

Характеристика провизорного морфогенеза промежуточной мезодермы млекопитающих на примере развития первичной почки крысы
На правах рукописи
Молокова Светлана Александровна
характеристика провизорного морфогенеза промежуточной мезодермы млекопитающих на примере развития первичной почки крысы
03.03.04 – клеточная биология, цитология, гистология
Автореферат
диссертации на соискание ученой степени
кандидата биологических наук
Томск – 2010
Работа выполнена в Государственном образовательном учреждении высшего профессионального образования Ханты-Мансийского автономного округа – Югры «Ханты – Мансийская государственная медицинская академия»
Научный руководитель:
доктор медицинских наук,
профессор Янин Владимир Леонидович
Официальные оппоненты:
доктор биологических наук,
профессор Петрова Ирина Викторовна
кандидат биологических наук Островерхова Надежда Васильевна
Ведущая организация:
Государственное образовательное учреждение высшего профессионального образования «Новосибирский государственный медицинский университет Федерального агентства по здравоохранению и социальному развитию»
Защита состоится «___» ___________ 2010 г. в ___ часов на заседании диссертационного совета Д 208.096.03 при Сибирском государственном медицинском университете по адресу: 634050, г. Томск, ул. Московский тракт, д. 2.
С диссертацией можно ознакомиться в научно-медицинской библиотеке Сибирского государственного медицинского университета.
Автореферат диссертации разослан «____» ___________ 2010 г.

Ученый секретарь
диссертационного совета Герасимов А.В.
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность исследования. Изучение закономерностей морфогенезов у высших позвоночных представляет собой один из ключевых вопросов биологии, поскольку необходимо для понимания механизмов и форм биологического движения и построения конструкций тканевого и органного уровней. Превращения промежуточной мезодермы являются основой морфогенезов мочевой и половой систем. Особенностью мезодермальных формообразовательных процессов, в сравнении с преобразованиями производных экто- и энтодермы, является их сложность, так как в их основе лежит взаимодействие провизорных и дефинитивных гисто- и органогенезов как отражение филогенетических преобразований. Важным методологическим подходом изучения морфогенезов служит концепция феномена провизорности (Соловьев Г.С., 2002; Соловьев Г.С. и соавт., 2004; Богданов А.В., 2005; Смышляева Р.К., 2006; Шилин К.О., 2010).
Филогенез мочевой системы у позвоночных постулирован и излагается как последовательное формирование предпочки, первичной почки и вторичной почки по мере «движения» видов по эволюционной лестнице (Гертвиг О., 1912; Пэттен Б.М., 1959; Кнорре А.Г., 1967; Токин Б.П., 1970; Фалин Л.И., 1976; Станек И., 1977; Янченко Н.В. 2005; Пантелеев С.М., 2006; Sainio K. et al., 2003; Dressler G., 2006; Davidson A.J., 2008; Inomata T. et al., 2009).
Первичная почка является провизорным органом, выполняющим важные функции в развивающемся организме и моделирующим механизмы развития и построения структурно-функциональных единиц (нефронов) и в целом метанефроса на уровне дефинитивного морфологического субстрата.
У млекопитающих и человека первичная почка образуется в эмбриогенезе как обязательный элемент формирующейся мочевой системы и состоит из нефронов, образованных мальпигиевым тельцем и канальцем, дифференцированным на проксимальный и дистальный отделы, открывающимся в мезонефральный проток, при этом структура нефрона вольфова тела сравнима со структурой нефрона метанефроса (Молчанова В.В., 1975; Krause W.J., Cutts J.H., Leeson C.R., 1979; Tiedemann K., Egerer G., 1984; Matsumoto T. et al., 1994; Maluf N.S., 1994; Янин В.Л. и соавт., 2000; Aly K.H., 2007).
Вместе с тем, имеются данные, что у некоторых млекопитающих (грызуны) нефроны первичной почки имеют структурные особенности и не в полной мере моделируют строение нефрона окончательной почки. Выявлены противоречия, касающиеся взаимоотношений вольфова протока и мезонефронов, строения мезонефронов в краниальном и каудальном отделах, этапов жизненного цикла органа. Отсутствует детальное описание структуры мезонефрона и его морфометрическая характеристика. Не достаточно внимания уделено изучению морфогенеза первичной почки с позиций концепции провизорности (Zamboni L., Upadhyay S., 1982; Бочаров Ю.С., 1988, Smith C., MacKay S., 1991; Соловьев Г.С., 2002; Sainio K. et al., 2003; Соловьев Г.С. и соавт., 2004; Смышляева Р.К., 2006).
Цель: выявить закономерности формирования и особенности структурно-функциональной организации провизорного органа – первичной почки в эмбриогенезе у крысы.
Задачи:
1. Выявить закономерности морфогенеза промежуточной мезодермы в ходе формирования первичной почки и особенности структурной организации мезонефронов в эмбриогенезе у крысы.
2. Определить морфометрические параметры мезонефронов первичной почки и их динамику в эмбриогенезе у крысы.
3. Определить информационные показатели первичной почки в эмбриогенезе у крысы.
4. Определить динамику содержания мочевины в амниотической жидкости в эмбриогенезе у крысы.
5. Уставить временные границы и этапы жизненного цикла первичной почки в эмбриогенезе у крысы.
Научная новизна исследования. Впервые проведено комплексное исследование провизорного морфогенеза промежуточной мезодермы в ходе развития первичной почки у крысы. Выявлены особенности строения мезонефрона первичной почки у крысы, проявляющиеся в структуре мезонефрального тельца и канальцев, взаимодействии мезонефронов с вольфовым протоком в краниальном и каудальном отделах первичной почки. Выявлена динамика морфометрических параметров мезонефрона, информационных показателей, содержания мочевины в амниотической жидкости в ходе формирования первичной почки. На основе полученных данных установлены длительность и этапы жизненного цикла первичной почки в эмбриогенезе у крысы.
Теоретическая и практическая значимость работы. В результате проведённых исследований существенно расширены представления о закономерностях превращений и морфогенетических возможностей промежуточной мезодермы, о принципах организации первичной почки в пренатальном развитии органов мочевой системы у млекопитающих. Полученные данные могут быть использованы при анализе результатов научных исследований по проблемам эмбрионального развития мочевой и половой систем у млекопитающих, а так же для понимания причин и механизмов возникновения врождённых аномалий органов мочевой и половой систем.
Реализация результатов исследования. Результаты исследования внедрены в учебный процесс кафедры биологии с курсом микробиологии Ханты-Мансийской государственной медицинской академии, кафедры гистологии, цитологии и эмбриологии Тюменской государственной медицинской академии, кафедры общей патологии Сургутского государственного университета в процессе преподавания разделов «Общая эмбриология», «Эмбриональное развитие мочевой и половой систем», «Систематика и характеристика типа Хордовых. Филогенез органных систем у хордовых. Онтофилогенетические предпосылки врождённых пороков развития систем органов у человека».
Положения, выносимые на защиту
1. Провизорный морфогенез промежуточной мезодермы в ходе развития первичной почки крысы характеризуется последовательными сальтаторными преобразованиями, имеющими кранио-каудальную направленность.
2. Мезонефроны первичной почки крысы характеризуются наличием примитивного мезонефрального тельца, проксимального и дистального канальцев. Мезонефроны краниального и каудального отделов органа различаются уровнем дифференцировки и способом взаимодействия с мезонефральным протоком.
3. Первичная почка крысы представляет собой провизорный экскреторный орган, жизненный цикл которого подразделяется на этапы.
Апробация результатов исследования. Материалы диссертации доложены на Всероссийской научной конференции «Фундаментальные и прикладные проблемы гистологии. Гистогенез и регенерация тканей» (г. Санкт-Петербург, 2004); Международном эмбриологическом симпозиуме «Закономерности эмбрио-фетальных морфогенезов у человека и позвоночных животных» (г. Ханты-Мансийск, 2004); Международной научной конференции «Медико-биологические и экологические проблемы здоровья человека на Севере» (г. Сургут, 2004); Y Всероссийском съезде анатомов, гистологов и эмбриологов (г. Казань, 2004); IV конгрессе Международной ассоциации морфологов (г. Казань, 2004); YII конгрессе Международной ассоциации морфологов (г. Орел, 2006); II Международном эмбриологическом симпозиуме «Югра-Эмбрио-2006: Закономерности морфогенезов в онтогенезе и эксперименте у человека и позвоночных животных» (г. Ханты-Мансийск, 2006); IX конгрессе Международной ассоциации морфологов (Республика Узбекистан, г.Бухара, 2008); Научной сессия, посвященной 10-летию Южно-Уральского научного центра РАМН «Медицинская академическая наука — здоровью населения Урала» (г. Челябинск, 2008); Международной гистологической конференции «Морфогенезы в эволюции, индивидуальном развитии и эксперименте», посв. 80-летию ЗДН РФ, проф. Дунаева П.В. (г.Тюмень, 2008); Всероссийской научной конференции «Нейробиологические аспекты морфогенеза и регенерации» (г.Оренбург, 2008); YII Всероссийской научной конференции «Бабухинские чтения» (г. Орел, 2009); X Конгресс Международной ассоциации морфологов (г. Ярославль, 2010).
Публикации. По теме диссертации опубликовано 22 научных работы, из них 9 – в научных журналах, рекомендованных ВАК России.
Объем и структура диссертации. Диссертация изложена на 173 страницах машинописного текста, содержит 20 таблиц, иллюстрирована 113 рисунками, в том числе 6 электроннограмами. Диссертационная работа состоит из введения, обзора литературы, материалов и методов исследования, результатов собственных исследований, обсуждения, выводов и библиографического списка, включающего 301 источников, из них 191 на русском и 110 – на иностранных языках.
Работа выполнена в рамках научно-исследовательской работы кафедры гистологии, эмбриологии и цитологии Ханты-Мансийской государственной медицинской академии «Закономерности морфогенеза у человека и позвоночных животных в онтогенезе, эксперименте и в условиях действия факторов среды», регистрационный номер 0120.081550.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Исследование проведено на 428 эмбрионах белых беспородных крыс от 11 до 15 суток внутриутробного развития, полученных от самок с датированным сроком беременности средней массой 200-250 грамм. Все манипуляции с животными выполнялись согласно приказу МЗ СССР «О гуманном обращении с экспериментальными животными» № 755 от 12 августа 1977 г. и в соответствии с «Правилами проведения работ с использованием экспериментальных животных» утверждёнными приказом Министерства высшего и среднего профессионального образования СССР № 742 от 13 ноября 1984 г.
Совокупность эмбрионов разделена на 17 возрастных групп с интервалом 6 часов в период развития от 11 суток 0 часов по 15 суток 0 часов суток post coitus (рс), что соответствует 13 – 17 стадиям развития крысы (Дыбан А.П., 1975). На каждом сроке развития изучены зародыши, полученные от 3-4 беременных самок (табл. 1).
У беременных самок после умерщвления зародыши извлекались из матки с неповрежденной амниотической оболочкой. У каждого зародыша амниотическая оболочка вскрывалась, производился забор амниотической жидкости, в которой определяли концентрацию мочевины колориметрическим методом с использованием набора реагентов для ферментативного определения мочевины в сыворотке крови производства ЗАО «Вектор-Бест». На автоматическом биохимическом анализаторе «Бэкман» измеряли оптическую плотность опытных (Е) и калибровочной (Ек) проб против контрольной пробы. Концентрацию мочевины рассчитывали по формуле С = Е/Ек х 5,00.
Таблица 1
Распределение эмбрионов крыс по стадиям развития и возрасту
| Стадия развития | Возраст эмбриона, сутки/часы | Количество самок | Количество эмбрионов |
| 13 | 11/0 | 3 | 17 |
| 11/6 | 3 | 26 | |
| 14 | 11/12 | 3 | 21 |
| 11/18 | 3 | 25 | |
| 12/0 | 4 | 33 | |
| 12/6 | 3 | 20 | |
| 15 | 12/12 | 3 | 26 |
| 12/18 | 3 | 20 | |
| 16 | 13/0 | 4 | 45 |
| 13/6 | 3 | 39 | |
| 13/12 | 3 | 22 | |
| 13/18 | 3 | 29 | |
| 17 | 14/0 | 4 | 33 |
| 14/6 | 3 | 15 | |
| 14/12 | 3 | 18 | |
| 14/18 | 3 | 18 | |
| 15/0 | 3 | 21 | |
| Всего: | 54 | 428 | |
Подготовку и гистологическую обработку биологического материала выполняли в соответствии с общепринятыми методами (Меркулов Г.А., 1961; Автандилов Г.Г., 1990; Саркисов Д.С. и соавт., 1996; Семченко В.В. и соавт., 2006).
Зародышей для светооптической микроскопии фиксировали в 10% нейтральном формалине, подвергали обезвоживанию, уплотнению, заливали в HISTOMIX. Срезы толщиной 1,5-4 микрометра изготавливали с помощью роторного микротома Microm HM 340Е (MICROM, Германия), окрашивали гематоксилином Караци и эозином, проводили ШИК-реакцию по Мак-Манусу. Гистологические препараты исследованы с использованием микроскопа Axio Imager Z1 (Zeiss, Германия).
Материал для электронно-микроскопического исследования фиксировался при 40С в параформальдегид-глутаральдегидной смеси с дофиксацией четырёхокисью осмия. Фиксаторы готовили на фосфатном буфере, рН растворов 7,2. Время от забора материала до фиксации не более 10 минут. Материал после фиксации обезвоживался и заливался в аралдит. Полутонкие срезы толщиной 1 мкм изготавливали стеклянными ножами на ультратоме «NOVA» («LKB», Швеция), окрашивали метиленовым синим. Ультратонкие срезы изготавливали алмазными ножами, контрастировали уранилацетатом и цитратом свинца (Уикли Б., 1975; Саркисов Д.С. и соавт., 1996). Исследование проведено на трансмиссионном электронном микроскопе JEM-1010 («JEOL», Япония).
Морфометрическое исследование выполнено с использованием программы AxioVision 4.6.3. Измерялись: площади эпителия, полостей в эпителиальных структурах, сосудов, мезенхимы в срезе мезонефроса (объектив: 40; окуляр: 10); минимального диаметра канальца и просвета канальца в разных отделах мезонефрона; максимального и минимального диаметров ядра клетки проксимального и дистального канальцев, висцерального и париетального листков капсулы мальпигиевого тельца мезонефрона. Площади канальцев и просветов канальцев вычислялись по формуле круга S=А2/4, где А – величина малого диаметра канальца в мкм. Площадь эпителия стенки канальцев определялась как разница площади канальца и площади его просвета. В каждом мезонефросе эмбрионов проводились измерения 10 – 20 канальцев. Всего в каждой возрастной группе измерялись 60 – 165 канальцев. Объёмы клеток и их ядер вычислялись по формуле V=/6.L.В2, где L – большой диаметр; В – короткий диаметр ядра (Автандилов Г.Г., 1990). Определись коэффициент элонгации ядра клетки по формуле Е=L/В; ядерно-цитоплазматические отношения для клеток канальцев и листков капсулы мезонефрона как отношение объёма ядра клетки к объёму цитоплазмы.
Индекс митоза (IМ) вычисляли как отношение числа митозов к общему количеству клеток и выражали в промиллях (‰) (Алов И.А., 1973). Индекс апоптоза (IА) вычисляли как отношение числа клеток с признаками апоптоза к общему количеству клеток и выражали в промиллях (‰). Индекс секреторной активности (IСЕКР) эпителиоцитов определяли как отношение числа клеток с признаками апокриновой секреции к общему количеству клеток и выражали в промиллях (‰).
Характеристика сложности и организации первичной почки определяли вычислением информационных показателей: информационной энтропии по формуле Шеннона: H = - Pi log2 Pi, где Р – объёмные доли тканевых компонентов первичной почки; максимальной энтропии как степени неопределённости информации по формуле Нmax= — m(Рi Iog2 Pi ), где под Pi подразумевается величина, обратная числу элементов системы m Pi=1/m; коэффициента относительной организации системы (или коэффициента избыточности) по формуле: R = (Hmax – H)/Hmax100% (Автандилов Г.Г., 1990).
Статистическую обработку результатов проводили с использованием программы Excel и пакета «Statistica for Windows ver. 6.0». Для каждого параметра определяли средние значения и ошибку среднего. Для оценки достаточности объектов исследования и количества измерений на изучаемых сроках эмбриогенеза первичной почки крысы использовался показатель точности опыта (Р), определяемый по формуле Р=(m:M)х100%, где M – среднее арифметическое площади, m – ошибка среднего. Во всех случаях данный показатель был меньше 5%, что указывает на достаточность проведенных исследований. После проверки распределения на нормальность, достоверность различий средних величин определяли по t-критерию Стьюдента или по U-критерию Манна-Уитни (Гельман В.Я., 2001). Для выявления различий между значениями индексов (IM, IA, IСЕКР) применяли критерий согласия 2. Оценку зависимости морфометрических параметров мезонефронов от возраста эмбриона проводили с использованием однофакторного дисперсионного анализа ANOVA.
Выражаю благодарность сотрудникам кафедры медицинской и биологической химии ХМГМА и лично д.м.н. Соловьёву В.Г. и Никулиной Е.Г. за оказанную консультативно-методическую помощь при проведении биохимических исследований.
Выражаю признательность за оказанную возможность проведения ультраструктурных исследований и методическую помощь сотрудникам лаборатории ультраструктурных исследований НИИ клинической и экспериментальной лимфологии Сибирского отделения РАМН и лично д.б.н., профессору Бгатовой Н.П., сотрудникам патологоанатомического отделения Окружной клинической больницы г. Ханты – Мансийска и лично к.м.н. Хадиевой Е.Д., к.м.н. Линингу Д.А.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
Структурная характеристика первичной почки крысы
На 13 стадии развития (11 суток 0 часов рс) у зародышей крысы на уровне 6-9 сомитов в боковых отделах тела наблюдается картина дифференцировки промежуточной мезодермы: формируются примитивные нефроны пронефроса, открывающиеся в целом. К окончанию 13 стадии (11 суток 6 часов рс) у эмбрионов каудальнее области головной почки в зоне промежуточной мезодермы обнаруживаются канальцы первичной почки и проток первичной почки. Канальцы, расположенные в краниальном отделе мезонефроса, имеют изогнутую, S-образную форму. Стенка канальцев образована однослойным цилиндрическим эпителием. Каналец слепо начинается в мезенхиме и противоположным концом открывается в вольфов проток. В каудальном отделе первичной почки наблюдаются шаровидные сгущения мезенхимных клеток.
На 14 стадии (11 суток 12 часов – 12 суток 6 часов рс) в краниальной части вольфова тела располагаются канальцы, слепо начинающиеся в мезенхиме и открывающиеся в мезонефральный проток. Канальцы характеризуются более высоким уровнем дифференцировки в сравнении со структурами, лежащими каудальнее. В составе краниального канальца, который идентифицирован нами как мезонефрон, формируются отделы, характеризующиеся особенностями структурно-функциональной организации: начальный отдел и собственно каналец, дифференцированный по протяженности на сегменты.
Начальный отдел мезонефрона слепо начинается в мезенхиме, расширен, имеет «грибовидную» форму и идентифицируется нами как примитивное мезонефральное тельце. Одна из стенок тельца куполообразно выгнута, образована одним слоем кубических и цилиндрических клеток и представляет собой париетальный листок капсулы тельца. Особенность данного листка - большое количество апоптозных телец.
Боковые стенки начального отдела образованы одним слоем цилиндрических клеток, расположенных на базальной мембране, с признаками апокриновой секреции. Одна из боковых стенок вдается в просвет начального отдела, при этом в клетках наблюдается перемещение ядер из базальной части в апикальную, базальные участки эпителиоцитов суживаются. Данные клетки представляют собой эпителиоциты, находящиеся на начальных этапах подоцитарной дифференцировки, и составляют висцеральный листок капсулы тельца. Со стороны мезенхимы к эпителиоцитами висцерального листка прилежит капилляр, соответствующий сосудистому компоненту почечного тельца. Между листками мезонефрального тельца располагается мочевое пространство.
Непосредственно с мезонефральным тельцем контактирует проксимальный каналец мезонефрона, имеющий наибольшие размеры в сравнении с последующим сегментом канальца. Стенка канальца образована одним слоем цилиндрических клеток с признаками апикально-базальной полярности и апокриновой секреции. Эпителиоциты располагаются плотно друг к другу, латеральные плазмолеммы имеют ровный рельеф, обнаруживаются специализированные межклеточные контакты по типу десмосом. В цитоплазме присутствуют свободные рибосомы, полисомы, цистерны гранулярной эндоплазматической сети. Митохондрии равномерно распределены в цитоплазме. В апикальном полюсе клетки присутствует комплекс Гольджи. Ядра эпителиоцитов овальной формы. В ядре преобладает эухроматин, часто присутствует 1-2 ядрышка. Базальная мембрана эпителия сформирована, имеет трехслойную структуру. Микроворсинки, характерные для эпителиоцитов проксимальных мезонефральных канальцев, описанные у других видов млекопитающих и человека, отсутствуют. Проксимальный каналец продолжается в дистальный каналец меньшего диаметра, выстланный низкопризматическими эпителиоцитами и открывающийся в вольфов проток.
Мезонефроны каудальной части органа, сохраняют структурные признаки мезонефронов, описанных выше. Однако дистальный каналец слепо заканчивается в мезенхиме и не открывается в вольфов проток. В наиболее каудальных отделах почки лежат сгущения клеток нефрогенной мезенхимы в виде тяжей, прилежащих к вольфову протоку.
К 15 стадии развития (12 суток 12 часов рс) мезонефроны краниальной части мезонефроса имеют типичную структуру и взаимодействуют с вольфовым протоком. Часть мезонефронов открывается в вольфов проток посредством общего канальца (рис. 1). Обращает на себя внимание развитая сосудистая система мезонефроса, представленная капиллярами, оплетающими мезонефроны и формирующими участки тесного контакта с базальной мембраной висцерального листка тельца.
 |  |
| Рис. 1. Зародыш крысы 12 суток 12 часов pc. Мезонефроны краниальной части первичной почки: 1 – начальный отдел мезонефрона, 2 – связующий каналец, 3 - вольфов проток. Окраска: гематоксилин и эозин. Увеличение: объектив: 40, окуляр: 10. | Рис. 2. Зародыш крысы 12 суток 18 часов pc. Мезонефральное тельце: 1 – париетальный листок; 2 – висцеральный листок; 3 – сосудистый компонент. Окраска: гематоксилин. Увеличение: объектив 40, окуляр 10. |
К окончанию 15 стадии средняя часть органа является наиболее развитой, о чём свидетельствуют крупные размеры мезонефронов, извитость канальцев, обильная васкуляризация. В некоторых мезонефронах тельце приобретает очертания, наиболее совпадающие с очертаниями мезонефральных телец, описанных в первичной почке человека и млекопитающих других видов (рис. 2).
В составе тельца выделяется подобие гломерулы, в структуре которой присутствуют цилиндрические клетки висцерального листка и сосудистый компонент. Клетки висцерального листка проявляют признаки апокриновой секреции. Эпителий париетального листка находится в состоянии деструкции: определяется большое количество апоптозных телец, клетки не формируют пласта, располагаются неупорядоченно, имеется непосредственный контакт просвета мезонефрона и мезенхимы.
Сравнение индексов митоза, апоптоза и секреторной активности позволяет выделить различия данных показателей в разных отделах мезонефрона (табл. 2).
Таблица 2
Значения IA, IM, IСЕКР для отделов мезонефрона крысы
| Индекс | Каналец | Листок капсулы | ||
| проксимальный | дистальный | висцеральный | париетальный | |
| IM | 60,93 | 44,04 | 55,73 | 3,09 |
| IA | 10,88 | 12,87 | 40,59 | 459,14 |
| Iсекр | 273,96 | 18,54 | 412,60 | - |
Фигуры митоза достоверно чаще определяются в эпителии проксимального канальца и висцерального листка капсулы тельца. Индекса митоза проксимального канальца 60,93‰, висцерального листка капсулы 55,73‰, дистального канальца 44,04‰, париетального листка капсулы 3,09‰. Значение IМ париетального листка капсулы достоверно (р0,05, по критерию 2) ниже IМ висцерального листка, проксимального и дистального канальцев.
Апоптоз наиболее выражен в эпителии париетального листка капсулы тельца: значение IА для него в среднем составляет 459,14 ‰. IА висцерального листка 40,59‰, IА эпителия проксимального канальца 12,87‰, IА эпителия дистального канальца 10,88‰. IА эпителия париетального листка капсулы мезонефрона достоверно (р0,05, по критерию 2) выше IА эпителия висцерального листка капсулы. Различия средних значений IА эпителия проксимального и дистального канальцев не достоверны.
Признаки апокриновой секреции проявляют эпителиоциты висцерального листка капсулы, проксимального и дистального канальцев. IСЕКР эпителия висцерального листка капсулы составил в среднем 431,12 ‰, проксимального канальца 278,79 ‰, дистального канальца 18,54‰. IСЕКР проксимального канальца достоверно (р0,05, по критерию 2) выше Iсекр дистальных канальцев. IСЕКР висцерального листка капсулы достоверно выше (р0,05, по критерию 2) IСЕКР проксимального канальца на 14 стадии развития (12 суток 6 часов рс) и на 16 стадии (13 суток 6 часов рс (р0,05, по критерию 2). На остальных сроках эмбриогенеза различия не достоверны.
На 16 стадии в мезонефросе наблюдается картина, свидетельствующая о начале регресса органа. В эпителии мезонефрона обнаруживаются многочисленные апоптозные тельца. Индексы апоптоза всех отделов мезонефрона достигают максимальных значений: IА проксимального канальца 50,00‰, дистального канальца 41,67‰, висцерального листка 231,88‰, париетального листка 695,12‰. Не смотря на то, что признаки апоптоза обнаруживаются в мезонефронах и краниального, и каудального отделов, апоптоз наиболее выражен в мезонефронах каудального отдела мезонефроса. На 17 стадии мезонефрос представлен канальцами, часть которых открывается в вольфов проток. Эпителиальные структуры утрачивают признаки мезонефронов и представляют собой канальцы, в последующем участвующие в морфогенезе половых желез.
Таким образом, структуры первичной почки у крысы определяются на сроках от 11 суток 6 часов рс до 14 суток 0 часов рс, однако период существования мезонефроса как провизорного органа мочеобразования следует ограничить 11 суток 6 часов рс – 13 суток 18 часов рс. Мезонефроны включают примитивное мезонефральное тельце, проксимальный и дистальный канальцы. В составе примитивного тельца выделяется висцеральный листок, с признаками апокриновой секреции и митотической активности, париетальный листок, подверженный апоптозу, мочевое пространство и сосудистый компонент, представленный кровеносным капилляром, не формирующим сосудистого клубочка. Проксимальный каналец, характеризуется более высокой, в сравнении с дистальным канальцем, митотической и секреторной активностью эпителия. Наиболее дифференцированные мезонефроны занимают в первичной почке краниальное положение и взаимосвязаны с мезонефральным протоком. В каудальной части вольфова тела располагаются мезонефроны, не имеющие структурной связи с мезонефральным протоком.
Морфометрическая характеристика канальцев мезонефронов
Морфометрический анализ увеличивает точность описания изучаемого морфогенеза. В результате морфометрического исследования установлено, что площади сечения, просвета и эпителия проксимального канальца достоверно больше площадей дистального канальца на всех изученных сроках эмбриогенеза (рис. 3).

Рис. 3. Площади проксимальных и дистальных канальцев мезонефрона крысы
Площади проксимальных канальцев в среднем составляют: поперечного сечения 1482,22 ± 13,97 мкм, просвета 53,88 ± 1,66 мкм, эпителия 1428,44 ± 13,02 мкм (табл. 3, рис. 4).
Таблица 3
Площади проксимальных канальцев
| Возраст (сутки/часы) | Площадь сечения | Площадь просвета | Площадь эпителия | |||
| X±Sx (мкм) | % прироста | X±Sx (мкм) | % прироста | X±Sx (мкм) | % прироста | |
| 11/12 | ||||||
| 11/18 | 1374,00±22,11 | - | 16,87±0,77 | - | 1357,13±21,75 | - |
| 12/0 | 1713,32±26,38* | 24,70 | 54,01±2,68* | 220,15 | 1659,31±25,62* | 22,27 |
| 12/6 | 1712,61±48,88 | -0,04 | 85,41±3,49* | 58,14 | 1627,20±42,74 | -1,94 |
| 12/12 | 1637,46±28,10 | -4,39 | 68,99±3,90* | -19,22 | 1568,46±26,21 | -3,61 |
| 12/18 | 1636,39±40,38 | -0,07 | 66,65±3,37 | -3,39 | 1569,75±38,14 | 0,08 |
| 13/0 | 1578,23±35,33 | -3,55 | 55,04±2,67* | -17,42 | 1523,19±32,91 | -2,97 |
| 13/6 | 1482,02±43,17* | -6,10 | 54,45±2,69 | -1,07 | 1428,44±39,34* | -6,22 |
| 13/12 | 1290,79±38,56* | -12,90 | 58,44±2,91 | 7,33 | 1232,35±36,27* | -13,73 |
| 13/18 | 915,21±32,83* | -29,10 | 25,07±1,30* | -57,10 | 890,14±32,66* | -27,77 |
| 11/18-13/18 | 1482,22±13,97 | - | 53,88±1,66 | - | 1428,44±13,02 | - |
Примечание: * - изменение статистически достоверно в сравнении с показателем предыдущего срока эмбриогенеза (* - р<0,05).
Доля просвета и эпителия в среднем составляют 3,63 % и 96,37 % соответственно. Площади дистальных канальцев в среднем составляют: поперечного среза 743,10 ± 7,17 мкм, просвета 15,29 ± 0,29 мкм, эпителия 727,83 ± 7,26 мкм (табл. 4, рис. 5). Доля просвета и эпителия в среднем составляют 2,06 % и 97,95 % соответственно. Площади сечения канальца, просвета и эпителия проксимального канальца статистически достоверно (р0,05) больше площадей дистального канальца у всех изученных эмбрионов.
Наибольших средних значений площади сечений и эпителия проксимальных канальцев достигают у эмбрионов возраста 12 суток 0 часов рс, площади просвета у эмбрионов 12 суток 6 часов рс (табл. 3, рис. 4).
Наибольшие средние значения площади сечения и эпителия дистальных канальцев приходятся на 12 суток 0 часов. Площадь просвета достигает наибольшего значения в 12 суток 18 часов (табл. 4, рис. 5).
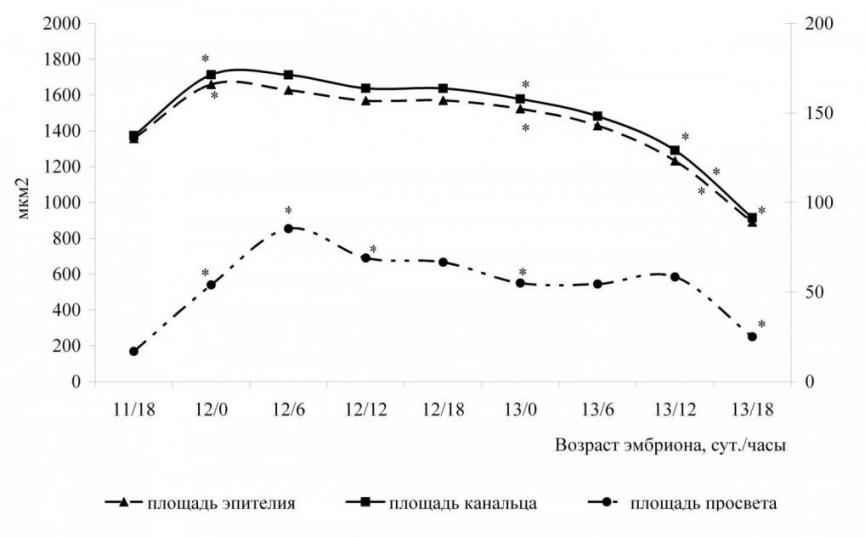 | 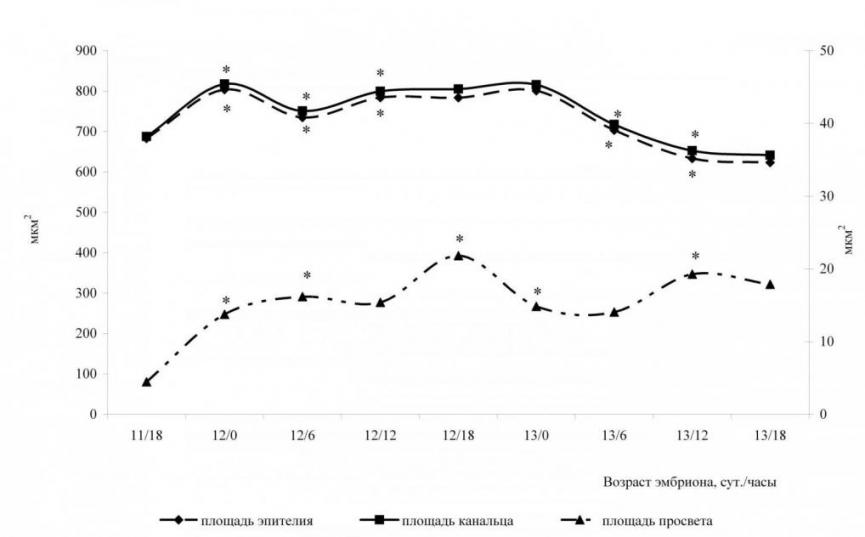 |
| Рис. 4. Изменение средних площадей проксимальных канальцев мезонефронов | Рис. 5. Изменение средних площадей дистальных канальцев мезонефронов |
| Примечание: * - изменение статистически достоверно в сравнении с показателем предыдущего срока эмбриогенеза (* - р<0,05). | |
Таблица 4
Площадь дистальных канальцев
| Возраст (сут./час.) | Площадь сечения | Площадь просвета | Площадь эпителия | |||
| X±Sx (мкм) | % прироста | X±Sx (мкм) | % прироста | X±Sx (мкм) | % прироста | |
| 11/18 | 687,23±21,18 | - | 4,47±0,19 | - | 682,96±24,57 | - |
| 12/0 | 817,64±14,53* | 18,94 | 13,75±0,67* | 207,61 | 803,88±14,19* | 27,81 |
| 12/6 | 750,93±10,92* | -8,16 | 16,17±0,80* | 17,60 | 734,76±10,42* | -8,60 |
| 12/12 | 799,55±24,44* | 6,47 | 15,39±0,65 | -4,82 | 784,16±23,93* | 6,72 |
| 12/18 | 805,21±25,54 | 0,71 | 21,80±1,03* | 41,65 | 783,66±24,49 | -0,06 |
| 13/0 | 815,80±19,57 | 1,32 | 14,85±0,73* | -34,02 | 800,95±23,94 | 2,21 |
| 13/6 | 717,54±22,25* | -12,04 | 14,06±0,62 | -2,25 | 703,48±21,83* | -12,17 |
| 13/12 | 652,56±20,50* | -9,06 | 19,27±0,96* | 37,06 | 633,30±20,49* | -9,98 |
| 13/18 | 641,67±21,67 | -1,74 | 17,87±0,81 | -7,27 | 623,35±20,96 | -1,57 |
| 11/18-13/18 | 743,10±7,17 | 15,29±0,29 | 727,83±7,26 | - | ||
Примечание: * - изменение статистически достоверно в сравнении с показателем предыдущего срока эмбриогенеза (* - р<0,05).
Площади сечений проксимальных и дистальных канальцев достоверно (р0,05) увеличиваются с возрастом эмбриона на 14 стадии развития (11 суток 18 часов рс – 12 сутки рс). Площади сечения канальцев не зависят (р=0,27) от возраста эмбриона в конце 14 стадии и на протяжении 15 стадии развития (12 суток 0 часов по 13 сутки 0 часов рс), т.е. данный этап жизненного цикла мезонефроса характеризуется стабильностью площадей сечения канальцев; на 16 стадии (13 суток 0 часов – 13 суток 18 часов рс) площади сечений канальцев достоверно (р0,05) уменьшаются с возрастом эмбриона.
Площади просветов проксимальных канальцев не зависят (р=0,140) от возраста эмбриона в конце 14 стадии и в течение 15 стадии развития (12 суток 6 часов – 12 суток 18 часов рс). Начиная с 16 стадии развития (13 суток 6 часов рс) можно говорить о следующем периоде в состоянии просветов проксимальных канальцев, когда площади просветов достоверно (р0,005) уменьшаются с возрастом эмбриона. Площади просветов дистального канальца достоверно (р0,05) зависят от возраста эмбриона на всём сроке существования мезонефроса. Период стабилизации площадей просветов дистального канальца не выявлен.
Площади эпителия проксимальных и дистальных канальцев достоверно (р0,05) увеличиваются с возрастом эмбриона на 14 стадии (с 11 суток 18 часов рс по 12 сутки рс). Площади эпителия канальцев не зависят (р=0,140) от возраста эмбриона с конца 14 и на 15 стации (с 12 суток 0 часов по 12 сутки 18 часов рс); на 16 стадии (с 13 суток 0 часов рс для проксимальных канальцев и с 13 суток 6 часов рс для дистальных канальцев) площади эпителия проксимальных канальцев достоверно (р0,05) уменьшаются с возрастом эмбриона.
Таким образом, сравнение морфометрических показателей выявляет наличие объективных признаков, свидетельствующих о дифференцировке нефрона на отделы. На 14-15 стадиях морфометрические параметры мезонефрона достигают наибольших значений, на 16 стадии наблюдается уменьшение морфометрических параметров.
Кариоцитометрическая характеристика отделов мезонефрона
Объём эпителиоцита проксимального канальца составляет в среднем 234,22±4,81 мкм3, дистального канальца – 210,32±3,69 мкм3. Объём ядра эпителиоцита проксимального канальца составляет в среднем 93,86±2,29 мкм3, объём ядра эпителиоцита дистального канальца 84,74±1,90 мкм3. Наибольшие объёмы клеток эпителия проксимального и дистального канальцев наблюдается у эмбрионов 12 суток 12 часов рс и составляет 323,77±20,24 мкм3 и 254,00±10,02 мкм3 соответственно. Объём клетки и ядра эпителиоцита проксимального канальца достоверно (р0,05) больше, чем объём клетки и ядра эпителиоцита дистального канапльца у эмбрионов на 15 стадия развития (12 суток 0 часов рс – 12 суток 18 часов рс).
Объём клеток висцерального листка тельца в среднем составляет 237,73±3,58 мкм3, клеток париетального листка 204,96±5,65 мкм3. Объём ядра эпителиоцита висцерального листка в среднем составляет 88,75±1,76 мкм3, объём ядра эпителиоцита париетального листка 84,87±2,67 мкм3. Наибольший объём клетки эпителия висцерального листка наблюдается у эмбрионов на 14 стадии (12 суток 6 часов рс) и составляет 275,01±9,49 мкм3, наибольший объём клетки эпителия париетального листка выявлен у эмбрионов на 16 стадии (13 суток 12 часов рс) и составляет 273,02±16,84 мкм3. Достоверное (р0,05) преобладание объёма эпителиоцитов висцерального листка над объёмом эпителиоцитов париетального листка выявлено у эмбрионов на 14-15 стадиях и в начале 16 стадии (11 суток 18 часов рс – 12 суток 12 часов рс и 13 суток 0 часов – 13 суток 6 часов рс).
Таким образом, данные кариоцитометрии позволяют выявить достоверное преобладание объёмов клеток и ядер проксимального канальца над аналогичными параметрами дистального канальца на 14 и 15 стадии развития эмбриона крысы. Объём клеток висцерального листка больше объёма клеток париетального листка мезонефрального тельца на 14, 15 и в начале 16 стадии, объём ядер эпителиоцитов висцерального листка больше объёма ядер эпителиоцитов проксимального листка на 14 стадии.
Информационная характеристика первичной почки
Мезонефрон является значимым, но не единственным компонентом первичной почки. На этапах жизненного цикла в органе меняется соотношение эпителиальных структур, полостей, сосудистого русла, мезенхимы. Для более полной характеристики направленности морфогенетических процессов необходимо рассмотрение органа как системы с позиций теории информации, выявление отношений между её компонентами, установление закономерностей пространственно-временной организации системы. Анализ изменения морфометрических характеристик с учётом теории информации позволил получить интегральные критерии состояния биологической системы на этапах онтогенеза.
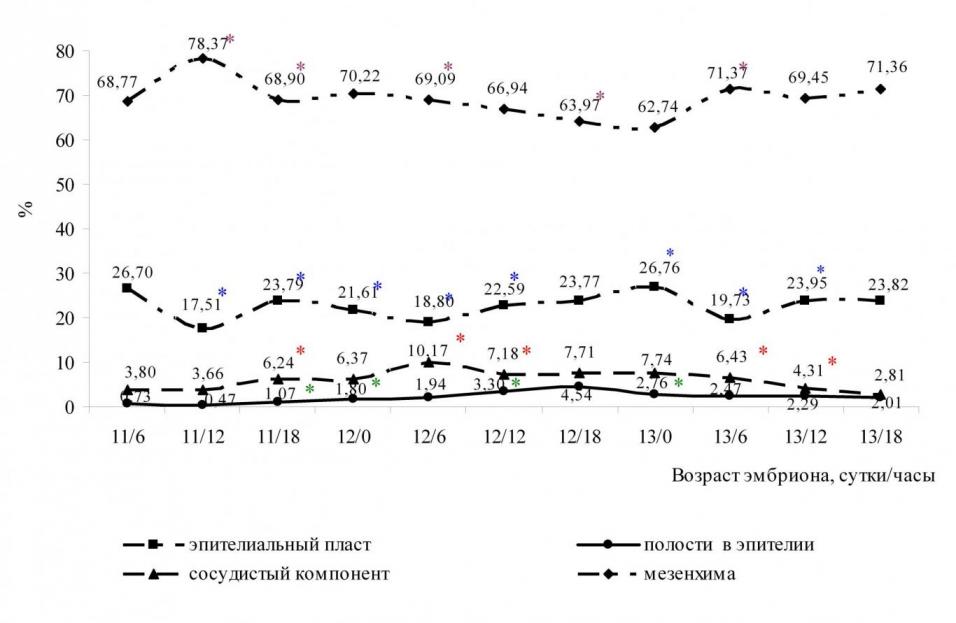
В первичной почке крысы статически достоверно (р0,05) преобладает мезенхимный компонент, не участвующий в образовании мочи. Доля мезенхимы первичной почки в среднем составляет 69,09% (рис. 6).
| А | 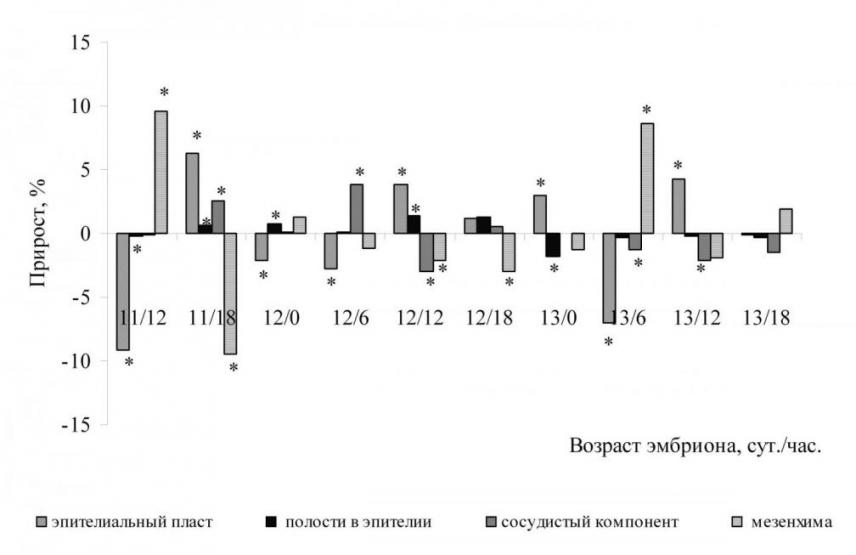 Б Б |
Рис. 6. Объёмные доли тканевых компонентов первичной почки крысы (А) и их прирост (Б)
Примечание: * - изменение статистически достоверно в сравнении с показателем предыдущего срока эмбриогенеза (* - р<0,05).
Доля эпителиального пласта в среднем за весь период наблюдений составляет 22,76%, минимальное значение отмечается у эмбрионов возраста 11 суток 12 часов рс (17,51%), максимальное значение – у эмбрионов возраста 13 суток 0 часов рс (26,76%). Доля полостей в эпителиальных структурах мезонефроса в среднем составляет 2,14%, максимальное значение приходится на 15 стадию и составляет 4,54 %. Доля сосудистого компонента мезонефроса в среднем составляет 6%, максимальное значение приходится на окончание 14 стадии и составляет 10,17%.
На 14 стадии развития (11 суток 18 часов) доля эпителия достоверно (р0,05) возрастает, что связано с развитием эпителиальных структур в органе. К окончанию 14 стадии происходит достоверное (р0,05) уменьшение доли эпителиального пласта за счёт увеличения объёмной долей полостей в эпителиальных структурах и доли сосудистого компонента мезонефроса. На 15 стадии развития объёмная доля эпителия возрастает, достигает максимального значения к началу 16 стадии (13 суток 0 часов рс). На 16 стадии наблюдается уменьшение долей эпителия, полостей и сосудов.
Биологические системы оценивают, используя понятия теории информации, ключевым из которых является информационная энтропия как мера неопределённости информации. Большинство многокомпонентных морфологических систем сочетают устойчивость с большими возможностями адаптации к изменениям среды, т.е., обладают относительной устойчивостью.
Информационная ёмкость системы рассчитана, исходя из количества компонентов системы, характеризуется показателем максимальной энтропии (Hmax). Первичная почка включает в себя четыре компонента: мезенхима, эпителий, полости в эпителиальных структурах, сосудистый компонент. Показатель Hmax составляет 2,00 бит.
Рассматривая морфогенез провизорной почки необходимо учитывать двоякий смысл происходящих изменений во взаимоотношениях компонентов системы. С одной стороны, изменение структуры провизорного органа – это проявление свойства открытости как роста возможностей выхода к новому структурно-функциональному состоянию – метанефросу. С другой стороны, изменение структуры первичной почки как системы обусловлено возрастающей функциональной нагрузкой, связанной с ростом эмбриона.
Информационная энтропия системы «первичная почка крысы» у эмбрионов на 13 стадии (11 суток 6 часов) составляет 1,06±0,02 бит. На 14 стадии развития крысы (11 суток 12 часов рс) происходит достоверное (р0,05) увеличение показателя энтропии (рис. 7). К окончанию 15 стадии его значение достигает максимума (1,36 бит) у эмбрионов возраста 12 суток 18 часов рс, что связано с изменением соотношения компонентов системы, а именно возрастанием долей эпителия, полостей в эпителиальных структурах и сосудов в связи с возрастающей функциональной нагрузкой. Повышение уровня энтропии также свидетельствует о снижении устойчивости системы. Информационная энтропия (Н) не достигает значения максимальной энтропии Hmax (2 бит) за весь период существования органа. Это свидетельствует о наличии избыточности системы. На 16 стадии энтропия достоверно (р0,05) снижается, что связано с возрастанием доли мезенхимы и преобладанием её над другими компонентами первичной почки.
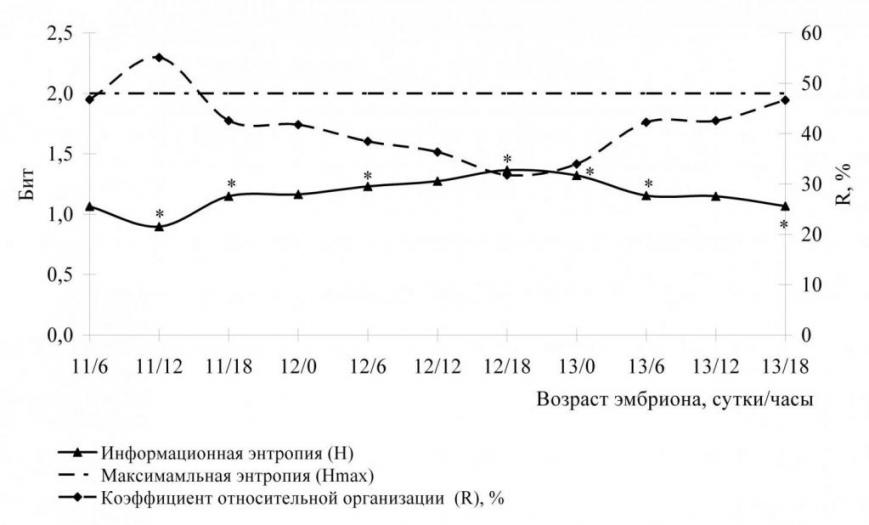
Рис. 7. Информационная энтропия, максимальная энтропия и коэффициент избыточности первичной почки
Примечание: * - изменение статистически достоверно в сравнении с показателем предыдущего срока эмбриогенеза (* - р<0,05).
Коэффициент относительной организации (избыточности) достигает минимального значения к окончанию 15 стадии (12 суток 18 часов рс) – 31,78%. На 16 стадии коэффициент избыточности возрастает, что связано с увеличением устойчивости системы, обусловленной преобладанием одного из компонентов первичной почки - мезенхимы (рис. 7).
Таким образом, исследование объёмных долей компонентов первичной почки показало, что в органе преобладает мезенхимный компонент, однако к окончанию 14 стадии увеличиваются объёмные доли эпителия, полостей в эпителии и сосудистого компонента. Максимального значения объёмная доля сосудистого компонента достигает к окончанию 14 стадии, эпителия в начале 16 стадии, полостей в эпителиальных структурах на 15 стадии. Энтропия первичной почки крысы увеличивается на 14 стадии, достигает максимального значения на 15 стадии и снижается на 16 стадии, что связано с увеличением доли мезенхимы и свидетельствует о нарастании избыточности в системе.
Анализ содержания мочевины в амниотической жидкости эмбрионов крысы
Известно, что основным метаболитом белкового обмена млекопитающих является мочевина. Мезонефральная моча у зародышей млекопитающих и птиц образуется путем клубочковой ультрафильтрации, канальцевой реабсорбции и секреции (Satlin L.M., 1994; Константинов В.М. и соавт., 2005), выводится через Вольфов проток в клоаку и аллантоис (Ros M.A., 1996), участвует в формировании аллантоисной и амниотической жидкостей (Wlodek M.E., 1988; Wu G., 1995; Wintour E.M., 1996).
Концентрация мочевины в амниотической жидкости эмбрионов крыс в среднем составила 6,90±0,14 ммоль/л. У эмбрионов возраста 11 суток 18 часов рс концентрация мочевины составила 4,90±0,21 ммоль/л. К 12 суткам 0 часам эмбриогенеза концентрация мочевины статистически достоверно (р0,05) увеличилась на 16,79% и составила 5,83±0,04 ммоль/л. У эмбрионов возраста 12 суток 6 часов рс концентрация мочевины достоверно (р0,05) увеличилась на 56,66% и составила 8,92±0,34 ммоль/л (рис. 8).
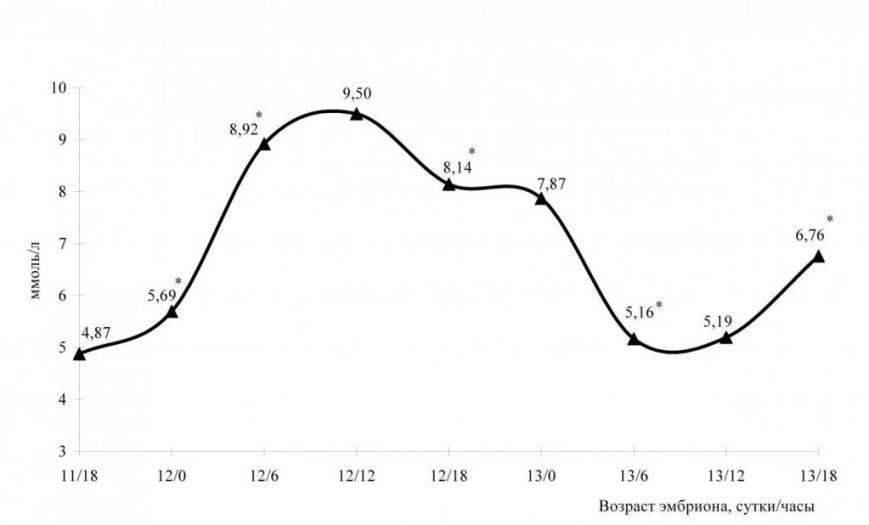
Рис. 8. Концентрация мочевины в амниотической жидкости эмбрионов крыс,
* - изменение статистически достоверно в сравнении с предыдущим сроком эмбриогенеза
Примечание: * - изменение статистически достоверно в сравнении с показателем предыдущего срока эмбриогенеза (* - р<0,05).
К 12 суткам 12 часам эмбриогенеза содержание мочевины в амниотической жидкости достигло максимума и составило 9,50±0,34 ммоль/л. Прирост составил 6,50%, изменение статистически не достоверно. У эмбрионов возраста 12 суток 18 часов рс концентрация мочевины достоверно (р0,05) уменьшилась на 14,27% (8,14±0,17 ммоль/л).
У эмбрионов возраста 13 суток 0 часов концентрация мочевины составила 7,91±0,36 ммоль/л. У эмбрионов возраста 13 суток 6 часов рс концентрация мочевины снизилась (р0,05) на 34,538% и составила 5,16±0,21 ммоль/л. У эмбрионов возраста 13 суткок 18 часов рс концентрация мочевины достоверно (р0,05) увеличилась на 30,13% и составила 6,78±0,23 ммоль/л.
Таким образом, на 14 стадии происходит достоверное увеличение концентрации мочевины в амниотической жидкости эмбрионов крыс, и к началу 15 стадии содержание мочевины достигает максимума. К окончанию 15 и на 16 стадии концентрация мочевины достоверно уменьшается.
Данные о структурной организации мезонефронов, динамике морфометрических параметров мезонефронов, информационных показателей первичной почки и концентрации мочевины в амниотической жидкости эмбрионов крыс позволяют определить границы и этапы жизненного цикла провизорного органа. Жизненный цикл первичной почки крысы соответствует 11 суткам 6 часам – 13 суткам 18 часам эмбриогенеза крысы и характеризуется наличием этапов: закладки и дифференцировки мезонефронов – 13 стадия; структурной стабильности – 14-15 стадии; регресса – 16 стадия и позднее.
ВЫВОДЫ
1. Провизорный морфогенез промежуточной мезодермы на этапах развития первичной почки у крысы характеризуется последовательными сальтаторными преобразованиями, имеющими кранио-каудальную направленность: «конденсация клеток мезенхимы» - «мезенхимно-эпителиальная дифференцировка клеток и формирование шаровидных клеточных сгущений» - «формирование эпителиальных замкнутых канальцев» - «дифференцировка эпителиальных канальцев в мезонефрон».
2. Мезонефроны первичной почки крысы обладают признаками структурной специфичности, что проявляется в наличии примитивного мезонефрального тельца, проксимального и дистального канальцев, характеризующихся разным уровнем митотической активности, апоптоза и апокриновой секреции. Мезонефроны краниальной части мезонефроса взаимосвязаны с вольфовым протоком, мезонефроны каудальной части не имеют связи с вольфовым протоком.
3. Морфометрические параметры мезонефронов возрастают в течение 14 стадии развития крысы, достигают максимальных значений на 15 стадии и уменьшаются в течение 16 стадии. Энтропия первичной почки крысы увеличивается на 14 стадии, достигает максимального значения на 15 стадии и снижается на 16 стадии. Концентрация мочевины в амниотической жидкости увеличивается на 14 стадии развития, достигает максимального значения к 15 стадии, снижается на 16 стадии.
4. Жизненный цикл первичной почки крысы соответствует 13 – 17 стадиям развития и включает этапы: закладки и дифференцировки мезонефронов – 13 стадия; структурной стабильности – 14-15 стадии; регресса – 16 стадия и позднее.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ
1. Дунаев, П.В. Динамика биологических потенций тканей в онтогенезе / П.В. Дунаев, Г.С. Соловьёв, В.А. Агарков, В.Л. Янин, О.Ф. Истомина, С.М. Пантелеев, С.А. Молокова и др. // Научный вестник Тюменской медицинской академии. – 2000. – № 3. – С. 86.
2. Соловьёв, Г.С. Проявления органотипический детерминированности провизорных и дефинитивных тканей / Г.С. Соловьёв, В.Л.Янин, В.А. Агарков, О.Ф. Истомина, С.А. Молокова и др. // Материалы I Всероссийской конференции «Актуальные проблемы эволюционной и популяционной физиологии человека» (г. Сургут). – 2001. – С. 85-86.
3. Соловьёв, Г.С. Эмбриологические исследования ранних этапов развития человека как основа оценки уровня здоровья населения / Г.С. Соловьёв, В.Л.Янин, С.М. Пантелеев, С.И. Матаев, В.А. Агарков, О.Ф. Истомина, С.А. Молокова и др. // Научный вестник Тюменской медицинской академии. – 2001. – № 3. – С. 68-69.
4. Соловьёв, Г.С. Характеристика пренатальных морфогенезов как основа эмбрио-фетальной медицины / Г.С. Соловьёв, В.Л. Янин, В.А. Агарков, О.Ф. Истомина, С.А. Молокова и др. // Научный вестник Тюменской медицинской академии. – 2001. - № 4 – С. 75.
5. Янин, В.Л. Особенности организации органных систем на ранних этапах пренатального онтогенеза у человека / В.Л. Янин, Г.С. Соловьёв, С.А. Молокова // Научный вестник Тюменской медицинской академии. – 2002. - № 7-8 – С. 146.
6. Соловьёв, Г.С. Показатели эмбриогенеза как критерии состояния и прогноза здоровья населения / В.Л. Янин, В.А. Агарков, О.Ф.Истомина, С.А. Молокова и др. // Сборник материалов международной научной конференции «Медико-биологические и экологические проблемы здоровья человека на Севере» (г. Сургут). – 2002. – С. 5-6.
7. Янин, В.Л. Эмбрио-фетальные стандарты как основа концепции оценки и прогнозирования уровня популяционного здоровья на территории Ханты-Мансийского автономного округа / В.Л. Янин, Г.С.Соловьёв, А.Н. Дурыгин, С.М. Пантелеев, А.В. Соловьёва, С.А. Молокова и др. // Медицина и охрана здоровья. – 2002. - № 7-8 – С. 146.
8. Янин В.Л. Мезометанефральные параллели в пренатальном онтогенезе у человека / В.Л. Янин, Г.С. Соловьев, Л.В. Вихарева, Р.К. Смышляева, С.А. Молокова и др. // Морфология.- 2004.- Т. 126, № 4.- С. 149.
9. Соловьев Г.С. Принцип провизорности как универсальный механизм морфогенезов у человека и млекопитающих / Г.С. Соловьев, В.Л. Янин, А.В. Богданов, А.В. Контарев, С.А. Молокова и др. // Материалы научной конференции «Фундаментальные и прикладные проблемы гистологии. Гистогенез и регенерация тканей» (г. Санкт-Петербург). – 2004. – С. 14-16.
10. Соловьев Г.С. Принцип провизорности как универсальный механизм эволюционирования гисто- и органогенезов / Г.С. Соловьев, В.Л. Янин, С.М. Пантелеев, Л.В. Вихарева, О.Ф. Истомина, А.В. Богданов, А.В. Контарев, О.В. Струихина, Р.К. Смышляева, С.А. Молокова и др. // Фундаментальные исследования (научно-теоретический журнал). – 2005.- № 9.- С. 32-34.
11. Соловьев Г.С. Роль принципа провизорности в реализации филэмбриогенезов / Г.С. Соловьев, В.Л. Янин, В.Д. Новиков, С.М. Пантелеев, Л.В. Вихарева, А.В. Контарев, С.А. Молокова // Морфология. - 2005.- Т. 128, № 4. С. 14-19.
12. Смышляева Р.К. Развитие и преобразование мезонефронов первичной почки птицы на этапах инкубации выводковой камеры / Р.К. Смышляева, Н.А. Агафонова, Н.П. Носова, С.А. Молокова // Медицинская наука и образование Урала. – 2007. - № 6. – С. 18-20.
13. Соловьёв Г.С. Морфогенез мезонефрона / Г.С. Соловьёв, В.Л. Янин, Р.К. Смышляева, Н.П. Носова, В.Г. Соловьёв, С.А. Молокова // Морфология. - № 2. – С. 126.
14. Смышляева Р.К. Механизмы морфогенеза почечных телец мезонефроса куриного эмбриона / Р.К. Смышляева, А.В. Контарева, В.Г. Соловьев, С.А. Молокова и др.//Морфология. - Том 133. – № 3. – 2008. – С.102.
15. Соловьёв Г.С. Реализация принципа провизорности в имплантантах / В.Л. Янин, С.М. Пантелеев, Л.В. Вихарева, О.Ф.Истомина, С.А. Молокова и др. //Морфология. – 2008. – Том 134. – № 5. – С. 93.
16. Янин В.Л. Нефроногенез первичной почки человека и птицы / В.Л. Янин, Г.С. Соловьёв, Р.К. Смышляева, А.В. Богданов, Н.А. Агафонова, О.Ф. Истомина, Н.П. Носова, С.А. Молокова и др. // Морфология. – 2008. – Том 134. – № 5. – С. 106-107.
17. Янин В.Л. Системогенез органов мочеобразования / В.Л. Янин, Г.С. Соловьев, Л.В. Вихарева, С.А. Молокова и др. // Морфология. – 2009. – т. 136. – №. 4. – С.162.
18. Янин В.Л. Эволюционная динамика и преемственность эмбриональных морфогенезов органов мочеобразования у высших позвоночных / В.Л. Янин, Г.С. Соловьев, Р.К. Смышляева, С.А. Молокова и др. // Ретиноиды. Альманах. Вып. 29. «Бабухинские чтения в Орле. 4-5 июня 2009 г.» Материалы Всероссийской научной конференции. Москва: изд-во ЗАО Ретиноиды, 2009. – С. 183-186.
19. Янин В.Л. Структурные и биохимические параллели морфогенеза мочевой системы и состава амниотической жидкости в пренатальном онтогенезе у белых крыс / В.Л. Янин, В.Г. Соловьев, Г.С. Соловьев, С.А. Молокова и др. // Сборник докладов IX Региональной конференции молодых ученых, специалистов и студентов. – Ханты-Мансийск: издательский центр ХМГМИ, 2009. – С. 111-114.
20. Баталов Е.Г. Структурно-функциональная характеристика мезонефральных телец первичной почки у эмбрионов белых крыс / Е.Г. Баталов, Г.Н. Севастьянова, А.А. Бабкина, Н.В. Иванова, С.А. Молокова // Научный вестник Ханты-Мансийской государственной медицинской академии. – 2010. - № 1-2. – С. 73-75.
21. Янин В.Л. Сравнительная характеристика мезонефроногенеза у млекопитающих и птиц / В.Л. Янин, В.И. Гарчук, Н.В. Иванова, О.Ф. Истомина, С.А. Молокова // Морфология. – 2010. – № 4. – С. 228.
22. Янин В.Л. Структурная характеристика морфогенеза мезонефроса у белых крыс / В.Л. Янин, Г.С. Соловьёв, С.А. Молокова, О.М. Бондаренко // Морфология. – 2010. – № 4. – С. 229.
СПИСОК ИСПОЛЬЗУЕМЫХ СОКРАЩЕНИЙ
рс - post coitus
IM – индекс митоза
IA – индекс апоптоза
IСЕКР – индекс секреторной активности