, следует рассматривать как одну из форм адаптивных реакций, которая наряду с гипертрофией надпочечников направлена на удовлетворение возросшей потребности женского организма в анаболических стероидах.
ЗАКЛЮЧЕНИЕ
Результаты изменений, проходящих в женском организме, полученные на самках крыс, позволили выстроить концепцию влияния интенсивных мышечных нагрузок на их репродуктивную систему.
Результаты морфометрической и гистологической оценок яичников, матки, маточных труб самок крыс, диагностика их гормонального статуса позволили установить ряд изменений в органах репродуктивной системы, которые обусловлены интенсивными мышечными нагрузками. В частности, установлено отставание в темпах увеличения массы и линейных размеров исследуемых органов у самок крыс экспериментальной группы. Общая тенденция изменений в половом аппарате самок крыс носит характер гипоплазии и гипотрофии.
Факторами, провоцирующими подобные изменения, являются гиперандрогения и повышение концентрации кортикостероидов, обусловленное гипертрофией сетчатой и клубочковой зон и высоким процентом атрезирующих фолликулов в яичниках. Угнетающее действие гормонов на темпы развития органов репродуктивной системы осуществляется как через центральные, так и периферические звенья гипофизарно-гонадной оси.
При этом необходимо учитывать, что сформировавшиеся как структурные, так и гормональные перестройки могут сохраняться и после прекращения тренировок, а развивающиеся вследствие этого дисфункции репродуктивной системы оставаться и даже прогрессировать.
Экстраполяция полученных экспериментальных данных на организм спортсменок позволяет выявить особенности адаптивных изменений в системах организма, в том числе и в органах репродуктивной системы, и будет содействовать разработке практических рекомендаций в физиологии женского спорта.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ
Работы, опубликованные в изданиях, рекомендованных ВАК при Министерстве образования и науки РФ
1. Самойленко, В.Ю. Влияние интенсивных мышечных нагрузок на репродуктивную систему женщин / Беляев Н.Г., Писков С.И., Самойленко В.Ю. // Вестник Российского университета дружбы народов. Серия «Медицина». – 2007. – №6. – С. 483-487.( 0,6 п/л, личный вклад 25%)
2. Самойленко, В.Ю. Возможные механизмы спортивной гиперандрогении / Беляев Н.Г., Самойленко В.Ю., Суслов К.Ю. // Вестник Ставропольского государственного университета. – 2011. –№74(3). – С. 75-80. ( 0,74 п/л, личный вклад 60%)
Работы, опубликованные в других изданиях
3. Самойленко, В.Ю. Влияние продолжительной мышечной нагрузки в условиях принимаемого фитопрепарата на функциональное состояние половых желёз. // Проблемы развития биологии и экологии на Северном Кавказе: материалы научной конференции «Университетская наука - региону». – Ставрополь: СКГТУ, 2004. – С.158 – 159 (0.12 п/л, личный вклад – 100%)
4. Самойленко, В.Ю. Адаптация организма к мышечным нагрузкам. // Проблемы развития биологии и экологии на Северном Кавказе: материалы научной конференции «Университетская наука - региону». – Ставрополь: Изд-во СГУ, 2005. – С. 245 – 247 (0,19 п/л, личный вклад – 100%)
5. Самойленко, В.Ю. Влияние мышечных нагрузок на морфофункциональное состояние репродуктивной системы самок / Беляев Н.Г., Самойленко В.Ю. // Стратегия развития Российской системы образования в сфере физической культуры (состояние, тенденции, перспективы). – Ставрополь, 2006. – С. 169–170 (0.12 п/л, личный вклад – 90%)
6. Самойленко, В.Ю. Некоторые гормональные особенности девушек, занимающихся вольной борьбой / Беляев Н.Г., Писков С.И., Резникова Е.В., Самойленко В.Ю. // Инновационные преобразования в сфере физической культуры, спорта и туризма. – Ростов-на-Дону, 2007. – С.278–280. (0.19 п/л, личный вклад – 40%)
7. Самойленко, В.Ю. Гормональные механизмы влияния тренировочных нагрузок на репродуктивную систему спортсменок / Беляев Н.Г., Писков С.И., Самойленко В.Ю. // Материалы международной научно-практической конференции "Научное обоснование физического воспитания, спортивной тренировки и подготовка кадров по физической культуре и спорту." - Минск: Изд.- во БГУФК, 2009. – Т.4. – С.304–307. (0.25 п/л, личный вклад – 50%)
8. Самойленко, В.Ю. Изучение последствий мышечных нагрузок для репродуктивной системы самок крыс / Беляев Н.Г., Самойленко В.Ю., Суслов К.Ю. // Фундаметальные исследования в биологии и медицине - Ставрополь: Изд.- во "Литера", 2009. – Вып.7. – С.3 – 5. (0.19 п/л, личный вклад – 60%)
9. Самойленко, В.Ю. Морфофункциональные особенности репродуктивной системы самок крыс адаптированных к мышечным нагрузкам / Беляев Н.Г., Самойленко В.Ю., Суслов К.Ю. // VIII Всероссийская конференция «Нейроэндокринология – 2010» – Санкт-Петербург, 2010. – С.32 – 33 (0.12 п/л, личный вклад – 60%)