

Пептиды тимуса и их роль в регуляции стресса
На правах рукописи
КИСЕЛЕВА Нина Михайловна
ПЕПТИДЫ ТИМУСА И ИХ РОЛЬ В РЕГУЛЯЦИИ СТРЕССА
03.03.01 – физиология
14.03.06 – фармакология, клиническая фармакология
Автореферат
диссертации на соискание ученой степени
доктора биологических наук
Москва - 2013
Работа выполнена в Государственном бюджетном образовательном учреждении высшего профессионального образования «Российский национальный исследовательский медицинский университет имени Н.И. Пирогова» Министерства здравоохранения Российской Федерации, Московском государственном университете имени М.В. Ломоносова, Федеральном государственном бюджетном учреждении науки «Научно-исследовательский институт физико-химической медицины» Федерального медико-биологического агентства Российской Федерации
Научные консультанты:
| доктор биологических наук | Иноземцев Анатолий Николаевич |
| доктор медицинских наук, профессор | Козлов Иван Генрихович |
Официальные оппоненты:
| доктор медицинских наук, профессор кафедры физиологии медико-биологического факультета ГБОУ ВПО «Российский национальный исследовательский университет им. Н.И. Пирогова» Минздрава России. | Алипов Николай Николаевич |
| доктор биологических наук, профессор лаборатории механизмов природных гипометаболических состояний ФГБУН Институт Биофизики Клетки РАН | Семенова Татьяна Павловна |
| доктор медицинских наук, профессор, член-корреспондент РАМН заведующий кафедрой общей и клинической фармакологии медицинского факультета РУДН | Лепахин Владимир Константинович |
Ведущее учреждение:
Федеральное государственное бюджетное учреждение Научно-исследовательский институт нормальной физиологии им. П.К. Анохина РАМН.
Защита состоится «27» мая 2013 года в 14.00 часов на заседании диссертационного совета Д 208.072.05 при ГБОУ ВПО РНИМУ им. Н.И. Пирогова Минздрава России по адресу: 117997, г. Москва, ул. Островитянова, д.1.
С диссертацией можно ознакомиться в библиотеке ГБОУ ВПО РНИМУ им. Н.И. Пирогова Минздрава России по адресу: 117997, г. Москва, ул. Островитянова, д.1.
Автореферат разослан «15» февраля 2013 года.
Ученый секретарь диссертационного совета
кандидат медицинских наук, доцент Т.Е. Кузнецова
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность. Феномены стресса и адаптации к нему являются одной из проблем, стоящих перед современной физиологией и медициной. Несмотря на большое количество исследований и публикаций, посвященных различным аспектам изучения данной проблемы, в литературе до настоящего времени все еще нет исчерпывающих сведений, содержащих целостное представление о стрессе как физиологическом процессе, о механизмах его возникновения, формирования, стрессустойчивости; кроме того не выработаны подходы к профилактике негативных последствий стресса.
В 1936 году H. Selye, выдвинув теорию об адаптационном синдроме, показал, что при любом стрессовом состоянии в организме возникает трехфазный процесс. При этом в условиях стресса основные сдвиги в регулирующих гомеостаз организма системах возникают со стороны эндокринных органов: гипоталамуса, гипофиза и надпочечников (Селье Г., 1960).
Гипоталамо-гипофизарно-надпочечниковая система образует ось стресс-системы, а основными ее гормонами являются глюкокортикоиды (ГК). В настоящее время появились дополнительные сведения, свидетельствующие о существовании единой саморегулирующейся системы, лимитирующей чрезмерную стресс-реакцию. В ее структуру включаются ГАМК-, опиоидергическая и моноаминоксидазная системы, а также субстанция Р, нейропептид Y и локальные компоненты, представленные простагландинами, аденозином и антиоксидазными системами (Пшенникова М.Г., 2001).
Однако известно, что при любом виде стресса как остром, так и хроническом (включая инфекционный) происходит уменьшение размеров тимуса, преимущественно за счет снижения количества тимоцитов, а в ряде случаев наблюдается жировое перерождение этого органа (Зайчик А.Ш., Чурилов Л.П., 2005; Ивановская Т.Е., Леонова Л.В., 1989; Пшенникова М.Г., 2001; Селье Г., 1960; Baatar D. et al., 2011; Choi D.C. et al., 2009; Gruver A.L. et al., 2008; Hammar J.A., 1936). При этом ни тимус, ни продуцируемые им полипептиды в число компонентов указанной системы не включаются, хотя известно, что между тимусом и гипоталамо-гипофизарной-надпочечниковой системой существуют теснейшие связи (Кемилева З., 1984; Фабрис Н., 1980; Ярилин А.А., Беляков И.М., 1996; Jiang J. et al., 2010; Jordan J., 1981; Martin J., 1984; Mizoguchi Y. et al., 2007).
Из гормонов периферических эндокринных желез основная роль в регуляции деятельности тимуса принадлежит ГК (Гюллинг Э.В., Самбур М.Б., 1986; Martin J., 1984; Pierpaoli W. et al., 1977; Talaber G. et al., 2011; Youn D.Y. et al., 2011 и др.). Доказательством послужили эксперименты с экстрактом тимуса, адреналэктомией и тимэктомией (Кемилева З., 1984; Поповичи Д., Сэхляну В., 1969; Comsa J., 1938). Имеются указания об ингибиторном влиянии тимических факторов полипептидной природы на адренокортикальную функцию надпочечников (Алиев М.Г., Курбанов Т.Г., 1987). Основываясь на опубликованных данных можно полагать, что тимус является весьма важным органом не только иммунной, но и стресс-лимитирующей системы.
Все это делает проблему анализа роли полипептидов тимуса в регуляции стресса актуальной не только для физиологии, фармакологии, но и в целом для медицины.
Цель исследования
Целью работы явилось изучение влияния полипептидов тимуса на формирование эмоционального стресса и участие их в деятельности стресс-лимитирующей системы.
Задачи исследования
- Исследовать уровень сывороточной тимической активности, являющейся показателем функциональной деятельности тимуса, в парах мать-дитя при родовом стрессе.
- Проанализировать влияние полипептидного препарата тимуса тактивина на ориентировочно-исследовательское поведение и двигательную активность животных. Сравнить полученные результаты с действием эталонного анксиолитического препарата диазепама и классического ноотропного препарата пирацетама.
- Проанализировать поведение животных под действием полипептидного препарата тимуса тактивина в тестах на тревожность. Сравнить полученные результаты с действием эталонного анксиолитического препарата диазепама и классического ноотропного препарата пирацетама.
- Изучить влияние полипептидов тимуса на формирование у животных пищевых условных рефлексов. Сравнить полученные результаты с действием эталонного анксиолитического препарата диазепама и классического ноотропного препарата пирацетама.
- Изучить влияние полипептидов тимуса на формирование у животных оборонительных условных рефлексов. Сравнить полученные результаты с действием эталонного анксиолитического препарата диазепама и классического ноотропного препарата пирацетама.
- Исследовать динамику восстановления реакции избегания у подопытных животных под влиянием полипептидов тимуса на фоне эмоционального стресса, вызванного обратимыми функциональными нарушениями памяти. Сравнить полученные результаты с действием эталонного анксиолитического препарата диазепама и классического ноотропного препарата пирацетама.
- Исследовать с помощью метода радиолигандного связывания ex vivo влияние тактивина на экспрессию ГАМКА-рецепторов в гипоталамусе крыс.
- Исследовать изменения распределения ГАМК и моноаминов (норадреналина, серотонина, дофамина и их метаболитов) под влиянием тактивина во фронтальной коре и структурах промежуточного мозга крыс в условиях эмоционального стресса.
Научная новизна
Впервые сформулировано положение об участии тимуса в реализации родового стресса.
Впервые на основании комплекса проведенных исследований установлено, что только у здоровых рожениц с неотягощенным анамнезом при физиологически протекающей беременности имеет место адекватная реакция организма на родовой стресс в виде повышения уровня сывороточной тимической активности (СТА).
Помимо этого впервые установлено, что у рожениц, беременность которых протекала в условиях ремиссии хронических соматических или гинекологических заболеваний и/или на фоне осложненного течения данной беременности токсикозом, угрозой прерывания или других осложнений беременности, выявлено статистически значимое снижение уровня СТА по сравнению со здоровыми роженицами. Данный факт может косвенно указывать на избыточную продукцию надпочечниками гормонов стресса у данной группы беременных, что способствует снижению функциональной активности тимуса.
Установлено, что у здоровых новорожденных детей уровень СТА не зависел от стояния здоровья матери и уровня ее сывороточной тимической активности.
В эксперименте на животных впервые показано, что полипептидный препарат тимуса тактивин оказывает мнемотропный эффект, сопоставимый с действием классического ноотропного препарата пирацетама, что выражается в ускорении выработки пищевых и оборонительных условных рефлексов.
На моделях нарушения памяти было также впервые показано, что полипептиды тимуса (тактивин) улучшают воспроизведение реакции избегания в условиях эмоционального стресса. Установлены ноотропные и стресс-лимитирующие эффекты препарата тимуса и обоснована возможность предупреждения с его помощью негативных последствий стрессовых воздействий на организм животных.
Впервые проанализированы нейрохимические механизмы действия полипептидного препарата тимуса (тактивина) на различные структуры мозга на фоне эмоционального стресса. Установлено, что на фоне эмоционального стресса, вызванного обратимым функциональным нарушением реакции избегания, под действием тактивина статистически значимо увеличивается концентрация серотонина и норадреналина в гипоталамусе и фронтальной коре. Положительный эффект указанного препарата при эмоциональном стрессе заключается в изменении соотношения баланса серотонин/норадреналин в пользу первого. Участие ГАМК-ергической системы в реализации стресспротекторного действия тимических пептидов не установлено.
Проведенные исследования позволили разработать новую гипотетическую модель работы стресс-лимитирующей системы, которая учитывает стресспротекторные свойства полипептидов тимуса.
Практическая значимость
Полученные результаты существенно расширяют имеющиеся представления о роли полипептидов тимуса (тактивина) в регуляции центральной нервной системы и позволяют углубить фундаментальные знания об адаптации организма в условиях стресса.
Установленный факт мнемотропного и стресспротекторного действия полипептидного препарата тимуса тактивина (разрешенного к применению в клинической практике как иммунотропного препарата), предупреждающего развитие негативных последствий эмоционального стресса у животных, позволяет рекомендовать его для дальнейших исследований в клинической практике для коррекции различных когнитивных расстройств и изучения профилактического стресспротекторного действия препарата у человека, подвергающегося выраженным стрессогенным воздействиям.
Положения, выносимые на защиту
- Сывороточная тимическая активность (интегральный показатель функциональной активности тимуса) в крови здоровых рожениц с неотягощенным течением беременности статистически значимо выше, чем у здоровых небеременных женщин детородного возраста, что является адекватной реакцией организма на родовой стресс. Установлен факт более низкого уровня СТА у рожениц с отягощенным анамнезом и/или с отягощенным течением беременности по сравнению со здоровыми роженицами с неотягощенным течением беременности, который может указывать на истощение компенсаторных возможностей организма.
- Полипептидный препарат тимуса тактивин помимо иммунотропных свойств проявляет мнемотропный эффект на моделях оборонительных и пищевых условных рефлексов, ускоряя их выработку, что наиболее ярко проявляется на начальных этапах формирования памятного следа.
- Полипептидный препарат тимуса тактивин снижает степень нарушения воспроизведения навыков при эмоциональном стрессе, вызываемом сбоем и пространственной переделкой реакции избегания, предупреждая негативные проявления эмоционального стресса, что может говорить о наличии у него стресс-протекторной активности.
- Тактивин изменяет баланс моноаминергических систем мозга после сбоя реакции избегания и меняет соотношение серотонин/норадреналин в пользу первого, что лежит в основе его стресс-протекторного действия.
Внедрение в практику
Результаты исследования внедрены в учебный процесс кафедры фармакологии педиатрического факультета Российского национального исследовательского медицинского университета имени Н.И. Пирогова Министерства Здравоохранения Российской Федерации, кафедры педиатрии медицинского факультета Российского университета дружбы народов и в научную работу лаборатории эволюции механизмов памяти кафедры высшей нервной деятельности Биологического факультета Московского государственного университета им. М.В. Ломоносова.
Апробация работы и публикации
Основные положения диссертации обсуждались на Международной конференции «Физиология и патология иммунной системы» и IV Международной конференции по иммунотерапии (Москва, 2008); VII съезде аллергологов и иммунологов СНГ и II Всемирном форуме по астме и респираторной аллергии (С-Петербург, 2009); Х и ХI Международном конгрессе «Здоровье и образование в ХХI веке» (Москва, 2009, 2010); Международном конгрессе по реабилитации в медицине и иммунореабилитации Всемирного форума педиатров (Дубай, ОАЭ, 2009); XVI и XVII Российском национальном конгрессе «Человек и лекарство» (Москва, 2009; 2010); XIV Международном конгрессе по реабилитации в медицине и иммунореабилитации (Тель-Авив, Израиль, 2009); 5-ой Международной конференции «Биологические основы индивидуальной чувствительности к психотропным средствам» (Клязьма, 2010); XXI Съезде физиологического общества им. И.П. Павлова (Калуга, 2010); Всероссийской конференции с международным участием, посвященной 85-летию со дня основания Института физиологии им. И.П. Павлова РАН (С-Петербург, 2010); XV конгрессе педиатров России с международным участием «Актуальные проблемы педиатрии» (Москва, 2011); XVI Международном конгрессе по реабилитации в медицине и иммунореабилитации (Париж, Франция, 2011); II Международной научно-практической конференции «Фундаментальные и прикладные проблемы стресса» (Витебск, Беларусь, 2011); VI Всемирном конгрессе по иммунопатологии и респираторной аллергии/VIII Конгрессе по аллергологии и иммунологии стран СНГ/V съезде иммунологов России (Москва, 2011); III Съезде физиологов СНГ (Ялта, Украина, 2011); V городской научно-практической конференции «Эндокринологические аспекты в педиатрии» (Москва, 2011).
Диссертация апробирована на совместном заседании кафедры фармакологии педиатрического факультета ГБОУ ВПО Российский национальный исследовательский медицинский университет имени Н.И. Пирогова Минздрава России, кафедры высшей нервной деятельности Биологического факультета Московского государственного университета имени М.В. Ломоносова и лаборатории молекулярной иммунологии и биохимии ФГБУН «НИИ Физико-химической медицины» ФМБА России.
По основным результатам диссертации опубликовано 53 работы, в том числе 18 статей в изданиях, рекомендованных ВАК РФ. Результаты исследования вошли также в пособие для врачей «Иммунокоррекция тактивином в комплексном лечении часто болеющих детей».
Объем и структура диссертации.
Диссертации изложена на 220 страницах машинописного текста и состоит из введения, обзора литературы (1-я глава), материалов и методов исследования (2-я глава), 5 глав результатов собственных исследований (3-7-я главы), обсуждения результатов (8-я глава), заключения, выводов, практических рекомендаций. Библиографический указатель включает 179 источника отечественной и 206 иностранной литературы. Работа проиллюстрирована 15 таблицами и 51 рисунками.
СОДЕРЖАНИЕ РАБОТЫ
Материалы и методы исследования
Анализ показателей сывороточной тимической активности
Проанализированы уровни СТА в 112 парах мать-ребенок в момент родового стресса. В качестве контроля анализировали уровень СТА здоровых не беременных женщин детородного возраста (n=10). Кровь для анализа брали у женщин из кубитальной вены. Для исследования СТА ребенка забор крови проводили из пупочной вены, избегая смешивания с материнской кровью.
СТА определяли по известному методу (Bach J.F. et al., 1975) в модификации Arion V.Ya. с соавторами (2001). В основе данного метода лежит способность азатиоприна ингибировать процесс Т-клеточного розеткообразования лимфоцитов, выделенных из селезенки молодых мышей. Данный процесс зависит от функционального состояния тимуса, так как Т-розеткообразующие лимфоциты селезенки взрослых тимэктомированных мышей менее чувствительны к азатиоприну, чем Т-лимфоциты интактных животных. Чувствительность Т-лимфоцитов селезенки тимэктомированных мышей к азатиоприну восстанавливается после их обработки in vitro тимическими экстрактами, человеческой сывороткой крови, сывороткой крови животных или цинк-тимулином. Так как в сыворотке крови присутствуют тимические пептиды, и, в частности, цинк-тимулин, то использование данного метода позволяет судить о функциональном состоянии тимуса (Consolini R. et al., 2000; Mezza C. et al., 2007). Данный биотест в настоящее время является единственным методом для выявления иммуноактивных молекул тимуса, циркулирующих в крови (Mezza C. et al., 2007).
Для постановки теста к 0,025 мл рабочего раствора азатиоприна (50 мкг/мл) добавляли 0,1 мл фильтрата плазмы, последовательно разведенной в соотношениях 1:4; 1:8; 1:16; 1:32 и т.д., и 0,05 мл суспензии клеток селезенки тимэктомированных мышей (1,5 106 клеток). Контрольные пробы вместо 0,025 мл раствора азатиоприна содержали 0,025 мл буфера для азатиоприна, разведенного средой 199 (исходный раствор: 0,2 мл буфера для азатиоприна + 3,8 мл среды 199). После 90 мин. инкубации при 37C в смесь добавляли 0,05 мл 1% взвеси эритроцитов барана. Таким образом, конечный объем пробы составлял 0,225 мл. Смесь центрифугировали 5 мин. при 200 g при 4C и оставляли в холодильнике при той же температуре на 1-18 ч., далее производили подсчет результатов.
Результаты выражались в виде log2 N, где log2 – это логарифм по основанию 2, а N – разведение добавляемой в инкубационную среду сыворотки (при этом объем инкубационной среды всегда постоянный). При этом log2 N = 1 если в инкубационную среду добавлялась неразведенная сыворотка, то есть титр равен 0; log2 N = 2 добавлялась сыворотка, разведенная в 4 раза, то есть титр равен 1:4 и т. д.
Экспериментальные исследования на животных
Эксперименты проводились на 510 крысах линии Wistar весом 180-200 г. Крысы были получены из питомника лабораторных животных «Пущино» (Московская обл., г. Пущино). Животных содержали в стандартных условиях вивария. Все эксперименты проводили согласно «Правилам проведения работ с использованием экпериментальных животных» (приказ Минвуза № 724 от 13.11.84 г.) и положениям международной конвенции о «Правилах работы с экспериментальными животными» (European Communities Council Directives, November 24, 1986, 86/609/EEC). В работе было выполнено 9 серий экспериментов (табл. 1).
В I-IV сериях было по 4 группы животных. В течение пяти дней внутрибрюшинно один раз в сутки (во второй половине дня) животным вводись следующие препараты объемом 0,5 мл: 0,9% раствор NaCl (контрольная группа); тактивин – в дозе 0,5 мг/кг; пирацетам – эталонный ноотропный препарат в дозе 300 мг/кг; диазепам – эталонный анксиолитик в дозе 0,5 мг/кг.У животных данных серий перед началом введения препаратов тестировали ориентировочно-исследовательское поведение и уровень двигательной активности в норковой камере и открытом поле для формирования идентичных по поведению групп. После тестирования и формирования групп животным проводили 5-и дневный курс введения препаратов. На 5-й день животных вновь тестировали в указанных тестах. На 6-7 день животных тестировали на уровень тревожности в тесте светло-темнового выбора и приподнятом крестообразном лабиринте. На 8-й день тестировали двигательную активность животных в челночной камере и приступали к выработке условных рефлексов. В процессе формирования условных рефлексов и эмоционального стресса продолжали введение животным тактивина и физиологического раствора через день в указанных дозах.
Животным V серии вводили физиологический раствор и тактивин интраназально в объеме 100 мкл. Опытная группа животных была разделена на две подгруппы: первая получала препарат в дозе 5 мкг/кг, вторая – 0,25 мкг/кг. Препарат и физиологический раствор вводили в течение 9 дней до начала выработки пищевого условного рефлекса и продолжали вводить в течение всего эксперимента.
| Таблица 1. | Дизайн исследования | ||||
| № серии | Рефлексы и нарушения | Препарат | Кол-во животных в группе | Предварительные поведенческие тесты/ их отсутствие | |
| I | УРАИ, эмоциональный стресс, вызванный сбоем реакции избегания и пространственной переделкой навыка | 0,9% р-р NaCl Тактивин Пирацетам Диазепам | 50 50 30 30 | День 1 Норковая камера 1 Открытое поле 1 День 5 Норковая камера 2 Открытое поле 2 День 6 Тест «светло-темнового выбора» День 7 Тест «приподнятый крестообразный лабиринт» День 8 Тест «челночная камера» и выработка рефлекса | |
| II | УРПИ | 0,9% р-р NaCl Тактивин Пирацетам Диазепам | 30 30 20 20 | ||
| III | мУРПИ | 0,9% р-р NaCl Тактивин Пирацетам Диазепам | 10 10 10 10 | ||
| IV | ПУР и рефлекс на отсчет интервалов времени | 0,9% р-р NaCl Тактивин Пирацетам Диазепам | 30 30 20 20 | ||
| V | ПУР и рефлекс на отсчет интервалов времени (интраназальное введение) | 0,9% р-р NaCl Тактивин 5 мкг/кг Тактивин 0,25 мкг/кг | 10 10 10 | Открытое поле | |
| VI | Тест Порсолта | 0,9% р-р NaCl Тактивин | 10 10 | ||
| VII | Радиолигандное связывание ГАМК А - рецептора | 0,9% р-р NaCl Тактивин | 10 10 | Виварийный контроль | |
| VIII | Нейрохимический анализ | 0,9% р-р NaCl Тактивин | 10 10 | Виварийный контроль | |
| IX | Нейрохимический анализ после УРАИ и эмоционального стресса | 0,9% р-р NaCl Тактивин | 10 10 | Тесты как в I-IV сериях | |
Сокращения: УРАИ – условный рефлекс активного избегания; УРПИ – условный рефлекс пассивного избегания; мУРПИ – условный рефлекс пассивного избегания в модифицированной установке; ПУР – пищевой условный рефлекс.
VI-IX серии включали 2 группы животных, которым внутрибрюшинно водили физиологический раствор или тактивин в дозе 0,5 мг/кг соответственно. В VI серии исследовали антидепрессивные свойства препарата в тесте вынужденного плавания по Порсолту. В VII-IX сериях исследовали изменения уровня моноаминов, распределения ГАМК и экспрессию ГАМКА-рецепторов под действием тактивина в различных структурах головного мозга крыс в условиях эмоционального стресса.
Все используемые в работе поведенческие методики, методики выработки условных рефлексов, а также методики формирования функциональных нарушений рекомендованы для изучения ноотропной, анксиолитической и антидепрессивной активности фармакологических веществ (Воронина Т.А., Островская Р.У., 2005; Воронина Т.А., Середенин С.Б., 2005).
Установки для тестов «норковая камера» и «открытое поле» представляют собой квадратную (40х40х40 см) и круглую (диаметром 97 см) камеры соответственно, пол которых разделен на сектора с отверстиями. У животных в течение 5 мин. регистрировали количество пересеченных секторов, вертикальных стоек и норковых реакций, продолжительность замираний и длительность актов груминга.
Тест светло-темнового выбора проводили в камере (60х30х30 см) из оргстекла, разделенной на два отсека. Один отсек был затемнен, а стенки другого – прозрачны. В течение 5 мин. регистрировали: латентный период (ЛП) захода в темный отсек, длительность пребываний в светлом отсеке, количество пересеченных отсеков, вертикальных стоек, длительность замираний животного и продолжительность актов груминга.
Дополнительные исследования двигательной и исследовательской активности проводили в челночной камере размером 60х30х30 см, разделенной перегородкой с отверстием на два равных отсека. В течение 5 мин. регистрировали: количество пересеченных отсеков, вертикальных стоек, количество и длительность замираний животного и актов груминга. Впоследствии в этой установке вырабатывали условный рефлекс активного избегания.
Приподнятый крестообразный лабиринт (ПКЛ) состоит из расходящихся под прямым углом от центральной площадки 4 рукавов (50х14 см): двух открытых и двух со стенками высотой 30 см. Установка приподнята над полом на 50 см. В течение 5 мин. регистрировали число заходов в рукава, стоек, выглядываний из закрытых рукавов, свешиваний с открытых рукавов, количество и длительность периодов неподвижности и актов груминга.
Тест Порсолта проводили в стеклянном цилиндре диаметром 20 см и высотой 40 см, на 1/3 заполненном водой с температурой 27 ± 1С. За поведением животного наблюдали в течение 5 мин., после чего животное возвращали в клетку. В протоколе фиксировали время и количество эпизодов активного плавания и иммобильности.
Выработка пищевого условного рефлекса на наличный раздражитель и рефлекса на отсчет интервала времени. Пищедобывательный рефлекс (ПУР) вырабатывали на наличный раздражитель – тон (400 Гц) по методике, разработанной Меринг Т.А. (1988). В течение 12 ч до выработки рефлекса проводили пищевую депривацию животных. Для выработки рефлекса использовали деревянную камеру (33х45х34 см) с пластиковым полом. В одной стенке камеры на высоте 2 см от пола находилось отверстие. У этой стенки с внутренней стороны располагалась платформа, становясь на которую, крыса могла получать пищу через отверстие. Тон предъявляли в строгом временном стереотипе с интервалом 60 с. Тон действовал в течение 15 с. За это время крыса должна была подняться на платформу и взять пищу. Для анализа были выбраны следующие параметры: число верных решений (в процентах), число межсигнальных реакций, количество стоек.
После пяти дней выработки ПУР тон отменяли и проверяли способность животных к воспроизведению рефлекса на отсчет интервалов времени. Пищу подавали в отверстие с интервалом 60 с. Рефлекс считался правильным, если в интервале 60 с было не более трех межсигнальных побежек. Анализировали следующие параметры: процент своевременных подходов к кормушке и количество межсигнальных реакций.
Условный рефлекс пассивного избегания (УРПИ). Крыс поочередно на 5 мин. помещали в освещенный отсек спиной к темному отсеку (стартовое положение) и наблюдали за их поведением. На следующий день крыс снова помещали в светлый отсек, и как только крыса вступала в темный отсек, подавали на электрифицированный пол ток (0,8 мА) до момента перехода в светлый отсек, но не более 5 с. Через 24 ч., 48 ч., одну и две недели после обучения животных вновь помещали в освещенный отсек и регистрировали латентный период входа в темный отсек, отсчитывая с момента помещения животного в камеру. Тест заканчивали, когда животное входило в темный отсек или если не делало этого в течение 3 мин.
Выработку модифицированного УРПИ (Иноземцев А.Н. и др., 2007) проводили в камере размером 90х30х30 см, состоящей из трех отсеков: двух темных и центрального светлого, разделенных перегородками с отверстиями. Крыс поочередно на 5 мин. помещали в освещенную часть камеры спиной к темному отсеку (стартовое положение) и наблюдали за их поведением. На следующий день крысу снова помещали в светлый отсек, и как только она проникала в один из темных отсеков, подавали ток 0,8 мА до момента выхода из отсека, но не более 5 с. Через сутки, 3 дня, неделю и две недели после обучения крысу помещали в установку и в течение 5 мин. наблюдали за ее поведением. Регистрировали выбор того или иного темного отсека и ЛП захода в них.
Условный рефлекс активного избегания (УРАИ) вырабатывали у крыс в челночной камере. К условному раздражителю (звук 700 Гц) через 10 с изолированного действия подключался безусловный раздражитель – электрический ток силой 0,4-0,9 мА. При переходе животного через отверстие в другую половину камеры во время действия тока (реакция избавления) звук и ток выключались. Если животное переходило во время следования только звукового сигнала (реакция избегания), тока не следовало, а звук прекращался. Если животное не совершало перехода при действии раздражителей, то через 10 с после совместного действия раздражители выключались одномоментно автоматически. Каждый опыт состоял из 25 предъявлений с межсигнальным периодом 30 с. Опыты проводили ежедневно в течение 14 дней до формирования устойчивого рефлекса (более 80% реакций избегания от числа предъявлений). Фиксировалось количество реакций избегания, избавления и межсигнальных реакций (МСР).
Для формирования эмоционального стресса в работе использовали два приема функциональных нарушений УРАИ, которые вызывались внезапными изменениями условий опыта: сбой и пространственную переделку навыка. За 45 мин. до проведения функциональных нарушений животным вводили препараты.
Сбой реакции избегания. После достижения животными критерия обученности УРАИ (80% реакций избегания) в опыт вводили следующие изменения: перебегание крысы в другую половину камеры не приводило к выключению раздражителей, что повторялось при 5 побежках; таким образом, животное подвергалось воздействию электрическим током в любой половине камеры (Иноземцев А.Н., Прагина Л.Л., 1989). После пятой реакции ток выключался немедленно, а звук – спустя 2 с. Затем в течение 20 предъявлений тестировали уровень воспроизведения реакций избегания в прежних условиях.
Функциональное нарушение, вызванное пространственной переделкой реакции избегания. Сбой является обратимым функциональным нарушением, что позволяет на одних и тех же животных последовательно изучать влияние препаратов сначала в условиях одной модели, а затем в условиях другой, получая при этом воспроизводимость фармакологического эффекта (Гарибова Т.Л. и др., 2005). Основываясь на этом, мы через день после сбоя восстанавливали УРАИ до критерия обученности, закрывали отверстие через которое животное переходило в другой отсек камеры и открывали отверстие в противоположном углу перегородки, в течение 20 предъявлений тестировали воспроизведение реакции в новых условиях (Иноземцев А.Н., Прагина Л.Л., 1991).
Радиолигандный анализ ГАМКА-рецепторов (эксперименты проводились совместно с лабораторией радиоизотопных методов исследований НИИ Фармакологии им. В.В. Закусова РАМН, руководитель д.м.н., профессор Г.И. Ковалев). После вывода интактных крыс из эксперимента, проводили декапитацию и из головного мозга выделяли гипоталамус и фронтальную кору. Приготовление мембранных препаратов, содержащих ГАМКА-рецепторы в указанных структурах проводили по модифицированным методам (Ito Y. et al., 1992; Hawkinson J.E. et al., 1996). После выделения структуры мозга замораживали в жидком азоте и хранили в низкотемпературном холодильнике при -85С. В день эксперимента структуры гомогенизировали в 20 мл ледяного буфера (0,32 М сахароза; рН 7,1) в гомогенизаторе “Potter S” (Sartorius Stedim, Германия) в системе «стекло-тефлон». Полученный гомогенат центрифугировали в ультрацентрифуге “Optima L-70K” (Beckman Coulter, Великобритания) при 1000 g 10 мин. Супернатант повторно центрифугировали при 20000 g 20 мин. Осадок ресуспендировали в 20 мл холодной дистиллированной воды и вновь центрифугировали при 8000 g 20 мин. Супернатант и желтый надосадочный слой центрифугировали при 48000 g 20 мин. Далее осадок суспендировали в 0,05 М Tris-цитратном буфере (pH 7,1) и повторно центрифугировали при 48000 g 20 мин. Полученную мембранную фракцию замораживали и хранили при -85С. В день эксперимента мембраны суспендировали в 40 объёмах Tris-цитратного буфера и центрифугировали при 48000 g 20 мин. Осадок суспендировали в 40 объёмах буфера, инкубировали при 24С 30 мин. и снова центрифугировали при 48000 g 20 мин. Концентрацию белка в образцах определяли по стандартной методике Lowry О.Н. (1951).
Радиолигандный анализ ex vivo проводили по модифицированной методике (Ito Y. et al., 1992; Heaulme M. et al., 1986). При изучении рецепторного связывания использовали меченный селективный лиганд для ГАМКА-рецепторов – [G3H]SR95531, с удельной радиоактивностью 50 кюри/ммоль, полученный методом твердофазного катализа в отделе химии физиологически активных веществ Института молекулярной генетики РАН. Инкубационная смесь (конечный объем 0,5 мл) содержала 50 мкл [G3H]SR95531 (в диапазоне концентраций от 0,1 до 30 нМ), 250 мкл Tris-цитратного буфера (pH 7,1) и 200 мкл суспензии мембран. Смесь инкубировали при 0С в течение 45 мин. По окончанию инкубации пробы фильтровали через стекловолокнистые фильтры GF/D (Whatman, Великобритания). Каждую пробирку промывали два раза холодным буфером, затем фильтры промывали еще два раза тем же объемом буфера. Фильтры просушивали на воздухе и помещали в сцинтилляционные флаконы, которые заливали 5 мл сцинтилляционной жидкостью на основе толуола (4 г РРО, 0,2 г РОРОР на 1 л толуола). Радиоактивность определяли на счетчике Tri-Carb 2900TR (Percin Elmer, Швейцария) с эффективностью счета 42-46%.
Неспецифическое связывание определяли в присутствии 50 мкл немеченого лиганда (+Bicuculline), в концентрации 0,1 мкМ. Специфическое связывание рассчитывали как разницу между общим и неспецифическим связыванием.
Результаты экспериментов ex vivo оценивали с помощью рассчитанных величин Кd и Вmax, отражающих степень сродства рецептора к лиганду (нМ) и количество мест связывания лиганда (фмоль/мг белка), соответственно.
Для обработки результатов радиолигандного связывания использовали программу Graphpad Prism 4 Demo.
Биохимическое определение содержания моноаминов в мозге крыс (эксперименты проводились совместно с лабораторией нейрохимической фармакологии НИИ Фармакологии им. В.В. Закусова РАМН, руководитель к.м.н., вед.н.с. В.С. Кудрин). Крыс после эмоционального стресса, вызванного сбоем реакции избегания, выводили из эксперимента. Головной мозг извлекали на льду, выделяли фронтальную кору, гипоталамус, гиппокамп, стриатум, прилежащее ядро и замораживали в жидком азоте. Перед определением содержания нейротрансмиттеров пробы размельчали в гомогенизаторе (тефлон–стекло) в 20 объемах 0,1 М хлорной кислоты и центрифугировали при 10000 g в течение 10 мин. Содержание моноаминов определяли методом высокоэффективной жидкостной хроматографии (ионпарная хроматография) с электрохимической детекцией на хроматографе LC-304T (BAS, West Lafayette, США) с обращенно-фазной колонкой ReproSil-Pur, ODS-3, 4х100 мм (длина волны возбуждения – 230 нм, длина волны испускания – 392 нм) (Кудрин В.С. и др., 1988).
Статистическую обработку полученных данных проводили с применением программы STATISTICA 6.0 (Statsoft, США), используя непараметрический критерий Вилкоксона (поведенческие эксперименты), Манна-Уитни (биохимические данные), а также c использованием F-критерия Фишера (радиолигандный анализ). Статистическая обработка результатов исследования СТА проводилась методом вариационной статистики с определением средней арифметической (М), стандартного отклонения (), стандартной ошибки средней (m). Достоверность различий определялась по критерию Стьюдента (t).
ОСНОВНЫЕ РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Анализ литературных источников показывает, что гормоны и полипептиды тимуса во многом проявляют действие, противоположное ГК, одним из ведущих гормонов стресс-системы. Ни тимусу, ни его регуляторным гормонам не нашлось места в современных схемах работы стресс- и стресс-лимитирующей систем (Пшенникова М.Г., 2001), хотя еще Селье Г. (1936) показал, что основные сдвиги при любом стрессовом состоянии возникают преимущественно со стороны гипофиза, надпочечников и тимуса. В то же время хорошо известен факт, что тимус тонко реагирует на различные физиологические и патологические воздействия. Авторы, исследующие тимус, подчеркивают, что акцидентальная инволюция тимуса (атрофия тимуса) является ни чем иным, как проявлением адаптационного синдрома в ответ на стрессовое воздействие (Mller-Hermelink H., 1986; Yarilin A.A., 1999). Таким образом, тимус рассматривается как орган, играющий ведущую роль только в регуляции иммунитета. Однако накопленные факты позволяют предположить его регулирующую роль при различных стрессовых воздействиях, осуществляемую через выделяемые пептиды, которые по своей сути обладают свойствами гормонов.
В течение всего 20-го века исследователи, изучавшие тимус, пытались использовать его экстракты, содержавшие гормоны и полипептиды тимуса, как для изучения влияния тимуса на организм, так и для лечения ряда заболеваний. В 70-80 годы прошлого века в США, Румынии, СССР из тимуса был выделен ряд препаратов: тимозин (Гольдштейн А., США), полипептиды и стерины (румынская школа исследователей), тималин (Морозов В.Г., Хавинсон В.Х., С-Петербург), тактивин (Арион В.Я., Москва). В настоящее время существуют факты успешного использования препаратов тимуса, в том числе и тактивина, в клинической практике в качестве иммуномодуляторов. В нашей работе мы исследовали действие тактивина (препарата низкомолекулярных полипептидов тимуса), обладающего иммунотропной активностью, при различных стрессорных воздействиях.
Анализ показателей сывороточной тимической активности
в парах мать-дитя при родовом стрессе
На первом этапе исследования наше внимание привлек родовой стресс. Поскольку, вероятно, пептиды тимуса участвуют в реализации стресса, мы исследовали сывороточную тимическую активность, отражающую уровень тимических гормонов в периферической крови и как следствие функциональную активность тимуса. В исследовании участвовали женщины, родившие здоровых доношенных детей (38-40 недель гестации).
В зависимости от состояния здоровья, особенностей соматического и акушерско-гинекологического анамнеза и течения беременности матери были разделены на четыре группы: 1-я группа (n=19) – здоровые матери с неотягощенным анамнезом и нормально протекавшей беременностью, средний возраст которых составлял 24,0±4,0 года; 2-я группа (n=14) – матери с отягощенным соматическим и/или акушерско-гинекологическим анамнезом, настоящая беременность у которых протекала также нормально, средний возраст которых составлял 25,0±5,1 года; 3-я группа (n=20) – матери с благоприятным соматическим и акушерско-гинекологическим анамнезом, но с осложненным течением беременности в виде раннего токсикоза и/или повышенного тонуса матки, средний возраст – 24,9±3,6 года; 4-я группа (n=59) – матери с отягощенным соматическим и/или акушерско-гинекологическим анамнезом и осложненным течением беременности, в числе которых такие же осложнения, как у матерей детей 3-й группы, средний возраст – 26,0±5,4 года. Группой сравнения (5-я группа – n=10) служили здоровые не беременные женщины, средний возраст которых составлял 30,0±8,1 года.
У матерей 1 группы уровень СТА был статистически значимо выше, чем аналогичный показатель у здоровых не беременных женщин (log2N=3,9±0,3 vs. log2N=2,6±0,1, p<0,001). Таким образом, полученные нами результаты подтвердили данные, выявленные ранее в 1991 году Лабунец И.Ф. и соавт., что исследуемый показатель статистически значимо повышается перед родами. Обращает на себя внимание факт, что у матерей 1 группы уровень СТА в сыворотке крови, взятой из кубитальной вены сразу после рождения ребенка, был статистически значимо выше (p<0,001), чем у матерей 2, 3 и 4 групп с патологическим и/или отягощенным течением беременности (табл. 2). Помимо этого установлено, что в парах мать-дитя в 3 и 4 группах уровень СТА у детей статистически значимо был более высоким, чем у их матерей (p<0,01 и p<0,001, соответственно).
По данным Лабунец И.Ф. и соавт. (1991), уровень аналогичного показателя в I и II триместре беременности не отличается от уровня здоровых небеременных женщин. Статистически значимые отличия появляются лишь в III триместре с максимумом непосредственно перед родами. Этот показатель сохраняется на высоком уровне после естественных родов у здоровых рожениц. Такое повышение уровня СТА, на наш взгляд, является адекватной реакцией тимуса на родовой стресс.
| Таблица 2. | Распределение значений СТА в пуповинной крови доношенных новорожденных детей и в крови их матерей, взятой из кубитальной вены сразу после рождения ребенка |
| Наблюдавшиеся группы | Число наблюдений | Уровень СТА (log2 N), M ± m | Уровень значимости различия показателей СТА в парах мать-ребенок, р | |
| женщины | дети | |||
| 1 | 19 | 3,9±0,3 | 2,8±0,3 | р<0,05 |
| 2 | 14 | 2,1±0,3** | 2,7±0,2 | р>0,05 |
| 3 | 20 | 2,5±0,2** | 3,3±0,2 | р<0,01 |
| 4 | 59 | 2,2±0,1** | 2,7±0,1 | р<0,001 |
| (2±3±4)* | 93 | 2,3±0,2** | 2,9±0,25 | р>0,05 |
| 5 | 10 | 2,6±0,1** | ||
*среднее значение уровня СТА во 2, 3, 4 группах, взятых вместе.
**р<0,001 по сравнению с 1 группой
У рожениц, беременность которых протекала в условиях ремиссии хронических соматических или гинекологических заболеваний и/или на фоне осложненного течения беременности (токсикоз, угрозой прерывания или других осложнений беременности), уровень СТА в родах не отличался от уровня не беременных женщин и был ниже уровня СТА у здоровых рожениц, беременность которых не была осложнена. Данный факт позволяет полагать, что у женщин 2-4 групп имеет место хронический стресс, лежащий в основе избыточной продукции ГК. Последняя способствует снижению функциональной активности тимуса в момент родового стресса. Результаты исследования, представленные в таблице 2, указывают на отсутствие зависимости функциональной активности тимуса новорожденных детей от функциональной активности тимуса матери. Следовательно, тимус новорожденного ребенка, а также плода (по крайне мере в период, предшествующий акту родов) функционально активен и работает самостоятельно вне зависимости от функциональной активностью тимуса его матери.
Экспериментальные исследования на животных
Следующим этапом исследования явилось изучение действия полипептидов тимуса (тактивина) на поведение крыс в различных по стрессогенности ситуациях.
В качестве препаратов сравнения были использованы ноотропный препарат пирацетам и классический бензодиазепиновый анксиолитик диазепам. Ноотропные препараты обладают не только способностью улучшать память и обучение животных, но и анксиолитической и адаптогенной активностью (Нисс А.И., 1984). Для исследования ноотропной активности веществ нередко прибегают к стрессогенным воздействиям (Воронина Т.А., Островская Р.У., 2005; Иноземцев А.Н., Прагина Л.Л., 1992). Таким образом, применение пирацетама в качестве препарата сравнения в данной работе было вполне оправданным. В качестве второго эталонного препарата мы применяли диазепам, основным фармакологическим эффектом которого является устранение тревоги и эмоционального напряжения.
Для создания целостной картины, свидетельствующей об участии тимуса в ограничении стресса, нами была использована целая палитра поведенческих тестов с разной степенью стрессогенности.
Влияние тактивина на ориентировочно-исследовательское поведение, двигательную активность и тревогу
Методики открытого поля и норковой камеры широко используются в экспериментальной психофармакологии для изучения влияния веществ на ориентировочно-исследовательское поведение и двигательную активность (Воронина Т.А., Островская Р.У., 2005). Всех интактных животных тестировали в «открытом поле» и норковой камере за сутки до введения препаратов. По результатам первого тестирования формировали равноценные по поведению экспериментальные группы. Результаты тестирования интактных крыс принимали за 100% (фоновые значения). Повторное тестирование в этих установках проводили после 5 дней введения препаратов. При втором тестировании во всех группах происходило снижение горизонтальной двигательной активности животных (рис. 1,2), что отражает их привыкание к установке.
В открытом поле введение тактивина, пирацетама и диазепама привело к статистически значимому снижению количества посещений периферических секторов до 44,2 ± 3,4%, 53,3 ± 3,1%, 46,4 ± 3,2%, соответственно (р<0,05), в то время как в контрольной группе этот показатель снижался лишь до 66,0±9,5% от уровня фоновых значений (рис. 1). Снижение количества посещений центрального сектора на фоне тактивина и диазепама достигало 50,0±9,3% и 55,3±9,5, соответственно, на фоне пирацетама этот показатель составил 62,3±7,7% (р<0,05), а наиболее выраженное снижение до 36,4±15,5% мы наблюдали в контрольной группе.

Рисунок 1. Поведение животных в открытом поле (% по отношению к результатам 1-го тестирования).
*р<0,05; **р<0,001 относительно контроля.
Количество норковых реакций также снижалось во всех группах по сравнению с 1-ым тестированием. Однако, только на фоне тактивина данное снижение было статистически значимо ниже контроля (59,0±2,7% vs. 71,75±4,5%, р<0,05), в то время как на фоне препаратов сравнения оно не отличалось от контроля (пирацетам – 72,5±4,4%; диазепам – 75,8±5,5%). Кроме этого только на фоне тактивина продолжительность реакции одной остановки снижалась относительно исходного уровня и составляла 82,5±10,7% (р<0,001), в то время как в других группах она увеличивалась по сравнению с первым тестированием на фоне пирацетама и диазепама до 104,8±6,2 (р<0,001) и 113,0±7,1%, (р<0,05) соответственно, а в контроле достигала 166,4±21,0%.
В норковой камере на фоне тактивина наблюдали снижение горизонтальной двигательной активности до 61,0±4,6% (р<0,05), в то время как в контроле она снижалась до 49,2±5,2%, а на фоне эталонных препаратов пирацетама и диазепама до 34,9±2,27% (р<0,05) и 47,2±2,6%, соответственно. Под влиянием тактивина помимо большей горизонтальной двигательной активности наблюдалось и большее количество стоек (63,3±4,2%, р<0,05), превышающее данный показатель в остальных группах (контроль – 50,8±5,8%; пирацетам – 33,3±2,9% (р<0,05); диазепам – 42,5±4,9%). Длительность одной остановки увеличивалась во всех группах животных, но на фоне тактивина и диазепама этот показатель был статистически значимо ниже уровня контроля и составлял (189,3±19,7% и 196,5±25,9%, соответственно; р<0,05), в то время как в контрольной группе средняя продолжительность одной остановки составила 286,2±32,8%, а на фоне ноотропа – 269,3±19,4% относительно значений первого тестирования (рис. 2).
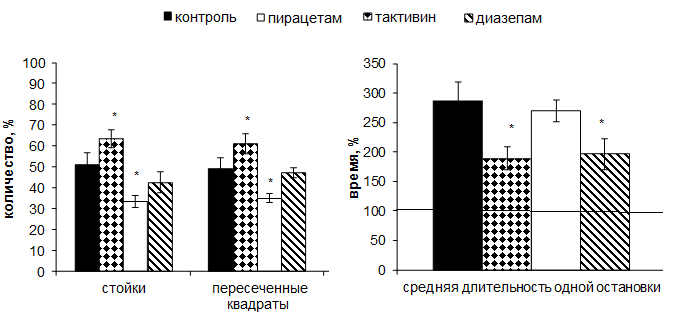
Рисунок 2. Поведение животных в норковой камере (% по отношению к результатам 1-го тестирования).
*р < 0,05 относительно контроля.
Ориентировочно-исследовательское поведение изучали также в челночной камере, в которой в дальнейшем вырабатывали УРАИ и проводили его функциональные нарушения. На фоне тактивина количество переходов крысы в течение 5 мин. в другую часть камеры превышало контрольный уровень (8,3±0,8 и 6,4±0,6, соответственно; р<0,05), при этом эффект у эталонных препаратов отсутствовал (пирацетам – 5,4±1,0; диазепам – 6,2±1,3). Уровень вертикальной активности на фоне тактивина (14,6±1,5 стоек) и диазепама (12,7±1,7 стоек) не отличался от контроля (12,4±1,1 стоек). Только на фоне действия пирацетама данный показатель превышал контрольный уровень (16,6±1,9 стоек, р<0,05) (рис. 3).
Помимо изучения ориентировочно-исследовательской активности проводили исследование тревожности крыс в приподнятом крестообразном лабиринте (ПКЛ) и тесте светло-темнового выбора. В ПКЛ тактивин, в отличие от препаратов сравнения, не проявил анксиолитической активности: посещение светлых рукавов и центральной площадки животными не отличалось от контроля. В тесте светло-темнового выбора тактивин, в отличие от пирацетама, также не продемонстрировал анксиолитической активности (табл. 3).

Рисунок 3. Поведение животных в челночной камере.
*р < 0,05 относительно контроля
.
| Таблица 3. | Данные тестов ПКЛ и светло-темнового выбора | |||||
| Группа | Кол-во животных | ПКЛ | Тест светло-темнового выбора | |||
| Светлые рукава | Центр | Латентный период захода в темный отсек (сек) | Суммарное время, проведенное в светлом отсеке (сек) | |||
| Контроль | 120 | 1,59±0,30 | 2,88±0,75 | 5,20±0,6 | 47,00±6,8 | |
| Тактивин | 120 | 1,68±0,15 | 3,25±0,50 | 5,37±0,48 | 49,60±5,6 | |
| Диазепам | 80 | 4,35±0,50* | 5,55±0,50* | 6,05±1,1 | 37,60±6,75 | |
| Пирацетам | 80 | 5,80±0,80* | 6,40±1,90* | 4,31±0,23* | 66,80±5,80* | |
*р<0,05 относительно контроля.
Таким образом, тактивин в открытом поле увеличивал относительно контроля центральную двигательную активность и уменьшал время одной остановки, что сопоставимо с эффектом диазепама и свидетельствует об уменьшении тревоги животных в данном тесте. Кроме того, полипептиды тимуса уменьшали число норковых реакций в открытом поле относительно контроля, что также свидетельствует об уменьшении тревоги в данном тесте. Следует подчеркнуть, что тактивин также активирует ориентировочно-исследовательское поведение в норковой и челночной камерах. Эти результаты, сопоставимые с действием эталонных препаратов, свидетельствуют о влиянии пептидов тимуса на функции ЦНС. По данным ряда авторов, с количественными показателями ориентировочно-исследовательского поведения коррелирует также индивидуальная устойчивость к эмоциональному стрессу. При этом низкую двигательную активность связывают с низкой стрессустойчивостью (Герштейн Л.М., 2001; Маркова Е.В., 2011; Юматов Е.А., Мещерякова О.А., 1990).
Влияние тактивина на выработку условных рефлексов
Влияние тактивина на выработку классического условного рефлекса пассивного избегания (УРПИ). Для доклинического изучения влияния различных веществ на процессы обучения и памяти модель УРПИ на животных считается наиболее удобной (Воронина Т.А., Островская Р.У., 2005). Опыты проводили в камере, состоящей из дух отсеков: светлого и темного. При обучении животное получало удар тока в темном отсеке, который приводил к увеличению латентного периода (ЛП) повторного захода в темный отсек, что рассматривается как критерий обучения. Тестирование проводили через 24 ч., 48 ч., 1 и 2 недели.
При тестировании через 24 ч. ЛП захода в темный отсек увеличился во всех группах животных (табл. 4) при этом статистически значимых отличий между животными, получавшими тактивин, физиологический раствор и диазепам не обнаруживалось.
| Таблица 4. | Латентный период захода в темный отсек (с) | |||||
| Группы животных | До УРПИ | Через 24 ч. | Через 48 ч. | Через 1 неделю | Через 2 недели | |
| Контроль | 5,4±0,8 | 130,2±3,5# | 96,3±4,2 | 96,1±4,5 | 95,0±5,2 | |
| Тактивин | 3,8±0,6 | 133,4±3,1# | 103,9±3,5 | 107,8±3,3** | 106,9±3,2** | |
| Пирацетам | 2,8±0,3 | 180,0#** | 180,0** | 180,0** | 180,0** | |
| Диазепам | 6,9±1,3 | 126,9±2,8# | 100,0±3,3 | 98,3±4,1 | 98,7±3,3 | |
#р<0,001 относительно значений до УРПИ; **р<0,001 относительно контроля.
Однако под действием тактивина наблюдался отсроченный эффект на память. Через неделю ЛП захода животного в темный отсек на фоне полипептидов тимуса статистически значимо (р<0,001) превышал контрольный уровень и этот эффект сохранялся через две недели (табл. 4). Животные, получавшие пирацетам при тестировании через 24 ч., 48 ч., 1 и 2 недели в темную камеру не входили, что отражалось в максимально большом значении ЛП. У животных, получавших диазепам, ЛП перехода в темный отсек не отличался от ЛП контрольных животных на протяжении всего опыта.
Следовательно, тактивин, в отличие от диазепама и подобно пирацетаму, оказал положительное влияние на память. Это позволяет заключить, что полипептиды тимуса обладают ноотропоподобными свойствами.
Влияние тактивина на выработку модифицированного УРПИ. Использование классического УРПИ не дает возможность ответить на вопрос избегает ли крыса темноты, связанной с электроболевым стимулом, или конкретного отсека, где этот стимул был применен. Поэтому нами использовалась модифицированная методика УРПИ. До обучения ЛП захода в темный отсек составлял в контроле – 5,4±0,8 с; на фоне тактивина – 3,8±0,6 с; пирацетама – 2,8±0,3 с; диазепама – 6,9±1,3 с. Через 24 ч. ЛП захода в опасный отсек в 3-камерной модифицированной установке статистически значимо возрастал (в контроле – до 48,0±4,1 с; на фоне тактивина – 149,0±2,6 с; на фоне пирацетама – 300,0±0,0 с; на фоне диазепама – 15±3,8 с; р<0,05), что с классической точки зрения свидетельствует об обучении животных. Однако как показывает анализ распределения возможных реакций контрольных животных, это не привело к предпочтению у животных заходов в безопасный отсек (табл. 5) ни через 24 ч., ни в более поздних тестированиях
| Таблица 5. | Выбор животными отсека в тесте условного рефлекса пассивного избегания в модифицированной установке |
| Время тестирования | Препарат | Число животных, первоначально выбравших | ||
| Темный опасный отсек | Светлый отсек | Темный безопасный отсек | ||
| 24 ч. | Контроль | 8 | 0 | 2** |
| Тактивин | 2 | 1 | 7** | |
| Пирацетам | 0 | 2 | 8** | |
| Диазепам | 5 | 1 | 4 | |
| 3 дня | Контроль | 8 | 1 | 1** |
| Тактивин | 5 | 1 | 4 | |
| Пирацетам | 0 | 3 | 7** | |
| Диазепам | 6 | 0 | 4 | |
| 1 неделя | Контроль | 6 | 0 | 4 |
| Тактивин | 7 | 1 | 2** | |
| Пирацетам | 1 | 3 | 6** | |
| Диазепам | 5 | 1 | 4 | |
| 2 недели | Контроль | 8 | 1 | 1** |
| Тактивин | 4 | 2 | 4 | |
| Пирацетам | 2 | 2 | 6* | |
| Диазепам | 5 | 1 | 4 | |
р<0,05, ** р<0,001, относительно выбора темного опасного отсека. Данные статистически обработаны с использованием критерия V согласования частот (Кулаичев А.П., 1999).
Следовательно, воздействие тока привело лишь к формированию условно-эмоциональной реакции страха. Таким образом, увеличение ЛП у контрольных животных не означает формирование памятного следа, отражающего пространственные характеристики. На фоне диазепама также не наблюдалось предпочтения безопасного отсека.
Тактивин, так же как пирацетам, приводил к предпочтению безопасных отсеков через 24 ч. В остальные дни эксперимента предпочтения темного безопасного отсека на фоне действия тактивина не наблюдалось (табл. 5). Это свидетельствует о мнемотропном влиянии полипептидов тимуса на пространственный компонент памяти на ранних стадиях формирования УРПИ.
Влияние тактивина на выработку условного рефлекса активного избегания (УРАИ). УРАИ вырабатывали у крыс в челночной камере. К условному раздражителю через 10 с изолированного действия подключался безусловный раздражитель – электрический ток. При переходе животного через отверстие в другую половину камеры во время действия тока звук и ток выключались. Если животное переходило во время следования только звукового сигнала, тока не следовало, а звук прекращался. Опыты проводили ежедневно в течение 14 дней до формирования устойчивого рефлекса.
Наши данные свидетельствуют о том, что под влиянием тактивина и пирацетама выработка УРАИ происходит быстрее, чем у контрольных животных на всем протяжении опыта, хотя число реакций избегания статистически значимо превышало контрольный уровень не во все дни. Диазепам угнетал выработку реакции избегания с 3 дня обучения (рис. 4).
Следовательно, в условиях данной модели также было выявлено мнемотропное влияние тактивина, сопоставимое с влиянием классического ноотропного препарата пирацетама. По нашим данным аналогичное влияние тактивина было обнаружено при изучении формирования пищевого условного рефлекса у крыс (Новоселецкая А.В., Киселева Н.М. и др., 2010).

Рисунок 4. Динамика формирования условного рефлекса активного избегания (УРАИ).
*р < 0,05 относительно контроля.
Таким образом, было выявлено стимулирующее действие тактивина на выработку условных рефлексов как с положительным, так и отрицательным подкреплением. Наиболее выраженное действие препарата наблюдалось в начале обучения.
Известно, что обучение во всех случаях сопровождается эмоциональным стрессом, его уменьшение до известных пределов приводит к улучшению процесса обучения.Положительный эффект препарата на ранних стадиях обучения, возможно, объясняется способностью препарата снижать эмоциональное напряжение животных, т.е. с его стресспротекторным эффектом.
Влияние тактивина на эмоциональный стресс
Следующим этапом нашей работы стало изучение влияния исследуемого препарата на эмоциональный стресс, вызываемый обратимыми функциональными нарушениями реакции избегания: сбоем и переменой местоположения отверстия (Иноземцев А.Н., Прагина Л.Л., 1992; Иноземцев А.Н. и др., 1996), которые являются моделями, рекомендованными фармакологическим комитетом РФ для расширенного изучения ноотропных и анксиолитических средств (Воронина Т.А., Островская Р.У., 2005; Воронина Т.А., Середенин С.Б., 2005).
Влияние тактивина на сбой УРАИ. В проведенных экспериментах показано, что сбой УРАИ вызывал глубокое нарушение выработанного навыка у контрольных животных. Число УРАИ в первом блоке предъявлений после функционального нарушения (ФН) уменьшалось в 3,3 раза относительно последних пяти предъявлений до сбоя (табл. 6).
| Таблица 6. | Влияние тактивина на воспроизведение условной реакции активного избегания после обратимых функциональных нарушений (ФН) |
| ФН | Вещество, кол-во животных | Реакции избегания, % | ||||
| Блоки предъявлений | ||||||
| До ФН | После ФН | |||||
| 16-20 | 1-5 | 6-10 | 11-15 | 16-20 | ||
| ФН – сбой УРАИ | Контроль n=50 | 92,0±7,3 | 28,0±7,3# | 76,0±6,3# | 76,0±4,3# | 80,0± 7,0 |
| Тактивин n=50 | 99,8±0,2 | 99,8±0,2* | 96,0±5,1* | 88,8±4,6* | 87,9±4,8 | |
| Пирацетам n=30 | 99,8±0,2 | 80,0±3,3#* | 73,3±4,8 | 93,3±3,9* | 99,8±0,2* | |
| Диазепам n=30 | 90,0±6,7 | 85,0±7,7#* | 80,0±6,8# | 70,0±5,4 | 90,0±5,8 | |
| ФН – ПМПО | Контроль, n=55 | 86,1±4,1 | 32,0±8,5# | 44,0±9,7# | 60,0±4,3 | 48,0±9,3# |
| Тактивин n=50 | 99,8±0,2 | 49,0±3,1#* | 74,2±7,3* | 77,5±4,1* | 79,2±4,7* | |
| Пирацетам n=30 | 99,8±0,2 | 43,8±2,8#* | 60,0±6,4#* | 93,3±3,7* | 96,1±3,5* | |
| Диазепам n=30 | 99,8±0,2 | 35,0±4,6# | 51,0±8,6 # | 55,0±5,8# | 58,0±5,2# | |
*р<0,05 относительно контроля; #р<0,05 относительно средней величины до ФН.
Тактивин полностью предупредил развитие сбоя, и величина УРАИ в первом блоке предъявлений не отличалась от исходной. Положительное влияние препарата на воспроизведение реакции избегания после сбоя проявилось также в том, что в первых трех блоках величина УРАИ была статистически значимо больше, чем у контрольных животных. Отмеченное влияние тактивина было выше, чем у эталонных препаратов пирацетама и диазепама.
Помимо уменьшения реакций избегания наблюдалось возрастание межсигнальных реакций (МСР), что свидетельствует о возникновении эмоционального стресса (Симонов П.В., 1981). Анксиогенный характер указанных реакций подтверждается в наших опытах их уменьшением на фоне диазепама и других транквилизаторов (Иноземцев А.Н. и др., 1996). На фоне тактивина и пирацетама мы также наблюдали уменьшение относительно контроля МСР и быстрое восстановление их уровня до исходных величин (рис. 5).

Рисунок 5. Межсигнальные реакции до и после сбоя реакции избегания.
*-р<0,05 относительно контроля; #-р<0,05 относительно средней величины до ФН
Перемена местоположения отверстия (ПМПО) приводила к более глубокому по сравнению со сбоем нарушению выработанного навыка у контрольных животных, поскольку исходный уровень УРАИ не достигался к концу опыта (табл. 6).
Под действием тактивина и пирацетама нарушение было менее выражено, и уровень избегания был выше контрольного (р<0,05). Диазепам не оказывал положительного влияния на воспроизведение реакции избегания в условиях данной модели. После ПМПО одновременно со снижением уровня избегания наблюдался рост генерализованной двигательной активности животных, который выражался в возрастании числа МСР (рис. 6). Пространственная переделка установки вызывала более значимое увеличение МСР, чем сбой УРАИ, что подтверждает более сильное стрессогенное влияние данного ФН. Рост числа МСР в первом блоке предъявлений после ПМПО был значимо менее выражен на фоне тактивина и пирацетама.

Рисунок 6. Влияние препаратов на межсигнальные реакции до и после ПМПО. *р<0,05 относительно контроля; #р<0,05 относительно средней величины до ФН.
Результаты этого раздела демонстрируют, что полипептидный препарат тимуса тактивин уменьшал нарушение выработанного навыка при эмоциональном стрессе, вызванным обратимыми функциональными нарушениями реакции избегания. Указанный эффект был аналогичен действию ноотропного препарата и отличался от действия анксиолитика диазепама.
Изучение механизма стресс-протекторного действия тактивина
Влияние тактивина на активность ГАМК-ергической системы крыс после сбоя УРАИ. В связи с полученными данными встает вопрос о механизмах стресспротекторного действия тактивина. На первом этапе мы исследовали его влияние на работу ГАМК-ергической системы – одной из ведущих стресс-протекторных систем. Как показали исследования тактивин не вызвал увеличения экспрессии ГАМКА-рецепторов и не изменял сродство данных рецептов к лиганду в гипоталамусе и коре интактных животных (табл. 7). На основании полученных данных можно заключить, что фармакологический эффект тактивина не зависит от состояния ГАМКА рецепторов в структурах мозга интактных животных.
| Таблица 7. | Характеристики радиолигандного связывания ГАМКА-рецепторов структур мозга крыс Вистар | ||||
| Контроль | Тактивин | ||||
| Bmax (фмоль/мг белка) | Kd (нмоль) | Bmax (фмоль/мг белка) | Kd (нмоль) | ||
| Кора | 1059±71 | 8±1 | 1168±65 | 8±1 | |
| Гипоталамус | 2000±196 | 15±3 | 2305±173 | 12±2 | |
Bmax – количество мест связывания лиганда, Кd – степень сродства рецептора к лиганду.
Кроме того, было проведено исследование уровня ГАМК в ряде структур головного мозга крыс на фоне эмоционалного стресса, вызванного сбоем реакции избегания. Как видно из данных, представленных в таблице 8, введение тактивина животным, подвергшимся воздействию эмоционального стресса, не повлияло на уровень ГАМК в исследованных структурах головного мозга крыс.
Исходя из полученных данных, можно предполагать, что тактивин не влияет на активность ГАМК-ергической системы как у интактных, так и у стрессированных животных.
| Таблица 8. | Уровень ГАМК (нмоль/мг ткани) в структурах головного мозга крыс Вистар на фоне эмоционального стресса |
| Структура | Содержание ГАМК в структурах мозга в группах животных | Уровень значимости различия результатов (р контр.-опыт) | |
| Контроль | На фоне тактивина | ||
| Гипоталамус | 0,844 ± 0,057 | 0,813 ± 0,056 | 0,70 |
| Фронтальная кора | 0,415 ± 0,009 | 0,406 ± 0,004 | 0,42 |
| Прилежащее ядро | 1,032 ± 0,059 | 0,988 ± 0,048 | 0,59 |
| Стриатум | 0,549 ± 0,017 | 0,538 ± 0,026 | 0,75 |
| Гиппокамп | 0,451 ± 0,017 | 0,442 ± 0,019 | 0,73 |
Влияние тактивина на уровень моноаминов в мозге крыс после сбоя УРАИ. Тактивин вызывал достоверные изменения в распределении моноаминов в большинстве изучаемых структур головного мозга у интактных крыс. Во фронтальной коре на фоне действия препарата статистически значимо снижался уровень дофамина (ДА), следствием чего являлось увеличение соотношения его метаболитов (дигидроксифенилуксусной и гомованиловой кислот) к самому медиатору. В гиппокампе статистически значимо увеличивался уровень норадреналина (НА), а в стриатуме – ДА. В гипоталамусе и прилежащем ядре уровень медиаторов, метаболитов и их соотношение у опытной группы не отличались от контроля (табл. 9).
Это позволяет заключить, что увеличение двигательной и исследовательской активности на фоне тактивина практически во всех тестах, вероятно, связано с повышением ДА в стриатуме.
После эмоционального стресса на фоне тактивина наблюдалось статистически значимое увеличение серотонина (5-ГТ) и НА во фронтальной коре, гипоталамусе и стриатуме (табл. 9). Так как указанные изменения не наблюдались у интактных крыс (без обучения и сбоя), то можно заключить, что наблюдаемые изменения в содержании моноаминов вызваны эмоциональным стрессом.
Увеличение синтеза и содержания серотонина в структурах головного мозга при стресс-реакциях отмечалось многими авторами (Горбунова А.В., 1998; Clement H.W., et al. 1998; Sammers C.H., et al. 1998). Такое увеличение медиатора рассматривают как вариант адаптации к стресс-воздействию. В частности показано увеличение 5-ГТ в условиях нашей модели (Литвинова С.В. и др., 2007).
На основе нейрохимического анализа корковых и стволовых структур мозга показано, что действие тактивина на фоне эмоционального стресса проявилось в изменении баланса серотонин/норадреналин в пользу первого в гипоталамусе и фронтальной коре крыс, что позволяет объяснить предотвращение функционального нарушения памяти, вызванного сбоем реакции избегания.
| Таблица 9. | Влияние тактивина на содержание моноаминов и их метаболитов в структурах мозга крыс до и после сбоя реакции избегания (% от контроля) |
| НА | ДА | ДОФУК | ГВК | ДОФУК /ДА | ГВК /ДА | 5-ОТ | 5-ГИУК | 5-ГИУК /5-ОТ | |
| фронтальная кора | |||||||||
| До сбоя | 92,9 ±4,1 | 62,3 ±7,3* | 99,4 ±8,9 | 106,5 ±4.6 | 159,1 ±9,5* | 148,7 ±5,3* | 88,0 ±8,1 | 91,3 ±7,7 | 100,7 ± 4,4 |
| После сбоя | 151,3 ± 3,4* | 62,5 ±6,8 | 137,2 ±18,7 | 146,3±26,4 | 214,3 ±16,7* | 238,9±9,6* | 137,5 ±8,6* | 289,9 ±17,1* | 215,2 ±13,8* |
| гипоталамус | |||||||||
| До сбоя | 95,3 ±3,1 | 85,9 ±8,7 | 90,1 ±10,2 | 73,9 ±10,3 | 101,8 ±5,1 | 80,2 ±16,7 | 96,8 ±7,6 | 105,8 ±6,9 | 107,5 ±7,7 |
| После сбоя | 115,8 ±4,0* | 122,6 ±5,0* | 103,2 ±6,7 | 108,1 ±9,4 | 79,1 ±0,9* | 89,3 ±9,8 | 167,1 ±5,7* | 95,9 ±9,8 | 93,5 ±10,6 |
| гиппокамп | |||||||||
| До сбоя | 133,6 ±9,7* | 96,8 ±6,5 | 81,9 ±10,1 | 113,3 ±5,9 | 87,8 ±11,5 | 135,6 ±20,4 | 97,5 ±5,7 | 100,6 ±3,1 | 103,9 ±4,8 |
| После сбоя | 102,2 ±5,6 | 25,1 ±3,7* | 91,4 ±7,9 | 107,9 ±7,1 | 196,5 ±6,8* | 18,5 ±6,2* | 109,4 ±14,1 | 88,4 ±3,4* | 193,6 ±4,5* |
| прилежащее ядро | |||||||||
| До сбоя | 160,9 ±38,4 | 86,9 ±12,6 | 114,1 ±10,3 | 95,2 ±4,6 | 125,4 ±18,6 | 102,9 ±7,3 | 90,4 ±7,5 | 98,3 ±5,8 | 108,1 ±8,9 |
| После сбоя | 96,6 ±8,2 | 102,1 ±3,5 | 236,4 ±18,7* | 91,9 ± ±17,4 | 257,7 ±13,7* | 95,3 ± 6,8 | 112,6 ±9,3 | 102,6 ±5,7 | 26,9 ±4,5* |
| стриатум | |||||||||
| До сбоя | 91,1 ±9,1 | 112,5 ±1,3* | 109,9 ±5,3 | 114 ±8,9 | 96,8 ±7,5 | 100,8 ±3,5 | 123,7 ±16,6 | 125,2 ±17,8 | 100,3 ±3,2 |
| После сбоя | 158,6 ±6,8* | 99,9 ±3,7 | 103,6 ±2,5 | 90,8 ±8,3 | 111,7 ±10,6 | 82,1 ±12,9 | 143,6 ±6,6* | 229,4 ±8,0* | 96,7 ± 5,1 |
Примечание: Приведены средние значения и стандартные ошибки среднего «m±S.E.M». *р<0,05 в сравнении с контролем (U-критерий Манна-Уитни). / р<0,05 в сравнении со значениями полученными до обучения и сбоя.
Сокращения: НА – норадреналин; ДА – дофамин; ДОФУК – дигидроксифенилуксусная кислота; ГВК – гомованиловая кислота; 5-ГТ – серотонин; 5-ГИУК – 5-гидроксииндолуксусная кислота.
Как показано в экспериментах, применение поведенческих тестов, а также выработка условных рефлексов и их функциональных нарушений позволили нам выявить влияние полипептидов тимуса на ЦНС, выражающееся в мнемотропном и стресспротекторном действии. Нейрохимический анализ различных структур мозга крыс после эмоционального стресса подтвердил антистрессорные свойства тактивина по количественной оценке баланса моноаминов в различных структурах мозга, вовлеченных в организацию поведения. Следовательно, полипептиды тимуса участвуют не только в реализации работы иммунной системы, но и активно регулируют работу стресслимитирующей системы.
Таким образом, при стрессе активизируется не только гипоталам-гипофизарно-надпочечниковая ось, но и гипатоламо-гипофизарно-тимусная система, которая ограничивает стресс-повреждающий эффект чрезмерной активности гипоталамо-гипофизарно-надпочечниковой оси.
ВЫВОДЫ
- Сывороточная тимическая активность в крови здоровых рожениц с неотягощенным анамнезом и неосложненным течением беременности выше, чем у небеременных женщин детородного возраста, что является адекватным ответом организма на родовой стресс.
- Уровень СТА рожениц с осложненным течением беременности и/или на фоне ремиссии хронических заболеваний был ниже, чем у здоровых рожениц. Это может косвенно свидетельствовать об истощении функции тимуса у данной категории рожениц.
- В пуповинной крови доношенных новорожденных детей (38-40 недель гестации) уровень СТА не зависит от состояния здоровья и уровня СТА их матерей, что свидетельствует о самостоятельном функционировании тимуса у новорожденных.
- Полипептидный препарат тимуса тактивин увеличивает у крыс уровень исследовательской и двигательной активности в «открытом поле», норковой и челночной камерах. Данный эффект, сопоставимый с действием классического ноотропного препарата пирацетама и эталонного анксиолитика диазепама, свидетельствует о влиянии пептидов тимуса на функции ЦНС.
- Препарат полипептидов тимуса тактивина оказывает активирующее влияние на выработку у крыс условных рефлексов, что наиболее ярко проявляется на начальных этапах формирования памятного следа. Мнемотропный эффект препарата аналогичен действию классического ноотропного препарата пирацетама.
- Тактивин оказывает сопоставимое с пирацетамом положительное влияние на пространственный компонент памяти при тестировании условной реакции пассивного избегания в трехкамерной установке и при обратимом функциональном нарушении реакции избегания - перемене местоположения отверстия.
- Препарат тимуса на фоне эмоционального стресса, вызванного обратимыми функциональными нарушениями – сбоем и пространственной переделкой навыка улучшает воспроизведение реакции избегания. Эффект полипептидов тимуса сопоставим с влиянием ноотропного препарата пирацетама и отличен от эффекта анксиолитика диазепама, улучшавшего воспроизведение реакции избегания только при сбое.
- Действие тактивина на фоне эмоционального стресса проявляется в изменении баланса уровня серотонин/норадреналин в пользу первого в гипоталамусе и фронтальной коре крыс. Изменений в ГАМК-ергической системе выявлено не было.
- На фоне действия тактивина статистически значимо увеличивается уровень дофамина в стриатуме у интактных животных, что объясняет увеличение двигательной и исследовательской активности на фоне препарата практически во всех поведенческих тестах.
- Совокупность полученных данных позволяет полагать, что при стрессе активизируется гипатоламо-гипофизарно-тимусная система и повышается уровень полипептидов тимуса, который ограничивает стресс-повреждающий эффект чрезмерной активности гипоталамо-гипофизарно-надпочечниковой оси.
ПРАКТИЧЕСКИЕ РЕКОМЕНДАЦИИ
- Учитывая факт мнемотропного действия полипептидного препарата тимуса тактивина (разрешенного к применению в клинической практике как иммунотропного препарата), рекомендовать его для дальнейших исследований в клинической практике для коррекции различных когнитивных функций и мнестико-интеллектуальных расстройств.
- Рекомендовать тактивин для дальнейшего изучения профилактического стресспротекторного действия у человека, подвергающегося выраженным стрессогенным воздействиям, поскольку препарат предупреждает развитие негативных последствий эмоционального стресса у животных.
СПИСОК ОПУБЛИКОВАННЫХ РАБОТ ПО ТЕМЕ ДИССЕРТАЦИИ
- Киселева, Н.М. Опиоидные пептиды и гормонально-обменный статус у детей раннего возраста в периоде клинического благополучия и на фоне ОРВИ по данным корреляционного анализа / Т.И. Назарова, Кузьменко Л.Г., Н.М. Киселева и др. // Вестник Российского Университета Дружбы Народов. Серия терапия. – 1995. – №1. – С. 81-84.
- Киселева, Н.М. Сывороточная тимическая активность пуповинной крови доношенных новорожденных детей / Н. Исмаиль Насер, Л.Г. Кузьменко, Н.М. Киселева и др. // Материалы IV-ой Международной научно-практической конференции «Здоровье и Образование в XXI веке. – М.: РУДН, 2003. – С. 273-274.
- Киселева, Н.М. Структурные изменения тимуса и иммунный статус в динамике осложненного течения острых респираторных заболеваний у детей раннего возраста / Л.Г. Кузьменко, Н.А. Тюрин, Н.М. Киселева и др. // Детские инфекции. – 2005. – Том 4, №1. – С.25-29.
- Киселева, Н.М. Отдаленный эффект препаратов тимуса у детей с рецидивирующими заболеваниями органов дыхания / Л.Г. Кузьменко, Н.А. Лопушанская, Н.М. Киселева // Материалы Российского респираторного общества 15-го Национального конгресса по болезням органов дыхания, 1-го Евроазиатского респираторного общества. – М. – 2005. – С.309.
- Киселева, Н.М. Ближайшие и отдаленные результаты лечения препаратами тимуса детей с различными заболеваниями / Л.Г. Кузьменко, Н.А. Лопушанская, Н.М. Киселева и др. // Детские инфекции. – 2005. – Том 4, №4. – С.42-45.
- Киселева, Н.М. Влияние тимуса экстракт на показатели Ig E при синдроме бронхиальной обструкции на фоне частых ОРВИ у детей раннего возраста / Л.Г. Кузьменко, В.К. Котлуков, Н.М. Киселева и др. // Актуальные проблемы педиатрии (сборник материалов XI конгресса педиатров России 5—8 февраля 2007). – М., ГЭОТАР-Медиа. – 2007. – С.359.
- Киселева, Н.М. Результаты лечения детей тимуса экстрактом / Л.Г. Кузьменко, Л.Я. Пушко, Н.М. Киселева и др. // Актуальные проблемы педиатрии (сборник материалов XI конгресса педиатров России 5—8 февраля 2007). – М., ГЭОТАР-Медиа. – 2007. – С.360.
- Киселева, Н.М. Влияние препаратов экстрактов тимуса на адаптацию детей со сниженной функцией Т-звена иммунной системы / Н.М. Киселева, Л.Г. Кузьменко, А.Н. Иноземцев // Материалы международного симпозиума «Адаптационная физиология и качество жизни: проблемы традиционной и инновационной медицины», посвящённого 80-летию академика РАМН Н.А. Агаджаняна 14—16 мая 2008 г. – М.,РУДН. – 2008. – С. 145-147.
- Киселева, Н.М. Уровень сывороточной тимической активности в пуповинной крови у доношенных новорожденных детей / В.Я. Арион, Л.Г. Кузьменко, Н.М. Киселева // Российский иммунологический журнал. – 2008. – Т.2 (11), № 2—3. – С.326.
- Киселева, Н.М. Тимус и его роль в работе стресс-лимитирующей системы / Киселева Н.М., Иноземцев А.Н. // Материалы VI Сибирского физиологического съезда 25—27 июня 2008 г. – Барнаул. – Т.2. – С.69-70.
- Киселева, Н.М. Значение иммунокорегирующего препарата тактивина при функциональном нарушении реакции избегания у крыс / Н.М. Киселева, А.В. Новоселецкая, И.В. Зимина и др. // Аллергология и иммунология. – 2008. –Т.9, № 3. – С.269.
- Киселева, Н.М. Тактивин в лечении детей с тимомегалией / Л.Г. Кузьменко, О.В. Быстрова, Н.М. Киселева и др. // Аллергология и иммунология. – 2008. – Т.9, № 3. – С.351-352.
- Киселева, Н.М. Интоксикационный стресс и иммунокоррекция / В.Я. Арион, И.В. Зимина, Н.М. Киселева и др. // Аллергология и иммунология. – 2008. – Т.9, № 4. – С.438-440.
- Киселева, Н.М. Показатели уровня сывороточной тимической активности пуповинной крови новорожденных детей / Л.Г. Кузьменко, В.Я. Арион, Н.М. Киселева и др. // Иммунофизиология: естественный аутоиммунитет в норме и патологии/Под.ред. Полетаева А.Б., Данилова А.Н. – М. – 2008. – С.114-116.
- Киселева, Н.М. Влияние тактивина на функциональное нарушение реакции избегания у крыс / Н.М. Киселева, А.В. Новоселецкая, И.В. Зимина и др. // Бюлл. экспер. биол. и мед. – 2009. – Т.147, №1. – С. 75-77.
- Киселева, Н.М. Влияние иммунокоррегирующего препарата на поведенческие реакции крыс (экспериментальное исследование) / Н.М. Киселева, А.В. Новоселецкая, И.В. Зимина и др. // Аллергология и иммунология. – 2009. –Т.10, № 1. – С.151-152.
- Киселева, Н.М. Влияние некоторых иммунотропных препаратов на когнитивные функции у крыс / А.В. Новоселецкая, Н.М. Киселева, А.Н. Иноземцев // Материалы ХVI Российского национального конгресса «Человек и лекарство». – М., 2009. – С.551.
- Киселева, Н.М. Этологический анализ поведения крыс на фоне тактивина / Н.М. Киселева, А.В. Новоселецкая, А.В. Крючкова и др. // Материалы Российского национального конгресса «Человек и лекарство». – М., 2009. – С.673.
- Киселева, Н.М. Влияние иммунотропных препаратов на процессы обучения и памяти / Н.М. Киселева, А.В. Новоселецкая // Материалы V международный междисциплинарного конгресса «Нейронаука для медицины и психологии». – М.:МАКС Пресс. – 2009. – С.122.
- Киселева, Н.М. Тимус и стресс-лимитирующая система / Н.М. Киселева, В.Я. Арион, И.В. Зимина и др. // Аллергология и иммунология. – 2009. – Том 10, №3. – С. 365-367.
- Киселева, Н.М. Влияние иммуномодулятора тактивина на число ядрышек в однородных популяциях нейронов трех структур головного мозга крыс / А.В. Крючкова, А.В. Новоселецкая, Н.М. Киселева и др. // International Journal on Immunorehabilitation. Международный журнал по иммунореабилитации. – 2009. – Том11, №1. – С.30.
- Киселева, Н.М. Воздействие полипептидов тимуса на работу стресс-системы / Н.М. Киселева // Материалы Х Международного конгресса «Здоровье и образование в XXI Веке» «Инновационные технологии в биологии и медицине». – М., РУДН. – 2009. –С.449-452.
- Киселева, Н.М. Влияние экстракта тимуса тактивина на формирование пищевого условного рефлекса у крыс / А.В. Новоселецкая, Н.М. Киселева, А.Н. Иноземцев и др. // Российский иммунологический журнал. – 2010. – Том 4 (13), №1. – С. 83-87.
- Киселева, Н.М. Влияние тактивина на поведение и обучение крыс / Н.М. Киселева, А.В. Новоселецкая, И.В. Зимина и др. // Вестник Российской Академии медицинских наук. – 2010. – №1. – С. 23-26.
- Киселева, Н.М. Анализ поведения крыс в приподнятом крестообразном лабиринте на фоне иммуно- и нейротропных средств / А.В. Новоселецкая, Н.М. Киселева, А.Н. Иноземцев // Материалы конгресса «Человек и лекарство». – М., 2010. – С. 691-692.
- Kiseleva, N.M. Effects of immunotropic drugs on passive avoidance response in rats / A.V. Novoseletskaya, N.M. Kiseleva, A.N. Inozemtsev // Proceedings 5th International Conference “Biological Basis of Individual Sensitivity to Psychotropic Drugs’. – M.: Folium, 2010. – P.55-56.
- Киселева, Н.М. Стресспротекторная роль тактивина / Н.М. Киселева, А.В. Новоселецкая, А.В. Крючкова и др. // Материалы XXI Съезда Физиологического общества им. И.П. Павлова. – М.-Калуга: Типография ООО «БЭСТ-принт», 2010. –С. 272.
- Киселева, Н.М. Изучение мнемотропного эффекта иммунотропных препаратов / А.В. Новоселецкая, Н.М. Киселева, А.Н. Иноземцев // Материалы XXI Съезда Физиологического общества им. И.П. Павлова. – М.-Калуга: Типография ООО «БЭСТ-принт», 2010. – С. 443-444.
- Киселева, Н.М. Возможная роль тимуса в работе стресс-лимитирующей системы / Н.М. Киселева, А.Н. Иноземцев // Иммунопатология, аллергология, инфектология. – 2010. –№3. – С. 13-20.
- Киселева, Н.М. Иммуномодулятор тактивин: новый спектр действия / Н.М. Киселева, А.В. Новоселецкая, А.Н. Иноземцев // Материалы ХI Международного конгресса «Здоровье и образование в ХХI веке» «Научные и прикладные аспекты концепции здоровья и здорового образа жизни». – М.- 2010.- С. 390.
- Киселева, Н.М. Условно-рефлекторная память под воздействием иммунотропных препаратов / А.В. Новоселецкая, Н.М. Киселева, А.Н. Иноземцев // Механизмы регуляции физиологических систем организма в процессе адаптации к условиям среды: Материалы Всероссийской конференции с международным участием, посвященная 85-летию со дня основания Института физиологии им. И.П. Павлова РАН. – С-Пб. – 2010. – С. 215 – 216.
- Киселева, Н.М. Функциональная активность тимуса в раннем онтогенезе человека / О.В. Быстрова, Л.Г. Кузьменко, Н.М. Киселева и др. // Материалы XV Конгресса педиатров России с международным участием «Актуальные проблемы педиатрии». – М., 2011. – С.113.
- Киселева, Н.М. Функциональная активность тимуса в парах мать-новорожденный / Н.М. Киселева, О.В. Быстрова, Л.Г. Кузьменко и др. // Материалы XV Конгресса педиатров России с международным участием «Актуальные проблемы педиатрии». – М., 2011. – С. 408.
- Киселева, Н.М. Функциональное состояние тимуса у доношенных новорожденных детей и их матерей / Л.Г. Кузьменко, О.В. Быстрова, И.В. Зимина и др. // Ж. Педиатрия им. Г.Н. Сперанского. – 2011. – Т.90, №1. – С. 8-13.
- Киселева, Н.М. Влияние иммунокорригирующего препарата (тактивина) на активность некоторых нейромедиаторных систем в структурах головного мозга крыс / Н.М. Киселева, А.В. Новоселецкая, А.В. Крючкова и др. // Аллергология и иммунология. – 2011. – Том. 12, № 1. – С. 44.
- Киселева, Н.М. Биодоступность иммунотропного препарата из тимуса тактивина, введенного неинвазиным методом / О.В. Быстрова, И.В. Зимина, Н.М. Киселева и др. // Аллергология и иммунология. – 2011. – Том. 12, № 1. – С. 41.
- Киселева, Н.М. Тактивин противодействует угнетению тяжелыми металлами обучения и памяти у крыс в условиях эмоционально-болевого стресса \ С.Б. Бокиева, А.В. Крючкова, Н.М. Киселева и др. // Фундаментальные и прикладные проблемы стресса: материалы II Международной научно-практической конференции, 21 апреля 2011 / Вит. гос. ун-т; редкол.: А.П.Солодков и др. – Витебск: УО «ВГУ им. П.М.Машерова», 2011. – С. 8-10.
- Киселева, Н.М. Стресс-протекторное действие тактивина / А.В. Новоселецкая, Н.М. Киселева, И.В. Зимина и др. // Фундаментальные и прикладные проблемы стресса: материалы II Международной научно-практической конференции, 21 апреля 2011/Вит. гос. ун-т; редкол.: А.П.Солодков (и др.). – Витебск: УО «ВГУ им. П.М.Машерова», 2011. – С. 231-233.
- Киселева, Н.М. Регуляторная роль тимуса в поддержании гомеостаза / Н.М. Киселева, А.В. Новоселецкая, И.В. Зимина и др. // Аллергология и иммунология. – 2011. – Том 12, № 3. – С. 237-239.
- Киселева, Н.М. Мнемотропные свойства иммунокоррегирующего препарата тимуса тактивина / А.В. Новоселецкая, Н.М. Киселева, А.Н. Иноземцев и др. // Аллергология и иммунология. – 2011. – Том 12, № 3. – С. 255-257.
- Киселева, Н.М. Тимус и стресс в клинике и эксперименте / Л.Г. Кузьменко, Н.М. Киселева // Вестник РУДН, серия Медицина. – 2011. – № 3. – С. 89-94.
- Киселева, Н.М. Иммуномодуляторы деринат и тактивин изменяют баланс моноаминов в мозге крыс после эмоционального стресса / А.В. Новоселецкая, Н.М. Киселева, А.Н. Иноземцев и др. // Материалы III Съезда физиологов СНГ / под редедакцией А.И. Григорьева, О.А. Крышталя, Ю.В. Наточина и др. – М.: Медицина – Здоровье, 2011. – С. 176 -177.
- Киселева, Н.М. Тимус и эндокринная система / Л.Г. Кузьменко, Н.М. Киселева // Материалы V городской научно-практической конференции «Эндокринологические аспекты в педиатрии». – Москва – 2011. – С. 25.
- Киселева, Н.М. Иммунотропный препарат тактивин противодействует нейротоксическому влиянию тяжелых металлов на обучение и память крыс / А.Н. Иноземцев, С.Б. Бокиева, Н.М. Киселева и др. // Российский иммунологический журнал. - 2011. – Т. 5(14), № 3–4. – С.274-278.
- Киселева, Н.М. Клиническая смерть как четвертая стадия астматического статуса у детей / Л.Г. Кузьменко, В.А. Бычков, Н.М. Киселева и др. // Материалы XII международного конгресса «Здоровье и образование в ХХI веке». – Россия-Москва, 2011. – С. 539-542.
- Киселева, Н.М. Критерии оценки уровня лимфоцитов и стресс / Л.Г. Кузьменко, Н.М. Киселева, М.М. Нкане Нкоза // Материалы XII международного конгресса «Здоровье и образование в ХХI веке». – Россия-Москва, 2011. – С. 360-361.
- Киселева, Н.М. Нейрохимическое изучение механизма действия полипептидного препарата тимуса (тактивина) при эмоциональном стрессе / Н.М. Киселева, А.В. Новоселецкая, А.Н. Иноземцев и др. // Вестник РУДН, серия Медицина.- 2011.- № 4. – С. 143-148.
- Киселева, Н.М. Стресс и лимфоциты / Н.М. Киселева, Л.Г. Кузьменко, М.М. Нкане Нкоза // Ж. Педиатрия им. Г.Н. Сперанского. – 2012. – Т. 91, № 1. – С. 137-143.
- Kiseleva, N.M. Regulatory role of thymus in maintenance of homeostasis / N.M. Kiseleva, A.V. Novoseletskaya, I.V. Zimina et al. //Allergy, Asthma & Immunophysiology: From Basic Science to Clinical Application/ Ed. R. Sepiashvili. – MEDIMOND s.r.I., 2012. – P. 217-223.
- Киселева, Н.М. Количественная оценка обучения и памятного следа при изучении мнемотропных свойств иммунотропных препаратов / Н.М. Киселева, А.В. Новоселецкая, Е.Б. Воеводина и др. // Бюлл. эксперимент. биолог. и мед. – 2012. – Т. 154, №8. – С. 197-201.
- Киселева, Н.М. Влияние препарата пептидов тимуса (тактивина) на уровень тревожности крыс / А.В. Новоселецкая, А.Н. Иноземцев, Н.М. Киселева и др. // Вестник РУДН. Серия медицина. – 2012. – №3. – С. 12-15.
- Киселева, Н.М. Влияние препарата пептидов тимуса тактивина на развитие стресс-вызванной аналгезии, вызванной иммобилизацией / Новоселецкая А.В., Матвеева О.Д., Киселева Н.М. и др. // Здоровье и образование в XXI веке. Серия Медицина– 2012. – № 1, Т.14. – С. 227- 228.
- Киселева, Н.М. Влияние препарата полипептидов тимуса (тактивина) на содержание возбуждающих и тормозных аминокислот в структурах мозга крыс / Новоселецкая А.Н., Киселева Н.М., Иноземцев А.Н. и др. // Российский Иммунологический журнал. – 2012. – Т. 6 (15), № 4. – С.395 -398.
УЧЕБНО-МЕТОДИЧЕСКИЕ ТРУДЫ
- Киселева, Н.М. Иммунокоррекция Тактивином в комплексном лечении часто болеющих детей (пособие для врачей) / Л.Г. Кузьменко, Ю.М. Лопухин, Н.М. Киселева и др. – М.; ГЭОТАР-Медиа. – 2005. – 48 с.
СПИСОК СОКРАЩЕНИЙ
ГАМК – гаммааминомасляная кислота
ГВК – гомованилиновая кислота
ГК – глюкокортикоиды
ДА – дофамин
ДОФУК – дигидроксифенилуксусная кислота
ЛП – латентный период
НА - норадреналин
МСР – межсигнальные реакции
ОП – опиоидные пептиды
ПКЛ – приподнятый крестообразный лабиринт
ПМПО – перемена местоположения отверстия
ПУР – пищевой условный рефлекс
СТА – сывороточная тимическая активность
УРАИ – условный рефлекс активного избегания
УРПИ– условный рефлекс пассивного избегания
мУРПИ– УРПИ в модифицированной установке
5 -ГИУК – 5-гидроксииндолуксусная кислота
5-ГТ – серотонин
РРО – 2,5-дифенилоксазол
РОРОР – 1,4-ди-[2-фенил-(5-окзазолил)]-бензол