

Кодирование признаков изображения и сложных зрительных образов нейронами коры головного мозга млекопитающих
На правах рукописи
Игорь Вечеславович БОНДАРЬ
КОДИРОВАНИЕ ПРИЗНАКОВ ИЗОБРАЖЕНИЯ И СЛОЖНЫХ ЗРИТЕЛЬНЫХ ОБРАЗОВ НЕЙРОНАМИ КОРЫ ГОЛОВНОГО МОЗГА МЛЕКОПИТАЮЩИХ
Автореферат
диссертации на соискание ученой степени
доктора биологических наук
03.03.01 - Физиология
Москва
2011
Работа выполнена в лаборатории физиологии сенсорных систем
(и.о. заб. лаб. доктор биологических наук Елена Семеновна Михайлова) Учреждения Российской академии наук Института высшей нервной деятельности и нейрофизиологии РАН
(директор – доктор биологических наук, профессор Павел Милославович Балабан)
Научный консультант: Академик РАН, доктор биологических наук
Игорь Александрович ШЕВЕЛЁВ
Официальные оппоненты: Доктор биологических наук
Галина Христофоровна МЕРЖАНОВА
Доктор биологических наук, профессор
Александр Васильевич ЛАТАНОВ
Доктор биологических наук, профессор
Александр Яковлевич СУПИН
Ведущая организация: Учреждение Российской академии наук
Институт физиологии им. И.П. Павлова РАН
Защита состоится 2011 года в 14:00 часов на заседании Диссертационного совета
Д-002.044.01 по защите докторских диссертаций при Учреждении Российской академии наук Институте высшей нервной деятельности и нейрофизиологии РАН по адресу:
117485, Москва, ул. Бутлерова, д. 5а.
Факс: (495) 3388500; e-mail: [email protected], [email protected]
С диссертацией можно ознакомиться в библиотеке Института высшей нервной деятельности и нейрофизиологии РАН
Автореферат разослан « ___ » ______________ 2010 года
Ученый секретарь Диссертационного совета
доктор биологических наук, профессор В.В. Раевский
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы
Обработка зрительной информации представляет собой сложный иерархически организованный процесс, цели которого – категоризация, классификация и выявление биологической значимости объектов окружающей среды. Основной единицей, занятой в анализе характеристических признаков зрительных объектов, является нейрон-детектор. Детекторный принцип работы мозга был сформулирован после того, как в 1959 г. МакКаллок и Питс с коллегами в сетчатке лягушки обнаружили нейроны, тонко настроенные на параметры сложных зрительных образов: размер, скорость движения, контраст и т.д. Позднее подобного рода клетки-детекторы были найдены на разных уровнях корковой системы анализа зрительной информации: детекторы ориентации линий в первичной зрительной коре кошки (Hubel & Wiesel, 1962); детекторы пересечений и ветвлений линий в стриарной коре кошки (Шевелев, 1993); нейроны-детекторы лиц в нижневисочной коре мозга макаки резус (Gross, 1972). Теоретические и практические исследования в области детекторной модели кодирования сенсорного сигнала были проведены Е. Н. Соколовым (Соколов, 1995).
Изучение свойств нейронов-детекторов тесно связано с теоретическими работами в области моделирования зрительной системы. Еще в 1982 Марр сформулировал формальные принципы, которые следует принимать в расчет при создании искусственных систем распознавания зрительных образов. Тем не менее, всеобъемлющая теория работы зрительного анализатора до сих пор не создана, а компьютерные системы распознавания образов показали свою функциональность лишь при определенных условиях зрительной среды. Именно поэтому так важны работы по изучению детекторных свойств нейронов, которые смогут объяснить принципы обработки информации в мозге животных. Понимание механизмов, лежащих в основе работы зрительного анализатора, позволит создать интеллектуальные системы распознавания зрительных образов, способные облегчить работу операторов как в сфере обеспечения безопасности жизнедеятельности, так и в разнообразных областях промышленности.
В процессе работы зрительный анализатор сталкивается и успешно решает проблемы, которые вызывают серьезные затруднения при создании искусственных систем распознавания зрительных образов. Так, даже незначительные вариации условий освещения зрительных объектов или изменение их размеров оказывают значительный негативный эффект на успешность распознавания объектов искусственным зрительным анализатором. В связи с этим крайне актуальны исследовательские работы, направленные на детальное изучение свойств нейронов-детекторов. Высокая эффективность нейронов-детекторов связана с их уникальной адаптивной способностью. При этом нервные клетки мозга могут не только приспосабливаться к быстро изменяющимся условиям среды, но и подстраиваться к особенностям зрительного окружения, с которым приходится длительное время контактировать живому организму. Другой важной отличительной чертой детекторов является их способность стабильно кодировать информацию о характеристических признаках изображения и зрительных объектах. И, наконец, однозначность кодирования обеспечивает высокую точность представления информации в мозге. Исследования последних лет значительно обогатили знания о принципах кодирования информации в зрительном анализаторе человека и животных. Тем не менее, изучение адаптивности, стабильности и однозначности кодирования занимают главенствующее положение в работе ведущих мировых лабораторий.
По существующим на настоящий момент представлениям процесс распознавания сложных зрительных объектов можно условно разбить на несколько стадий: выделение фигуры из фона – определение характеристических признаков – связывание отдельных характеристик в единое внутреннее представление – сопоставление с хранящейся в памяти информацией – категоризация. Таким образом, успешное распознавание зрительного образа не заканчивается созданием его внутренней копии, лишь категоризация и выявление биологической значимости объекта логически завершают этот процесс.
Так, было показано, что информация о зрительном образе уже на уровне сетчатки и далее в первичной зрительной коре разделяется на два основных потока, каждый из которых заканчивается в определенных структурах головного мозга (Wssle & Boycott, 1991). Дорзальный путь, закачивающийся в теменных отделах коры больших полушарий, участвует в анализе информации о зрительном пространстве и перемещениях объектов в нем, тогда как включенные в вентральный путь нейронные структуры принимают участие в обработке информации, идентифицирующей зрительные образы. На вершине иерархии вентрального пути находятся области передней нижневисочной коры (Ungerleider & Mishkin, 1982). Именно здесь были обнаружены нейроны, селективно реагирующие на появление в поле зрения сложным образом организованных зрительных стимулов, таких как лица человека или обезьяны (Gross, 1972).
Способность к анализу информации о лицах является одной из самых интригующих функциональных особенностей мозга приматов. В процессе эволюции у приматов сформировалась высокоразвитая мимическая мускулатура, которая играет важную роль в обеспечении социальных взаимооотношений у этих животных, ведущих групповой образ жизни. Поэтому в ходе эволюции в мозге приматов возникли специализированные структуры, анализирующие информацию о лицах и их эмоциональных выражениях (Perrett et al., 1992). С помощью методов функциональной магнитно-резонансной томографии удалось локализовать такие структуры как в мозге человека (Kanwisher, 1997; Tsao et al., 2008), так и обезьян (Logothetis et al., 1999; Tsao et al., 2006), а детальные микроэлектродные исследования позволили выявить специфические свойства нейронов-детекторов лиц (Freiwald et al., 2009). Эти нейроны обладают большими рецептивными полями, что дает им возможность анализировать информацию с протяженных участков поля зрения (Gross, 1972). Детекторы обладают свойством инвариантности: изменения параметров освещения зрительных объектов или их размеров не оказывают значительного влияния на их активность (Vogels & Orban, 1996; Vogels & Biederman, 2002). Существует точка зрения, что кодирование информации о сложных зрительных образах происходит не только с помощью увеличения частоты импульсации, но и за счет временных модуляций нейронного разряда (Richmond et al. 1987; Richmond & Optican, 1987; Sugase et al, 1999). Система распознавания таких сложных зрительных стимулов как лица является, по крайней мере частично, врожденной: нейроны-детекторы лиц были обнаружены даже у новорожденных детенышей макак (Rodman, 1991).
В последнее время появился ряд психофизических работ, которые демонстрируют важную роль прототипа или так называемого «усредненного лица» в организации системы распознавания лиц (Leopold et al., 2001). Предполагается, что узнавание индивидуальных лиц происходит путем сравнения их с прототипом, который формируется у субъекта в результате накопления и обработки информации о лицах в процессе жизни. Принцип кодирования информации о сложных зрительных образах, в основе которого лежит сравнение с прототипом, считается универсальным принципом, который широко ипользуется в мозге для формирования внутреннего представления объектов и событий в окружающей среде (Edelman, 1999). В связи с этим особый интерес приобретает исследование роли нейронов-детекторов в кодировании информации о прототипе и индивидуальных лицах.
Исследование сложных свойств нейронов-детекторов предъявляет особые требования к методикам, используемым в таких экспериментах. Перспективным может быть как использование функционального картирования больших популяций нейронов, так и применение электрофизиологических подходов, с помощью которых возможна стабильная одновременная регистрация нескольких отдельных нейронов. Дело в том, что к настоящему моменту получено значительное количество данных, подверждающих гипотезу о кодировании сложных признаков нейрональными ансамблями (Georgopoulos et al., 1986; Nicolelis et al., 1995; Deadwyler & Hampson, 1997). Регистрация популяционного ответа с помощью визуализирующих работу мозга методик, а также одновременное отведение активности нескольких отдельных нейронов позволяет получить ценную информацию о кооперативной работе клеток мозга, направленной на решение определенных сенсорных задач. Особенно важным представляется использование методик, позволяющих наблюдать за активностью идентифицированного нейрона в течение длительного времени. Хронические имплантации микроэлектродов делают возможными такие долговременные наблюдения за активностью (а также ее изменениями) одной и той же клетки. Речь идет о регистрациях активности, которые отставлены друг от друга не только на несколько часов, но и на дни, недели или даже месяцы. Увеличение количества регистрирующих электродов в хроническом эксперименте позволяет получить уникальные данные по сочетанной работе отдельных клеток в составе нейронных ансамблей. В настоящее время развитие экспериментальных технологий позволяет вплотную приблизиться к созданию таких методик (Porada, 2000).
Цели исследования
Целью настоящей работы является детальное изучение принципов, лежащих в основе представления информации о характеристических признаках изображения и сложных зрительных объектах: адаптивности, стабильности и однозначности кодирования.
Задачи работы
1. Изучить с помощью оптического картирования по внутреннему сигналу особенности представления информации об ориентации зрительного стимула в первичной зрительной коре головного мозга кошки. Наблюдение за большими популяциями нейронов позволит оценить степень адаптации детекторной системы первичной зрительной коры к особенностям зрительного окружения.
2. Разработать методику стабильной одновременной регистрации активностей нескольких нейронов с помощью хронически имплантируемых в мозг обезьян микроэлектродов, а также разработать критерии, позволяющие по характеристикам нейронного ответа оценивать стабильность регистрации сигнала от одного и того же нейрона в разные экспериментальные дни.
3. С помощью хронически имплантированных множественных микроэлектродов изучить способность нейронов-детекторов нижневисочной коры сохранять селективные свойства в условиях отсутствия обучающих воздействий и обеспечивать стабильность кодирования информации.
4. Используя математическую модель морфинга лиц создать наборы зрительных стимулов (лиц), которые могли бы позволить в психофизических экспериментах на обезьянах проверить гипотезу о важной роли прототипа («усредненного лица») в распознавании лиц.
5. Используя наборы проверенных в психофизических опытах стимулов, в электрофизиологических экспериментах исследовать настройку нейронов-детекторов лиц и охарактеризовать полученные данные с позиций представлений по кодированию информации о лицах, ключевым звеном которых является использование однозначного кода при участии прототипа.
Научная новизна работы
В представленной работе с помощью современных методических подходов получены принципиально новые данные. Были применены три уникальных методики, позволивших сформулировать оригинальные теоретические выводы об особенностях функционирования зрительной системы: метод оптического картирования коры по внутреннему сигналу, хронические имплантации множественных микроэлектродов и математическая система морфинга лиц. Оптическое картирование по внутреннему сигналу позволяет опосредованно регистрировать активность больших популяций нейронов в ответ на изменяющиеся условия внешней стимуляции. С помощью данного подхода возможно выявление тонкой функциональной организации корковых областей, а также исследование вовлеченности отдельных функциональных модулей коры в процессы переработки информации. Оптическое картирование позволило показать, что популяции нейронов-детекторов в первичной зрительной коре тонко подстроены к статистикам анализируемого зрительного окружения.
По-своему уникален также метод хронических имплантаций множественных микроэлектродов в мозг, который за время выполнения данной работы был успешно адаптирован для использования в длительных экспериментах на обезьянах. Новизна данного подхода заключается в том, что он обеспечивает стабильную регистрацию клеточной активности от одного и того же нейрона. Такого рода данные являются абсолютно уникальными и приоритетными, поскольку позволяют напрямую тестировать не только стабильность свойств нейронов-детекторов, но и позволяют наблюдать за формированием селективности нейронов и ее изменениями у клеток мозга. На основе формы потенциала действия нейрона и распределения межспайковых интервалов были разработаны критерии оценки, которые позволяют выявлять периоды стабильности клеточного сигнала. Впервые напрямую удалось показать, что стимул-специфический паттерн разряда нейронов нижневисочной коры мозга обезьян остается неизменными в течение длительного периода времени: максимальный период наблюдения за такого рода стабильной активностью составил 17 дней.
На основе модели морфинга лиц впервые была разработана шкала лиц с градуальным именением уровня индивидуальности. Изменения уровня индивидуальности были возможны в диапазоне от 0% (прототип или «усредненное лицо») до 160% (карикатура). Осуществлено прямое сравнение результатов психофизических экспериментов по распознаванию лиц, проводимых с человеком и обезьяной. Было показано, что человек и обезьяна используют один и тот же принцип при распознавании лиц: степень сходства с хранящимся в памяти прототипом.
Изучение характеристик настройки нейронов-детекторов лиц в коре мозга обезьян показало, что ключевую роль в активности этих нейронов играет прототип: кривые настройки были центрированы относительно усредненного лица. Были обнаружены два типа нейронов-детекторов лиц в нижневисочной коре обезьян. Одна часть нейронной популяции увеличивала частоту разряда в ответ на усиление индивидуальных черт лица, тогда как другая ее часть реагировала сильнее на появление в поле зрения животного прототипа. Именно сравнение активностей указанных типов нейронов может служить однозначным кодом, лежащим в основе функционирования системы распознавания лиц. По всей видимости, принцип кодирования, основанный на сравнении с прототипом, является универсальным принципом и для распознавания других сложных зрительных объектов.
Теоретическое и научно-практическое значение работы
Основное теоретическое значение данной работы заключается в подтверждении принципов кодирования информации нейронами-детекторами зрительного анализатора млекопитающих, которые могут быть в дальнейшем использованы для создания искусственных интеллектуальных систем распознавания зрительных образов.
Развитие методик визуализации популяционной активности нейронов и картирования мозга интересно с практической точки зрения, в связи с использованием различных модификаций оптического картирования для диагностических целей в нейрохирургии. Ценный методический опыт, накопленный в ходе выполнения данной работы, может быть успешно использован в широком спектре практических задач в будущем.
Методика длительной регистрации клеточной активности может быть использована дла новых вариантов интерфейсов мозг-компьютер и нейропротезов. В настоящий момент для управления внешними устройствами используют электроэнцефалограмму, которая, однако, не позволяет достигать той степени специфичности реакций, которая наблюдается на нейронном уровне. Поэтому создание безопасных внутримозговых зондов может получить широкое применение в будущем. Накопленные в ходе работы данные по биосовместимости имплантатов и стабильности регистрации клеточной активности приближают применение данных технологий в практике.
Основные положения, выносимые на защиту
1. В условиях нормального зрения функциональные модули первичной зрительной коры настраиваются на статистики окружающей зрительной среды, обнаруживая свойство долговременной адаптивности.
2. При отсутствии обучающих воздействий нейроны-детекторы нижней височной коры мозга обезьяны способны длительное время сохранять селективный стимул-специфический паттерн разряда, что обеспечивает стабильность кодирования сенсорной информации.
3. Системы распознавания лиц человека и обезьяны принципиально похожи: в основе распознавания лежит принцип сравнения с прототипом.
4. В качестве однозначного кода нейроны-детекторы лиц используют монотонные изменения частоты импульсации относительно базового уровня разряда, который задается реакцией клеток на прототип.
Апробация работы
Основные материалы диссертации были представлены на Гёттингенских нейробиологических конференциях (1995, 1996, 1998); на 1-ой Тюбингенской конференции по исследованию восприятия (1998); конференциях Американского общества по нейронаукам (Новый Орлеан, 2000; Сан Диего, 2001; Новый Орлеан, 2003); на Сьездах физиологического общества России им. И.П. Павлова (Екатеринбург, 2006; Москва, 2008; Калуга, 2010); на 29-ой Европейской конференции по зрительному восприятию (Санкт-Петербург, 2006); на XIV-ом всемирном Конгрессе по психофизиологии «Olympics of the brain» (Санкт-Петербург, 2008); на Международной конференции «Прикладная оптика - 2010» (Санкт-Петербург, 2010) и апробированы на межлабораторной конференции ИВНД и НФ РАН (Москва, 2010).
Структура и объем диссертации
Диссертация состоит из введения, обзора литературы, описания методик, четырех разделов в главе с изложением результатов, обсуждения, выводов и списка литературы. Общий объем работы составляет страниц. Работа проиллюстрирована рисунками.
МЕТОДИКА ИССЛЕДОВАНИЙ
Подопытные животные: Исследования функциональной анатомии первичной зрительной коры были проведены на 25 взрослых беспородных кошках обоих полов.
Эксперименты по отведению клеточной активности осуществлены на двух взрослых самках яванской макаки (Macaca fascicularis), на четырех взрослых самцах макаки-резус (Macaca mulatta) и на одном взрослом самце игрунки (Callithrix jacchus). Психофизическое тестирование восприятия лиц проведено на одном самце макаки.
Опыты осуществляли в соответствии с положением Института ВНД и НФ РАН о работе с экспериментальными животными с соблюдением принципов гуманности, изложенных в директивах Европейского сообщества (86/609IEС) и одобренных комитетом по медицинской этике.
Эксперименты по оптическому картированию мозга по внутреннему сигналу
Подготовка животного к операции. Первая фаза подготовки животного к операции направлена на обеспечение инициальной анестезии, подавление спонтанного дыхания животного и подключение его к аппарату искусственного дыхания. Запись данных синхронизовали с дыхательным циклом животного, что позволяло исключить возникновение артефактов в записи, связанных с притоком насыщенной кислородом крови к мозгу.
Для подавления вегетативной реакции слюноотделения, вызываемой механическим раздражением полости рта и гортани при интубации, животному внутримышечно вводили атропин. После инициации анестезии в вену передней лапы устанавливали катетер, через который в организм вводили наркотизирующее вещество, обеспечивающее наркоз длительного действия (тиопентал либо пропофол). Для поддержания постоянного водно-солевого баланса и обеспечения животного питательными веществами через внутривенный катетер в организм животного подавали раствор Рингера и глюкозу со скоростью 3-4 мл/час.
В момент прекращения спонтанных дыхательных движений, вызванного внутривенным введением парализующего вещества (ардуан), в трахею животного вводили интубационную трубку, которую затем подключали к аппарату искусственного дыхания. Для контроля за уровнем анестезии и жизненными функциями животного использовали приборы, измеряющие процентное содержание кислорода и углекислого газа в выдыхаемом воздухе, температуру тела животного, частоту сердечных сокращений и уровень насыщения крови кислородом. В течение операции и регистрации данных каждые 1,5-2 часа животному дополнительно внутривенно вводили парализующее вещество.
Расширения зрачка добивались нанесением на поверхность роговицы глаз 1% раствор атропина. Специальные жесткие контактные линзы предохраняли роговицы глаз животного от высыхания.
Операция. Целью операции является обеспечение оптического доступа к поверхности коры больших полушарий.
Дополнительно к системному наркозу осуществляли местную анестезию операционного поля подкожными инъекциями лидокаина. С теменной и затылочной областей черепа животного удаляли участок кожи. Поверхность черепа очищали от мягких тканей и надкостницы.
В черепе высверливали трепанационное отверстие диаметром 7-10 мм с центром, расположенным в координатах: АР = -4 мм, МL = 4 мм (система стереотаксических координат Хорслей-Кларка).
Над трепанационным отверстием с помощью зубоврачебной пластмассы закрепляли шахту, которую позднее заполняли силиконовым маслом и герметично закрывали стеклом. Это позволяло искусственно восстановить целостность черепа и избежать появления в записи оптического сигнала артефактов, связанных со сдвигом поверхности мозга вследствие дыхательных и пульсовых движений.
Шахта представляет собой металлический цилиндр диаметром 3.5 см, высотой 1.4 см и толщиной стенок 2 мм. На внутренней поверхности шахты имеются три отверстия диаметром 1 мм, соединенные с металлическими трубочками, и служащие для удаления излишков масла и компенсации давления на мозг, возникающего при закрытии камеры стеклом. Перед заполнением шахты с поверхности мозга удаляется твердая мозговая оболочка.
Экспериментальная установка для оптического картирования мозга. Экспериментальная установка состоит из CCD-камеры, осветителя, компьютера для предъявления зрительных стимулов, компьютера, осуществляющего регистрацию данных и приборов, осуществляющих контроль состояния животного и уровнем анестезии.
ССD-камера (фирма RedShirtImaging, USA) имеет светочувствительную матрицу, состоящую из 256 х 256 пикселей. Размер матрицы – 6 х 6 мм. Оптической системой камеры является «макроскопа», состоящего из двух фотообъективов, соединенных друг с другом «лицом к лицу». Макроскоп обеспечивает незначительное увеличение (1 – 4 раза) и малую глубину резкости, что необходимо для регистрации оптического сигнала из относительно тонкого слоя коры.
Равномерное освещение препарата осуществлялось при помощи осветителя со сменными световыми фильтрами. Питание лампы осветителя обеспечивалось источником постоянного тока, что исключает флуктуации уровня освещения.
Регистрацию данных и управление камерой осуществляли при помощи компьютера, который через специальный модуль, обеспечивающий получение запускающего сигнала, был соединен с компьютером для зрительной стимуляции. Начало стимуляции происходило после получения запускающего сигнала стимулирующим компьютером от аппарата искусственного дыхания, что крайне важно, поскольку запись данных должна быть синхронизована относительно респираторного цикла, так как он является причиной дыхательных смещений поверхности коры и, следовательно, значительных артефактов в записи оптической активности от поверхности мозга. Таким образом, после получения запускающего импульса от дыхательного аппарата, стимулирующий компьютер включал зрительный стимул и подавал сигнал на начало записи данных регистрирующему компьютеру.
Зрительная стимуляция и регистрация оптического сигнала. После установки на череп животного шахты и заполнения ее силиконовым маслом, CCD-камеру фокусировали для получения изображения поверхности коры.
Зрительные стимулы подавали на экран монитора, находящийся на расстоянии 57 см от глаза животного. При таком расположении один сантиметр экрана соответствует одному угловому градусу.
Для компенсации дефектов зрения, вызванных введением парализующего вещества, при помощи офтальмоскопа подбирали корректирующую линзу и устанавливали ее перед стимулируемым глазом животного для компенсации дефекта зрения.
В качестве зрительных стимулов использовали изображения решеток разной ориентации. Пространственная частота решеток составляла 1.6 на цикл. Запись оптического сигнала производили одновременно с началом зрительной стимуляции. Регистрировали два типа оптического сигнала: «зеленый образ» (длина волны 550 нм) и сигнал, полученный при освещении поверхности мозга светом длиной волны около 600 нм. Регистрация оптического сигнала при освещении поверхности коры головного мозга зеленым светом позволяет получить четкое изображение кровеносных сосудов. Полученное изображение использовали при обработке данных для уменьшения артефактов, связанных с движением крови по поверхностным сосудам.
Оптический сигнал, регистрируемый с поверхности коры головного мозга, имеет метаболическое происхождение, поэтому является медленно развивающимся во времени. По сравнению с электрофизиологическими методами для получения четких изображений необходима длительная стимуляция. В наших экспериментах она была равна 5 секундам. Накопление данных после окончания стимуляции составляло еще 5 секунд. Интервал между предъявлениями зрительных стимулов длился 15-20 секунд.
Принцип анализа экспериментальных данных. Обработка данных осуществлялась при помощи программ, разработанных в лаборатории с использованием пакета MatLab.
Экспериментальные данные представляют собой последовательность видеокадров с разрешением 256х256 пикселей и записанных с частотой 25Гц. Кадры, отобранные для анализа (от 32 до 128 фрэймов), усредняются как внутри одного предъявления стимула, так и для разных предъявлений (до 20 предъявлений определенного стимула в течение одного эксперимента). Так, для построения усредненной карты для одного зрительного стимула используется от 640 до 2560 фрэймов. Усредненный ответ на один тип стимула сравнивается с так называемой картой “cocktail blank”: усредненным ответом на все стимулы. “Cocktail blank” получают при усреднении от 5120 до 20480 фрэймов. Использование большого количества усреднений необходимо, поскольку регистрируемый от коры оптический сигнал составляет 0,6 – 3,0% от исходного уровня освещенности.
Таким образом, большое количество усреднений способствует выделению сигнала из биологического шума и позволяет получать функциональные карты активации коры в ответ на один тип зрительного стимула. В дальнейшем функциональные карты, полученные в условиях стимуляции одним зрительным образом, исследовали на предмет стабильности структуры. Они служили основой для выделения «областей интереса», в которых выделяли участки активированных корковых колонок и определяли общую площадь поверхности коры, занятую колонками.
Для улучшения качества усредненных дифференциальных карт путем повышения их контрастности проводили выравнивание гистограмм распределения яркости для точек изображения в «зоне интереса». Затем в «зоне интереса» определяли площадь активированных корковых колонок при разных пороговых значениях отсечения слабо активированных и не активированных точек. Использовали три уровня такого отсечения: 80, 60 и 40%. Уровень 80% соответствует включению в карту только наиболее активированных точек изображения, активность которых находилась в пределах от 80 до 100% от максимальной, уровень 60% – от 60 до 100%, а уровень 40% – от 40 до 100%, соответственно. При этом 100% активации коры соответствуют наиболее темные точки изображения карт.
Эксперименты по регистрации клеточной активности в коре головного мозга обезьян.
Начальное приручение обезьян к приматологическому креслу. Во время психофизических экспериментов и регистрации клеточной активности животное может находиться в дискомфортной ситуации. Ограничение движений, непосредственная близость эксприментатора, манипуляции при подключении микроэлектродов и эксприментальное окружение играют при этом решающую роль. Ежедневное приучение к этим факторам помогало сгладить стрессовое влияние экспериментальной обстановки. Как правило, для каждой обезьяны время тренировок было вначале достаточно коротким (5 мин.), но постепенно его увеличивали. В итоге животные могли находиться в кресле от 2 до 4 часов. Приучение к креслу проводил экспериментатор, который был хорошо знаком обезьянам. Во время тренировок в качестве подкрепления животные получали особенно любимую ими пищу.
Психофизическое тестирование в экспериментах по распознаванию лиц. В данных экспериментах принимали участие две обезьяны: N97 и C99, каждую из которых тренировали с индивидуальным набором лиц. В результате тренировки обе обезьяны должны были приобрести навык к различению четырех индивидуальных лиц, однако к концу тренировки, несмотря на большое количество предъявлений стимулов, лишь обезьяна N97 в полной мере овладела навыком различения четырех индивидуальных лиц.
Поведенческое тестирование представляет собой адаптированный для обезьян вариант задачи форсированного выбора из четырех альтернатив. При предъявлении на экране монитора лица, обезьяна с помощью двух рычагов, каждый из которых мог двигаться в двух направлениях (отжатие/притягивание), сигнализировала о том, к какому одному из четырех выученных лиц относится данный стимул. После того как обезьяна с высокой долей вероятности (90-95%) давала правильный ответ, ей начинали предъявлять видоизмененные лица, у которых уровень индивидуальности варьировал. Стимулы были подготовлены с помощью базы данных лиц (Face Database, Тюбинген, Германия) Института биологической кибернетики общества Макса Планка (http://faces.kyb.tuebingen.mpg.de/index.php).
Психофизическое тестирование порогов восприятия у обезьяны N97 проводили следующим образом. После короткого предупреждающего звукового синала на мониторе компьютера появлялась фиксационная точка, и от животного требовалось задержать взгляд на ней. После паузы длиной 500 мсек на экране возникало изображение лица, длительность предъявления которого составляла 200 или 400 мсек.
В качестве подкрепления за правильную поведенческую реакцию животное получало яблочный сок. В среднем в течение одного экспериментального дня животное выполняло от 800 до 1200 попыток. Обезьяна N97 получала подкрепление по специальной схеме «варьируемого подкрепления» в зависимости от сложности и правильности выполнения задачи по различению лиц.
Эксперименты с фиксацией взгляда. В электрофизиологических экспериментах по изучению селективных свойств нейронов в нижневисочной коре важно иметь возможность регистрировать активность нервных клеток в повторяющихся условиях зрительной стимуляции. С этой целью две обезьяны – N97 и E98 – были обучены выполнять простейшую задачу по фиксации взляда, в которой положение глаз измеряли с помощью гибкой микропроволоки, имплантированной в склеру одного из глаз.
Каждая попытка в фиксационной задаче начиналась с появления на мониторе компьютера небольшого светлого пятна – фиксационной точки – размером 0.5° x 0.5°. От обезьяны требовалось зафиксировать взгляд на точке в течении всего периода регистрации данных. При отклонении взляда от фиксационной точки более чем на 1° попытка автоматически прерывалась. После задержки в 700-1000 мсек, первый зрительный стимул предъявляли на мониторе компьютера, а затем, после паузы в 600 мсек, второй стимул появлялся на экране. Длительность предъявления второго стимула была такой же как и первого.
Стимулы представляли собой наборы различных изображений, включавших в себя лица, цветные фотографии животных, объектов и геометрических паттернов. Если обезьяна удерживала фиксацию в течение всего периода регистрации электрофизиологических данных, то в качестве подкрепления она получала каплю яблочного сока. В среднем обезьяна во время одного экспериментального дня выполняла до 1200 попыток (разброс составляет от 300 до 3000 попыток в день), что соответствует предъявлению до 2400 стимулов в день.
Хронические имплантации множественных микроэлектродов. Изучение зрительного восприятия в разных условиях зрительной стимуляции важно само по себе, однако настоящую ценность такие исследования приобретают при параллельной записи клеточной активности и последующей ее обработки с целью выявления закономерностей в работе зрительного анализатора. На наш взгляд, значительного успеха можно добиться, если использовать множественные хронически имплантированные микроэлектроды, которые позволяют длительное время регистрировать активность от одного и того же нейрона.
Для таких хронически имплантированных электродов крайне важно, чтобы электроды удерживались долгое время в фиксированной позиции. Это требование определяет всю принципиальную конструкцию имплантата, который состоит из следующих частей:
а. отводящих электродов и связанного с ними микроштекера, с помощью которого происходит подключение животного к регистрирующей аппаратуре;
б. несущей трубки, в которой электроды подводят к месту имплантации и позднее удерживают в одной позиции.
в. конструкции для закрепления микроштекера на поверхности черепа и для его защиты от повреждений.
Для изготовления множественных электродов использовали никельхромовую проволоку диаметром 12 мкм, изолированную полиимидом («Isaohm», фирма «Izabellenhtte», Дилленбург). Использование этой крайне тонкой проволоки нужно для достижения высокого сопротивления на кончике электрода, что является необходимым условием для отведения слабых электрических сигналов. Хорошая биосовместимость изолятора полиимида была доказана ранее (Babb & Kupfer, 1984).
Проволоку нарезали кусками по 10 см. На одном конце удаляли изоляцию на протяжении 3-4 миллиметрах. Пучок электродов состоял из 32 или 64 отдельных проволок. Четырех- либо восьми-контактные штекеры («Buchsenleiste gerade 8po», фирма Brsig Gmbh, Neckarsulm) с предварительно укороченными контактами склеивали вместе с помощью двухкомпонетного эпоксидного клея («Plus endfest 300», фирма UHU).
Припаивание отдельных никельхромовых проводков проводили под микроскопом. Для пайки использовали специальное средство на цинк-хлорной основе (фирма «Castolin», Lausanne) и серебряно-цинковый припой (фирма «Castolin», Lausanne). После того как все провода были припаяны, остатки средства для пайки удаляли с помощью ультразвуковой ванны. Очищенные таким образом контакты покрывали в некоторых случаях акриловым цементом («Grip Cement», фирма «Caulk/Dentsply», Milford, USA) или эпоксидной смолой.
На следующей стадии электроды склеивали вместе, формируя подобие кисти, при этом передние части элетродов были свободны на протяжении 6 мм, тогда как оставшиеся части до самого штекера оказывались склеенными вместе. В качестве клея мы использовали растворенный в дихлорэтане плексиглаз.
Электроды доставляли к месту имплантации в несущей трубочке, в качестве которой служила обыкновенная игла для инъекций (внешний диаметр - 0.5 мм). Перед имплантацией электроды закрепляли в стереотаксическом приборе и погружали в стерилизирующий раствор.
В экспериментах на обезьяне был использован специальный имплантат с микроманипулятором. Преимущество данного имплантата заключается в том, что в нем сочетаются стабильность удержания пучка электродов и возможность дополнительной корректировки их положения в мозге после имплантации. Для корректировки положения пучка электродов можно использовать микроманипулятор, а также была предусмотрена возможность корректировки положения несущей трубочки в мозге. Микроманипулятор был защищен от повреждений специальной шахтой, выполненной из сплава титана. Нижняя поверхность шахты была сконструирована таким образом, чтобы идеально соответствовать поверхности черепа обезьяны N97.
Регистрация внеклеточного сигнала и анализ экспериментальных данных в экспериментах по изучению свойств нейронов нижневисочной коры у макак-резус. Каждый день с имплантата снимали защитный колпак (обезьяна N97) и через микроштекер животное подключали к одному из усилителей (MCP-8 Plus, фирма AlphaOmega, Израиль; FAX32X, фирма Multi Channel Systems GmbH, Германия). Все 64 микроэлектрода в имплантированном пучке исследовали на предмет выявления одиночных нейронов и сравнивали с регистрациями в предыдущие дни. Каналы, на которых обнаруживали потенциалы действия, оцифровывали с частотой 22 кГц и сохраняли на жестком диске. Выделение спайков проводили после окончания эксперимента с помощью программ, написанных с использованием пакетов Tcl/Tk (http://www.tcl.tk) и Matlab (MathWorks, Natick, MA). Спайки-кандидаты для сортировки выделяли из фоновой активности с помощью порогового критерия. Порог для потенциала устанавливали на уровне двух стандартных отклонений от усредненного потенциала фонового шума. С выделенными таким образом спайками-кандидатами проводили анализ методом главных компонент. Этот метод позволяет выявлять набор функций, которые наилучшим образом описывают сходство форм спайков между собой. По результатам данного анализа строили графики, которые представляют собой проекции индивидуальных форм спайков на оси описательных функций. Вся популяция форм спайков выглядит на таких графиках как облако (или группа облаков), состоящее из отдельных точек. Кластеры, соответствующие той или иной форме спайка, отделяли друг от друга вручную, задавая границы вокруг каждого облака. После выделения кластеров чистоту сортировки отдельного нейрона проверяли, используя гистограммы межстимульных интервалов.
Делая выводы о том, что регистрация от одного и того же нейрона велась в течение нескольких последовательных дней, мы применяли консервативные критерии. Решение принимали на основании сходства между формами спайков и гистограмм межспайковых интервалов. В случае глобальных изменений амплитуды и формы потенциалов действия, данный нейрон отвергали как кандидата для последующего анализа между разными днями регистраций.
Отсортированные спайки использовали для построения растерных диаграмм и перистимульных гистограмм. Достоверность вызванных зрительной стимуляцией ответов нейронов подтверждали с помощью t-критерия и критерия Колмогорова-Смирнова.
Показатель сходства, основанный на корреляции. Для количественной оценки отношений между селективными ответами отдельных нейронов мы вычисляли показатель, который принимал в расчет не только изменения в силе ответа (соответствует частоте разряда нейрона), но так же и временные модуляции частоты, вызванные каждым стимулом. Этот показатель вычисляли на основе постстимульных гистограмм, рассчитанных для всех стимулов. Так, для каждого стимула строили гистограммы распределения частот разряда на интервале времени между 40 и 640 мсек с момента начала предъявления зрительного стимула. Ширина временного бина составляла 40 мсек. Из полученных таким образом 16-ти элементных векторов формировали матрицу, в которой строки соответствовали стимулам, а столбцы – временным бинам. С помощью функции «corrcoef» пакета Matlab вычисляли парные коэффициенты корреляции между матрицами. Показатель сходства между матрицами рассчитывали для трех разных ситуаций. В первом случае, показатель сходства рассчитывали между матрицами, построенными для двух нейронов, активность которых регистрировалась одновременно в одном эксперимента, но с разных электродов. Во втором случае, сходство между ответами вычисляли для двух нейронов, активность которых была записана параллельно на одном и том же электроде. И в третьем случае, коэффициент корреляции рассчитывали для матриц полученных для одного и того же нейрона, активность которого регистрировали в разные экспериментальные дни. В случае сравнения матриц полученных для одного и того же нейрона анализ проводили для первого и последнего дня регистраций данной клетки. Высокая степень сходства соответствует значениям показателя близким к единице, тогда как отсутствие сходства между матрицами селективности соответствует значениям показателя близким к нулю.
ОСНОВНЫЕ РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Изучение особенностей функциональной архитектуры первичной зрительной коры методом оптического картирования по внутреннему сигналу
В первой серии опытов по оптическому картированию первичной зрительной коры у 9 кошек был зарегистрирован оптический сигнал с поверхности мозга в ответ на зрительную стимуляцию.
По полученным данным были построены функциональные карты ориентационной чувствительности. Рисунок функциональных карт различен для индивидуальных животных. Карты отличаются друг от друга как по частоте, равномерности распределения по поверхности коры и форме активных областей, так и по их выраженности (контрасту). Такие различия в картах могут быть связаны с индивидуальными особенностями животного.
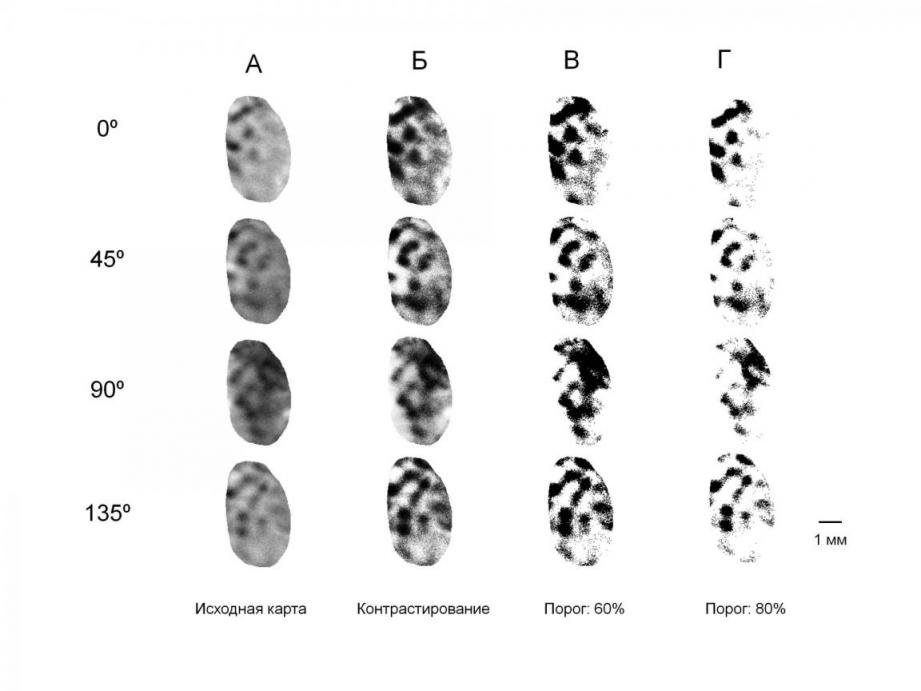
Поскольку оптический сигнал является слабым, наиболее четкие функциональные карты могут быть получены при большом количестве усреднений, что требует большого числа предъявлений зрительных стимулов. В литературных данных количество предъявлений одного стимула может доходить до 100. Длительность эксперимента возрастает с числом используемых стимулов, поэтому важно, чтобы во время всей процедуры накопления данных функциональные карты, получаемые в результате обработки, были стабильны. Для оценки стабильности регистрации оптического сигнала строили функциональные карты по данным, полученным с промежутком в несколько часов. При анализе таких карт было обнаружено, что количество и конфигурация активных областей первичной зрительной коры при стимуляции не изменяются в течение одного эксперимента. Таким образом, за все длительное время проведения эксперимента (до 12 часов) получаемые карты ориентационной чувствительности остаются стабильными, что позволяет обрабатывать совместно данные, записанные с промежутком в несколько часов, и подтверждает то, что регистрируемый сигнал является следствием активации нейронов стриарной коры.
Рисунок 1. Усредненные функциональные карты участка первичной зрительной коры мозга кошки при стимуляции глаза решётками разной ориентации. Активированные ориентационные колонки коры соответствуют зонам затемнения коры. А - исходные усредненные карты, Б - карты после контрастирования, В - результат выделения активированных колонок с порогом 60% и Г – то же, что на В, с порогом 80%.
Для точной оценки площади активированной коры мы провели выделение участков карт, соответствующих разной степени активации (рис. 1). Для сравнения мы использовали уровни отсечения, при которых на картах оставались только наиболее активированные точки изображения (порог отсечения – 80% от максимума), либо исключали сравнительно малоактивные или неактивные точки изображения (порог отсечения 60%), либо – только наименее активированные точки на функциональной карте (порог отсечения 40%). Нулевой гипотезой для сравнения площадей коры, занятых базисными и наклонными ориентационными колонками, служило выдвинутое ранее в литературе предположение об анизотропии в представительстве ориентаций: увеличение в пользу горизонтально и вертикально ориентированных линий.
По этим данным для каждой карты, полученной при стимуляции решеткой определенной ориентации, рассчитывали площадь, занимаемую активированными участками коры относительно общей площади «зоны интереса» (рис. 2). Под зоной интереса следует понимать выделенный экспериментатором участок функциональной карты, свободный от крупных сосудов и глобальных артефактов. На рис. 2 сравниваются площади коры, активированные базисными и наклонными ориентациями для 3-х уровней порога отсечения.
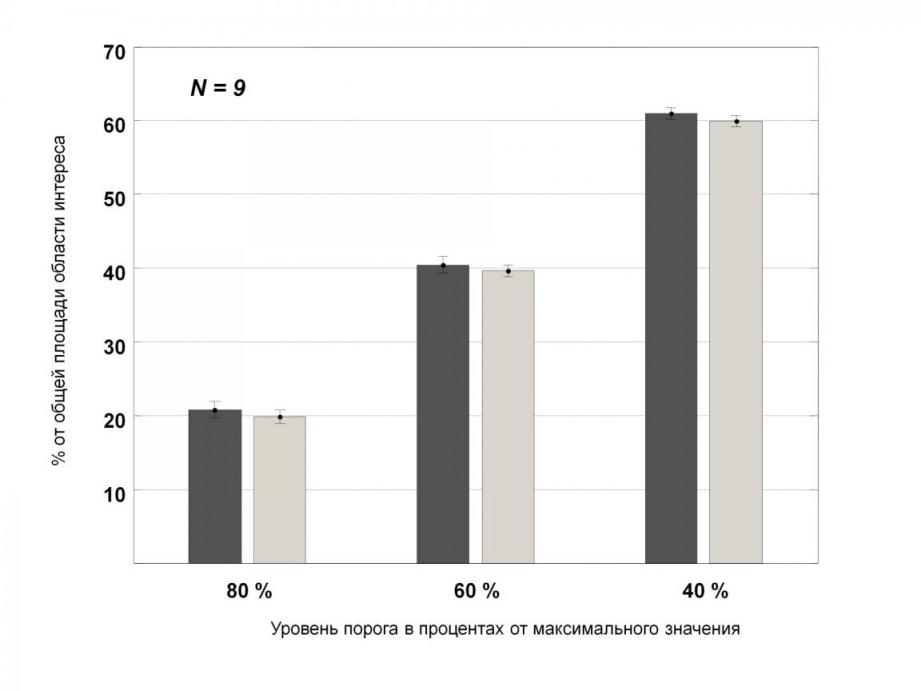
Рисунок 2. Соотношение площади первичной зрительной коры, активированной при предъявлении линий двух базисных ориентаций – вертикали и горизонтали (темно-серые столбцы; в процентах относительно общей площади «зоны интереса», принятой за 100%) и диагональных ориентаций (светло-серые, в процентах). В каждом столбце суммированы данные по 9 животным. Приведены данные для трех уровней порога выделения активных ориентационных колонок коры (80, 60 и 40% от максимального значения активации).
Данные статистической оценки показали, что колонки, активированные базисными и наклонными ориентациями, имеют близкую площадь. Так, при пороге отсечения 80% вероятность ошибки различия средних оценок p = 0.36, при пороге 60% p = 0.38, а при наименее строгом критерии (40%) – p = 0.14. Это означает, что при всех уровнях отсечения относительно слабо активированных и не активированных точек карты средние относительные оценки тангенциальной площади активированных корковых колонок при действии основных и диагональных ориентаций стимула достоверно не различаются.
Таким образом, достоверные различия между представительством в коре разных ориентаций не выявлены. Рассчитанный для всех пороговых значений коэффициент корреляции между площадью коры, активированной линиями базисных и диагональных ориентаций, составил 0.99 (p<0.00005), а уравнение регрессии y=-0.004+0.988*x. Это также указывает на крайне близкую величину средней тангенциальной площади колонок, нейроны которых настроены на выделение базисных и диагональных ориентаций.
Таким образом, нулевая гипотеза – предположение об увеличенном представительстве в первичной зрительной коре мозга кошки нейронов, которые анализируют две базисные ориентации (горизонталь и вертикаль), – в условиях наших опытов не подтвердилась и может быть отвергнута.
Стабильность свойств нейронов-детекторов нижней височной коры.
Стабильное отведение активности одиночных нейронов на примере регистраций в нижневисочной коре макак-резусов. Накопленные в ходе первых экспериментов по имплантации электродов в первичную зрительную кору макак и игрунки данные были использованы для подготовки имплантатов, которые позднее были введены в нижневисочную кору макак-резусов. Несмотря на различия в конструкциях для фиксации пучка электродов и микроштекера на поверхности черепа животных, в обоих случаях – обезьяны N97 и E98 – были возможны стабильные долговременные регистрации клеточной активности. Полученные данные позволили нам количественно оценить стабильность внеклеточно регистрируемых форм потенциалов действия (спайков) и выработать критерии для принятия решений об стабильности регистрации сигналов от одного и того же нейрона в разные экспериментальные дни.
В первую очередь хотелось бы обратить внимание на общие характеристики регистрируемых с помощью микропроволок нейронных сигналов. На рисунке 3А показаны сигналы, которые регистрировали параллельно на девяти микроэлектродах в течение одного эксперимента. Данные были отфильтрованы с целью удаления низкочастотных компонентов сигнала. Таким образом сигнал на отдельном электроде составлен из высокочастотного нейронного шума и нейронной активности в виде спайков. На четырех электродах можно обнаружить потенциалы действия большой амплитуды. Более того, на указанных электродах хорошо различимы спайки разной амплитуды, которые приходят от разных близкорасположенных нейронов. На остальных каналах амплитуда потенциалов действия, как и частота разряда нейронов, была меньше. Однако, и в этом случае спайки можно было легко выделить из шума с помощью порогового критерия.
Одним из показателей качества сигнала может служить отношение сигнал/шум. В нашем случае сигнал – отдельный потенциал действия. Амплитуды потенциалов действия мы сравнивали с амплитудой нейронного шума, который также регистрирует микроэлектрод во время экспериментов. Распределения отношений сигнал/шум показаны на рисунке 5Б. Чаще всего мы регистрировали потенциалы действия, амплитуда которых в 4 раза превышала величину нейронного шума. Кроме того были случаи, когда амплитуда спайка в десять раз превышала амплитуду нейронного шума. Таким образом, качество регистраций с помощью микропроволок приближается к таковому у заостренных электродов. Разница лишь заключается в том, что заостренные электроды с помощью микроманипуляторов можно приближать к нейрону и получать сигнал большой амплитуды, тогда как в нашем случае мы стремились получить сигнал от максимально возможного количества отдельных нервных клеток.

Рисунок 5. Демонстрация отношения сигнал/шум. А. - примеры регистрации активности в нижней височной коры обезьяны E98. На четырех электродах – B7, D2, D3, E8 – было возможно регистрировать потенциалы действия одновременно от нескольких нейронов. Б. – суммарный график, показывающий распределения отношений сигнал/шум, полученных за все время экспериментов. Гистограмма черного цвета обобщает данные по всем случаям успешной регистрации нейронного сигнала, тогда как гистограмма серого цвета соответствует тем случаям, когда нейроны отвечали на зрительную стимуляцию. На вставках показаны типичные формы спайков, для которых соотношения сигнал/шум соответствовали определенным значениям гистограммы.
Основной целью отведений клеточной активности с помощью хронически имплантированных электродов является стабильное отведение сигнала от одной и той же клетки в течение длительного периода времени. В ходе данной работы мы обнаружили, что часто на одном и том же электроде, но в разные экспериментальные дни, можно обнаружить формы потенциалов действия, которые очень похожи друг на друга.
На рис. 4 показан пример таких регистраций. Так на верхних двух рядах графиков показаны формы потенциалов отдельных нейронов, которые регистрировали в течение 17 дней на разных микропроволоках пучка электродов, помещенных в нижнюю височную кору обезьяны N97. При сравнении форм спайков следует отметить, что потенциалы действия разных нейронов отличаются друг от друга. Так потенциал действия нейрона n97B501 имеет более компактную форму, тогда как потенциал действия n97I801 развивается во времени медленнее. Если сравнивать спайки на одном электроде, то видно, что изменения между экспериментальными днями затрагивают скорее амплитуду спайков, но не их форму. Это справедливо и для двух нейронов, формы потенциалов действия которых показаны на двух нижних рядах графиков. Один из этих нейронов был зарегистрирован в мозге обезьяны E98, a второй – обезьяны N97.

Рисунок 4. Стабильная регистрация потенциалов действия одиночных нейронов. Показаны примеры потенциалов действия четырех клеток. Первые два нейрона – n97I801 и n97B501 – регистрировали параллельно на двух разных электродах в течение 17 дней. На нижних примерах показаны нейроны (e98D302 и n97D401), стабильная регистрация которых была возможна в течение 4 дней.
Изменения амплитуды спайка могут быть вызваны тем, что регистрирующий кончик электрода незначительно смещается относительно тела клетки. При этом форма спайка может оставаться той же. Для того чтобы иметь возможность сравнивать спайки одной формы, но разной амплитуды, мы нормировали спайки относительно их амплитуды. Полученные в результате нормировки усредненные спайки для разных экспериментальных дней сравнивали между собой. Дополнительно сравнивали нормализованные распределения межспайковых интервалов, полученные в результате анализа данных для данного нейрона в разные экспериментальные дни. Распределение межспайковых интервалов является индивидуальным функциональным показателем нервной клетки.
Как оказалось формы нормализованных потенциалов действия и нормализованные распределения межспайковых интервалов для одного и того же нейрона обнаруживают высокую степень сходства для разных экспериментальных дней. Для выработки критериев оценки стабильной регистрации спайков одной формы в разные экспериментальные дни мы провели количественный анализ сходства на популяции нейронов. С этой целью сравнивали форму потенциалов действия, форму гистограмм межспайковых интервалов и амплитудных соотношений для потенциалов действия, которые были зарегистрированы на одном электроде. В качестве контроля использовали распределения вышеназванных величин, которые были получены при сравнении параллельно регистрируемых на разных электродах нейронов.
Установлено, что при стабильной регистрации внеклеточных потенциалов действия определенной формы на индивидуальном электроде соотношения амплитуд спайков в разные дни стремится к единице, т.е. происходят лишь незначительные флуктуации величины спайков. Кроме того, обнаружены четкие различия между распределениями евклидовых расстояний между нормированными формами спайков и формами гистограмм межспайковых интервалов в зависимости от того насколько стабильной была регистрация активности. С помощью t-критерия Стьюдента показаны достоверные различия между вышеназванными характеристиками стабильного нейронного разряда по сравнению с контрольными данными, полученными с разных электродов. Это позволило нам выработать критерии оценки стабильной регистрации спайков одной формы в разные экспериментальные дни и применить их на практике для дальнейшего анализа данных, направленного на изучение селективных свойств нейронов-детекторов нижней височной коры.
Временные модуляции и стабильность ответов нейронов нижней височной коры
Локализация электродов в мозге. С помощью магнитно-резонансной томографии установлено, что у животного N97 пучок электродов находился в области TЕ рядом с передней частью средней височной борозды. У животного E98 пучок размещался на дне верхней височной борозды, по всей вероятности в области PGa. Несмотря на то, что нейроны указанных областей, возможно, играют различную роль в опознании зрительных образов, ответы клеток в обеих локализациях демонстрировали: 1) выраженные селективные реакции по отношению к определенным стимулам; 2) сложные динамические модуляции разряда, вызванные зрительной стимуляцией; 3) стабильность ответов в разные дни регистрации. Поэтому для последующего анализа стабильности селективных свойств нейронов было принято решение объединить популяции нейронов, полученные в экспериментах на разных животных.
Общая характеристика нейронов нижневисочной коры. Во время экспериментов, которые на животном N97 длились около одного года, а на обезьяне Е98 регистрация клеточной активности была возможна в течение трех месяцев, мы зарегистрировали активность 158 нейронов. Речь идет только о тех нейронах, которые демонстрировали четкие и статистически достоверные ответы хотя бы на один из зрительных стимулов, использованных в эксперименте. Так у обезьяны Е98 в коре, расположенной в глубине верхней височной борозды, было зарегистрировано 66 нейронов, тогда как у животного N97 в передней части средней височной борозды - 92 нейронов.
Показатель распределенности кодирования является уникальной характеристикой нейрона-детектора нижневисочной коры, которая отражает его способность реагировать увеличением частоты разряда на часть зрительных образов из всего набора тестовых стимулов. В идеальном случае – при использовании бесконечно большого набора тестовых стимулов – показатель распределенности кодирования будет отражать степень вовлеченности данного нейрона в работу нейронных сетей, осуществляющих обработку зрительной информации. Значение показателя распределенности кодирования близкое к единице означает, что нейрон реагирует на все стимулы тестового набора. Тогда как значения этого показателя, приближающиеся к нулю, говорят о высокой специфичности реакций данного нейрона. Следует отметить, что нейроны передней части средней височной извилины (обезьяна N97) демонстрируют более высокий уровень распределенности кодирования. Среднее значение этого распределения равно 0.56, тогда как для распределения, полученного при анализе нейронов верхней височной борозды (обезьяна E98), среднее значение равно 0.36. Тест Колмогорова-Смирнова показал статистически значимые различия между двумя распределениями, что может свидетельствовать о том, что свойства нейронных популяций двух указанных областей имеют различия в плане распределенности кодирования. Вероятно, нейроны из глубины верхней височной извилины (обезьяна E98) являются более специализированными, чем нейроны передних отделов нижневисочной коры.
Высокий уровень показателя распределенности кодирования у нейронов нижней височной коры подразумевает наличие дополнительных возможностей для кодирования информации, поскольку кодирование по типу «один нейрон – один зрительный объект» кажется маловероятным. И действительно, во многих случаях нейроны обнаруживают значительные стимул-специфические модуляции частоты импульсации. На рис. 5А показаны перистимульные гистограммы, построенные по результатам накопления данных при регистрации активности одного нейрона. Перистимульные гистограммы (и соответствующие им зрительные стимулы, рис. 5Б) выстроены в ряд по мере убывания средней частоты разряда на интервале усреднения 50-450 мсек после предъявления зрительного стимула. Видно, что ответы нейрона на разные стимулы различаются по латентности, наличию острых пиков в импульсации, а так же по чередованию периодов возбуждения и торможения. В некоторых случаях (например, первая перистимульная гистограмма слева в верхнем ряду и четвертая гистограмма справа в этом же ряду) активность нейрона носит осцилляционный характер. Одновременно, во второй слева гистограмме нижнего ряда четко выражена тормозная пауза.

Рисунок 5. Примеры разнообразных реакций нейрона (n97D601, животное N97) на тестовый набор зрительных стимулов. Перистимульные гистограммы (А) и соответствующие им стимулы (Б) упорядочены по мере убывания средней частоты разряда в интервале времени между 50-ой и 450ой мсек с момента начала зрительной стимуляции.
Подобные временные модуляции нейронного разряда не являются уникальным свойством лишь одного нейрона, и дополнительные примеры этого феномена показаны на рис. 6, иллюстрирующем стабильность селективных свойств нейронов
Стимул-специфические временные модуляции частоты разряда нейронов нижневисочной коры были использованы для анализа другой специфической характеристики этих детекторных клеток – стабильности селекттивности.
Стабильность селективности нейронов-детекторов нижневисочной коры. В ходе экспериментов мы часто наблюдали стабильно регистрируемые формы сайков на одном и том же электроде. Такие формы спайков мы считали принадлежностью одного и того же стабильно регистрируемого нейрона. Активность таких нейронов исследовали, объединяя данные разных дней регистрации в общие растерные диаграммы и перистимульные гистограммы. Примеры растерных диаграмм и перистимульных гистограмм четырех стабильно регистрируемых нейронов можно видеть на рис. 6. Фоновые полоски разного цвета на растерных гистограм обозначают разные дни регистрации.
Интересно, что в разные дни регистрации регулярно обнаруживалась сохранность селективности как высокочастотных (частота разряда нейрона в определенный интервал времени после предъявления зрительноого стимула), так и низкочастотных компонентов нейронных ответов (появление пиков активности, тормозных пауз и участков стабильной частоты разряда). Напомним, что мы включили в анализируемый материал только данные для надежно изолированных одиночных нейронов со стабильной формой спайка. Так, форма спайка нейрона n97I801, активность которого показанна на рис. 6А, была стабильной в течение по крайней мере 17 дней. Стабильность нейронных ответов выражена здесь в точном временном расположении («тайминге») отдельных спайков, что на растерных диаграммах ведет к появлению вертикальных линий, а на перистимульных гистограммах - острых пиков.
Существенно, что эти динамические аспекты вызванного стимулом ответа нейрона сохраняют высокую временную точность, несмотря на большое число синаптических переключений, которые потенциально должны служить источником шума и способствовать накоплению ошибок по мере перемещения сигнала от сетчатки через первичную зрительную кору к нижней височной коре. Можно предположить, что такая временная точность динамического паттерна является продуктом более позднего анализа и появляется в локальных нейронных сетях коры на продвинутых стадиях обработки зрительного сигнала.

А.

Б.
Рисунок 6. Разнообразные и стабильные ответы нейронов в нижевисочной коре обезьян N97 (А) и E98 (Б). Под каждым из зрительных стимулов показаны растерные диаграммы импульсации, вызванной предъявлением этого образа. Цветом показаны данные разных экспериментальных дней. Видно, что реакции стабильны во время всего периода наблюдения (до 17 дней). Под растерными диаграммами приведены перистимульные гистограммы ответов.
Стабильность селективности в популяции нейронов. Для количественной оценки сходства между сложными динамическими паттернами ответов нейронов нижневисочной коры проводили их сравнительный анализ в трех разных ситуациях:
а) для пар нейронов, записанных одновременно, но на разных электродах (рис. 7А);
б) для пар нейронов, активность которых одновременно регистрировали на одном и том же электроде (рис. 7Б);
в) для активности одиночного нейрона, наблюдение за которой вели в течение нескольких дней (рис. 7В).

Во всех этих случаях с помощью корреляционного анализа сравнивали матрицы, полученные на основе постстимульных гистограмм. В результате, при оценке сходства индивидуальной селективности нейронов принимали в расчет не только среднюю частоту разряда нейрона в ответ на стимуляцию, но и сложные временные модуляции частоты возникновения потенциалов действия.
Рисунок 7. Оценка стабильности ответов нейронов нижневисочной коры. Для количественной оценки стабильности использовали показатель, рассчитываемый по матрицам, составленным из постстимульных гистограмм (А). Показатель сходства между матрицами селективности рассчитывали между матрицами, которые были получены при (Б) параллельной регистрации активности двух нейронов на разных электродах, (В) одновременной регистрации активности близко расположенных нейронов на одном электроде, (Г) стабильной регистрации активности одного и того же нейрона в разные дни.
Было обнаружено, что сходство матриц у одного и того же нейрона выше, чем для одновременно регистрируемых на одном электроде нейронов с разной формой потенциалов действия. Коэффициент сходства был еще ниже для матриц, полученных от записанных с разных электродов нейронов. Среднее значение индекса сходства для стабильно регистрируемых нейронов составило 0.67, тогда как среднее для индекса сходства селективности разных нейронов равно 0.31 (регистрация на одном электроде) и 0.18 (разные электроды).
Исследования стабильности кодирующих свойств нейронов-детекторов нижней височной коры показали, что как для кривых настроек, которые были построены по усредненным данным, так и для модуляций разряда нейронов обнаруживается повторяемость характеристик между разными днями регистраций. При этом максимальная сохранность в нашем случае наблюдалась в течении 17 дней. Таким образом, впервые напрямую было продемонстрировано уникальное свойство детектирующих клеток - стабильность представления информации о сложных зрительных образах.
Кодирование информации об индивидуальности лиц в мозге обезьян
Поведенческий эксперимент по распознаванию лиц обезьяной N97. В психофизических экспериментах по распознаванию лиц основным параметром, который градуально изменяли в ряду стимулов, являлся «уровень индивидуальности». Индивидуальность человеческого лица задается целым рядом отдельных характеристик. В простом случае это могут быть такие параметры как ширина и высота овала лица, ширина подбородка, расстояние между глазами, ширина рта и т.д. В использованной нами модели морфинга лиц уровень индивидуальности задается сложным образом всей совокупностью точек, описывающих то либо иное лицо. Взаимное расположение точек в трехмерном пространстве, их яркостные и цветовые характеристики играют при этом важную роль. Четкое математическое описание этих взаимоотношений позволяет градуально менять уровень индивидуальности определенного лица. Таким образом, появляется возможность построить кривую зависимости правильности распознавания лица испытуемыми от уровня его индивидуальности. Предполагается, что, как и в классическом психофизическом эксперименте, градуальное увеличение уровня индивидуальности будет вызывать эквивалентные изменения в правильности распознавания конкретного лица.
Данные, полученные на семи испытуемых и опубликованные ранее (Leopold, 2001), приведены в левой части рис. 8. В правой части рисунка в том же масштабе представлены данные, полученные в поведенческих экспериментах на обезьяне N97. Видно, что закономерности опознания человеческих лиц животным и человеком сходны. В первую очередь рассмотрим серые кривые двух графиков. Они соответствуют простым психофизическим экспериментам выбора из четырех образцов, описанным в методической части данной работы. После предъявления на экране лица испытуемый (или животное) должны были сигнализировать нажатием на кнопку (или рычаг) какое из четырех ранее выученных лиц он видит. Форма кривой является типичной для психофизических экспериментов: градуальное приращение силы стимула («уровня индивидуальности лица») вызывает улучшение его распознавания. Поскольку в эксперименте были использованы четыре альтернативных возможности ответа, то уровень случайных реакций находится на 25%. Интересно, что и человек, и обезьяна по настоящему случайным образом отвечают на предъявление прототипа («усредненного лица»), который по определению полностью лишен черт индивидуальности.

Рисунок 8. Психометрические кривые зависимости правильности распознавания лица от уровня его индивидуальности. В левой части рисунка представлены экспериментальные данные, полученные на 7 испытуемых, а в правой части рисунка – данные опытов с обезьяной N97. И в том и в другом случаях кривые серого цвета соответствуют экспериментам выбора из четырех альтернатив. Кривые черного цвета получены в задаче с адаптацией к антилицу. Черные кружки соответствуют экспериментам со специфической адаптацией, а черные квадраты - неспецифической. Различия во времени адаптации между человеком и обезьяной (5 и 4 секунды, соответственно) связаны с тем, что обезьяне труднее выдержать длительную фиксацию взгляда.
Психометрические кривые человека и обезьяны имеют разный угол наклона. Более пологая кривая обезьяны может свидетельствовать о ее меньшей способности к распознавании человеческих лиц, связанной с видоспецифическими особенностями системы распознавания лиц. Порог уверенной детекции индивидуальных лиц у человека находится на 11%-ом уровне индивидуальности, а у обезьяны – на 17.3%-ом. Специфическая адаптация к антилицу вызывает сдвиг порогов восприятия как человека (Leopold, 2001), так и обезьяны (психометрические кривые черного цвета с точками в виде черных кругов). В то же время неспецифическая адаптация ухудшает распознавание индивидуальных лиц (кривые черного цвета с точками в виде черных квадратов).

Нейронная активность в нижневисочной коре при предъявлении лиц. Чтобы проверить, как описанные преобразования лиц влияют на ответы нейронов нижневисочной коры, мы исследовали реакции клеток при действии лицевых стимулов, соответствующих разным точкам в модельном пространстве лиц. Каждый день животным предъявляли набор лиц, расположенных на четырех разных осях карикатуризации, соответствующих четырем разным индивидуумам. Для каждой из обезьян – N97 и C99 - использовали индивидуальный набор стимулов. У одного из животных (C99) дополнительно проводили тестирование с лицами, которые были расположены на тангенциальной траектории, соединяющей два индивидуальных лица.
Рисунок 9. Ответы одиночного нейрона (C99-117) нижневисочной коры обезьяны C99 на стимуляцию лицами с разной степенью индивидуальности или карикатуризации. На каждой панели показан стимул, растерные диаграммы и перистимульные гистограммы ответов на предъявление этого стимула. Длительность экспозиции лица составляла 400 мсек (черная горизонтальная линия на перистимульных гистограммах). Четыре ряда графиков соответствуют радиальным осям карикатуризации в стимульном пространстве для индивидуальных лиц F1-F4. Уровень индивидуальности меняется от 0% (слева, идентичен в каждом случае) по направлению к полной индивидуальности (100%) вплоть до 160 % (справа, карикатура).
Пример, приведенный на рис. 9, иллюстрирует типичный эффект влияния разных уровней индивидуализации лиц на активность нейронов-детекторов лиц. В данном случае для каждого тестируемого лица использовали девять уровней индивидуализации, варьирующих от 0% (усредненное лицо) до 100% (индивидуальное лицо), и далее до 160% (карикатура). Этот нейрон, как и многие другие, демонстрировал практически линейное возрастание частоты разрядов в зависимости от уровня индивидуальности. При этом нейрон четко сигнализировал даже о незначительных изменениях в уровне индивидуальности лица. Интересен тот факт, что изменения частоты разряда нейрона отражали разницу между стимулами даже у лиц, уровень индивидуальности которых лежал близко к пороговому значению по поведенческому критерию (см. постстимульную гистограмму, полученную при усреднении ответов нейрона в ответ на предъявление лица с уровнем индивидуальности 20%; рис. 9, верхний ряд графиков модификации лица F1).

Рисунок 10. Кривые настройки нейронов-детекторов лиц. Слева более крупно даны примеры активности нейронов, один из которых отвечал усилением активности на предъявление стимулов с высоким уровнем индивидуальности. Второй нейрон был более чувствителен к усредненному лицу. Заключенные в ярко-оранжевые и ярко-зеленые рамки кривые настойки принадлежат нейронам, записанным в экспериментах на обезьяне N97, тогда как оставшиеся кривые – обезьяне C99. Разные цвета кривых соответствуют разным наборам морфов индивудуальных лиц.
При анализе кривых настроек нейронов-детекторов лиц выяснилось, что для большей части клеток характерно монотонное увеличение частоты разряда по мере смещения стимула по шкале индивидуальности в направлении ее усиления. На рисунке 10 представлены примеры активностей 10 нейронов-детекторов лиц передней нижневисочной коры. Как правило, тестирование нейронов проводили с ограниченным набором видоизменных лиц: уровни индивидуальности были заданы положительными значениями на траектории индивидуальности. Для построения кривых настроек для отдельных нейронов комбинировали усредненные данные двух наборов стимулов-лиц. Это было сделано для того, чтобы получить кривые настройки классического вида. Таким образом, каждому набору стимулов соответствует участок кривой настройки определенного цвета. Отрицательные значения по осям абсцисс, по которым отложены значения уровня индивидуальностей, на самом деле не являются таковыми: они лишь лежат на другой оси карикатуризации. Нулевое значение уровня индивидуальности соответствует усредненному лицу.
Полученные данные демонстрируют, что в популяции нейронов-детекторов лиц существуют, по крайней мере, два типа нейронов. Один тип нервных клеток обладает повышенной чувствительностью к прототипу, т.е. частота разряда таких нeйронов максимальна при предъявлении усредненного лица. Другой тип клеток напротив настроен на детектирование различий между прототипом (усредненным лицом) и индивидуальными лицами.
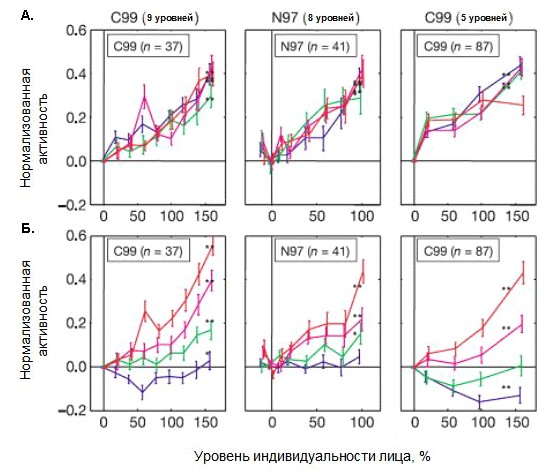
Рисунок 11. Зависимость популяционных ответов нейронов от уровня карикатуризации лица. А. популяционные ответы для каждой обезьяны (в некоторых экспериментах с обеьзьяной С99 использовали 5 уровней карикатуризации, в других - 9) для всех четырех использованных индивидуальных лиц (кривые разного цвета), различных для каждой обезьяны. Б. лица упорядочены по рангам от вызывающих минимальный до вызывающих максимальный ответ. Ответы усредняли отдельно для каждого ранга и уровня карикатуризации (приведены средние значения и ошибки среднего). Звездочками обозначены кривые демонстирурющие достоверный тренд (*р = 0.05; **р = 0.01).
Популяционные данные по кодирующим свойствам нейронов-детекторов лиц в обобщенном виде показаны на рис. 11. Для получения усредненных графиков для четырех наборов стимулов (рис. 11А), расположенных на соответствующих осях индивидуальности, данные нормировали отдельно для каждой траектории. При проведении нормировки сначала из ответа на определенное лицо набора вычитали среднюю активность на лицо с уровнем индивидуальности 0%. Затем определяли максимальный ответ нейрона среди реакций на данный набор стимулов и делили полученные после вычитания значения на максимумальное. После чего данные по всем нейронам можно было суммировать для одной траектории индивидуальности. Здесь, в первую очередь, следует отметить факт, что активность нейронов-детекторов находится практически в линейной зависимости от уровня карикатуризации лица. Такого рода монотонность наблюдается вне зависимости от того, сколько уровней карикатуризации было использовано. Следовательно, одним из характеристических признаков нейронов-детекторов лиц является способность монотонно усиливать активность по мере увеличения индивидуальных черт определенного лица.
Полученные экспериментальные данные показывают, что ось карикатуризации является первичной определяющей для частоты разряда нейронов нижневисочной коры, селективных к лицам. Специальная роль усредненного лица в формировании настройки нейронов в данной области коры мозга может отражать способность системы выявлять естественные вариации в характеристических признаках лиц.
На основании полученных данных можно предположить, что настройка нейронов передней части нижневисочной коры может быть сформирована под влиянием статистики реальных образов зрительного мира. Поэтому возможно, что наблюдаемые нами настройки являются результатом использования данного набора стимулов. Конечно, практически невозможно отделить характер настройки этих нейронов от таких параметров как степень знакомства со стимулом или определенность ответа. Данные, полученные нами на двух обезьянах в двух отличающихся друг от друга экспериментах, свидетельствуют против влияния данных факторов на нейронные ответы.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Адаптивность нейронного представления: функциональная анатомия первичной зрительной коры кошки
В острых опытах на анестезированных и обездвиженных кошках с помощью оптического картирования поля 17 мы не обнаружили достоверного различия в относительной тангенциальной площади ориентационных колонок, активированных решётками базисных и диагональных ориентаций. Таким образом, наши экспериментальные данные не подтвердили «эффекта промежуточных ориентаций», т.е. увеличенного представительства в первичной зрительной коре кошки детекторов вертикали и горизонтали.
Теоретически в ходе постнатального онтогенеза мозга на любой участок сетчатки глаза могут проецироваться элементы контуров любых ориентаций, поэтому логично предположить, что все ориентации должны быть представлены в зрительной коре в равной степени (Coppola & White, 2004; Purves et al., 2002). С другой стороны, считается (Coppola et al, 1998a), что ориентационная анизотропия отражает преобладание в зрительном мире линий двух базисных ориентаций – горизонтальной и вертикальной. Действительно, анализ цифровых фотографий показывает, что наш зрительный мир анизотропен – в нём преобладают контуры, ориентированные горизонтально и вертикально (Coppola et al, 1998б). Исследования на нескольких видах млекопитающих с использованием различных методов подтверждают, что анизотропия в корковом представительства ориентационной чувствительности может быть общей чертой корковой зрительной организации, во всяком случае, у хищных и приматов (De Valois, 1982). Итак, существующие в литературе свидетельства в пользу исходной гипотезы не вызывают сомнения в их достоверности (Appelle, 1972; Li et al., 2003).
В чём же причина различия наших результатов и данных литературы? Можно перечислить и обсудить комплекс факторов, возможно, объясняющих эти различия. Это, во-первых, особенности метода, определяющего критерий оценки активации элементов коры. Во-вторых, это – расхождения в протоколе эксперимента (например. виде экспериментального животного и типе используемого анестетика). В-третьих, это - особенности зрительного опыта в онтогенезе экспериментальных животных. Остановимся на этих обстоятельствах подробнее.
Начнём с метода исследования. Особенность нейронных исследований заключается в том, что микроэлектрод при погружении в кору, как правило, избирательно выделяет активность сравнительно крупных нейронов. В отличие от этого использованный нами в качестве критерия внутренний оптический сигнал коры имеет популяционную природу и в равной мере отражает вклад в её активацию всех клеточных элементов данной области коры. Оптическое картирование позволяет проводить длительное исследование пространственной упаковки разных функциональных популяций нейронов коры, т.е. тангенциального распределения её нейронных модулей.
Нельзя не отметить, что оптическим методом нейронная активность картируется только в верхних слоях коры (700-800 мкм). В них сконцентрированы в основном нейроны с торцевым торможением в рецептивном поле (2- и 3-й слои) и поля простого типа (верхние подслои 4-го слоя). Можно было бы полагать, что их ориентационная настройка изотропна в отличие от нейронов 5- и 6-го слоёв со сложными рецептивными полями. Однако этому противоречат убедительные электрофизиологические данные, полученные на 4418 нейронах, о выраженной анизотропии в субпопуляции простых, а не сложных клеток (Li et al., 2003). Таким образом, противоречие между этими данными и результатом нашего оптического картирования не объясняется и не снимается.
Второе обстоятельство – вид животного и особенности протокола эксперимента. В литературе в основном описаны данные, полученные на хорьках. Эффект анизотропии исследован лишь в одной работе с оптическим картированием зрительной коры кошки (Wang et al., 2003). Что касается протокола эксперимента, то ранее мы обнаружили роль вида и уровня наркоза в оценке данных, полученных оптическим картированием (Шевелев и др., 2005). Так, в части опытов при использовании ингаляционного наркоза (галотан с закисью азота и с кислородом) в коре кошки обнаружены не только классические ориентационные колонки, но и реакции на пересечения линий. В отличие от этого при внутривенном введении тиопентала (более глубокий наркоз) выявляет лишь ориентационные эффекты.
Интересной нам представляется и третья возможная причина различия наших результатов и данных литературы – особенность зрительного опыта в онтогенезе экспериментальных животных. Как было показано на хорьках, эффект анизотропии сильнее всего проявляется на ранних стадиях онтогенеза, а в процессе накопления зрительного опыта он в значительной мере нивелируется. Существенно, что во всех работах, где выявлен «эффект промежуточных ориентаций», использовали животных, рождённых и выращенных в виварии для экспериментальных целей. Можно полагать, что обеднённая зрительная среда вивария в процессе выращивания таких животных определяет превалирование базисных ориентаций. Хорошо известно, что особенности среды, т.е. зрительный опыт, значительно влияют на развитие связей нейронов зрительной коры и на характер представительства в ней ориентационной чувствительности (Chapman & Bonhoeffer, 1998; Coppola & White, 2004; Imbert, 1979). Важно, что в коре зрительно депривироваnных хорьков ориентационная анизотропия выражена значительно сильнее, чем в нормальных условиях. Поэтому авторы полагают, что нормальный зрительный опыт постепенно обеспечивает изотропное представление в коре ориентационной чувствительности (Coppola & White, 2004).
Таким образом, в наших экспериментах не удалось обнаружить ориентационной анизотропии в первичной зрительной коре кошки. Предположительно, такой результат может быть связан с особенностями индивидуального зрительного опыта животных. Однако, напрямую это предположение не было проверено. В будущем мы планируем проверить данную гипотезу в прямых экспериментах с изменяемыми условиями зрительной среды.
Стабильность кодирующих свойств нейронов-детекторов.
Долговременная регистрация активности одиночных нейронов. Одной из целей нашей работы было создание методики, которая позволила бы обеспечить эффективное и, по возможности, наименее травмирующее для животного накопление экспериментальных данных. Хронически имплантированные микроэлектроды имеют ряд преимуществ перед обычными методиками регистрации нервных клеток в мозге бодрствующих животных. В первую очередь, использование данной методики позволяет одновременно наблюдать за активностью нескольких нейронов длительное время. В такой ситуации становится возможным получение качественно нового типа нейронных данных: данных о кооперативных взаимодействиях между нейронами. Это особенно важно, поскольку в последнее время наметилась тенденция к предпочтению теории кодирования информации нейронными ансамблями. Таким образом, именно активности групп нейронов отводится центральная роль в процессах обработки и хранения информации в мозге. И в данном случае не важно, каким на самом деле является нейронный код (популяционным или заданным синхронной активностью), лишь множественные электроды позволяют наблюдать за активностью нескольких клеток одновременно. При стабильных отведениях нейронной активности появляется возможность проследить за изменениями параметров кооперативной деятельности во времени.
Не менее важным является то обстоятельство, что хронические имплантации множественных электродов позволяют уменьшить количество экспериментальных животных без ухудшения количества и качества получаемых экспериментальных данных. Сочетание стабильности регистраций и возможности коррекции положения пучка электродов в мозге открывают новые возможности перед экспериментатором. Так, он может чередовать периоды длительного наблюдения за активностью группы нейронов с моментами перемещения кончиков электродов и переходами к отведениям новых групп нервных клеток. В случае одного из наших последних имплантатов, в котором пучок электродов был закреплен в микроманипуляторе, по крайней мере в течение года были возможны регистрации клеточного сигнала хорошего качества. За это время в экспериментах было накоплена информация об активности 92-х нейронов, за работой 45 клеток из которых мы могли наблюдать более чем в одном эксперименте.
Таким образом, в данной работе было показано, что с помощью нашей конструкции электродного пучка возможны регистрации отдельных нервных клеток в течение длительного времени. Из литературных данных известно, что качество регистрации активности с помощью имплантированных на поверхности мозга электродов ухудшается уже через 3-4 недели (Legendy, 1984). В случае ухудшения качества регистраций некоторые авторы рекомендуют применять микростимуляцию через имплантированные электроды (Schmidt et al., 1993). По всей видимости, имеются различия в поверхностных и глубинных имплантациях микроэлектродов, поскольку некоторые авторы описывают ситуации успешных регистраций клеточной активности на протяжении 8-15 месяцев после имплантации (Nikolelis, 1996). Такие различия можно объяснить более массивным иммунным ответом, который наблюдается на поверхности мозга, поскольку в такой ситуации затронутой оказывается и твердая оболочка мозга, через которую вводят электроды. В случае глубинных имплантаций или при введении электродов со стороны белого вещества мозга травматическое механическое воздействие на твердую и мягкую оболочки мозга минимизируется, уменьшается вероятность повреждения кровеносных сосудов и в целом практически не оказывается воздействий на непосредственное место имплантации. Все это позволяет сохранить метаболизм данной области мозга в сохранности.
Использование тонких микропроволок и жесткого закрепления их в несущей трубочке придает нашей конструкции способность к отслеживанию микродвижений мозга, что обеспечивает стабильность отведения клеточной активности. В рамках данной работы удалось выработать основанные на параметрах регистрируемого от нейрона сигнала критерии, которые позволяют судить о стабильности отведения клеточной активности. В качестве таких параметров мы использовали амплитудные характеристики спайка, форму потенциала действия и распределения межспайковых интервалов. В случае стабильной регистрации сигнала от одного и того же нейрона эти характеристики остаются практически неизменными. Так изменения амплитуды спайка составляют около 20%. По литературным данным (Gray, 1995) 40% процентное изменение амплитуды спайка возможно при смещении кончика электродов на 15 м относительно тела клетки. Таким, образом в нашем случае сдвиг электродов возможен лишь в пределах < 15 м, что и обеспечивает стабильность регистраций. Несмотря на то, что все использованные нами для оценки стабильности регистраций критерии более надежным является одновременное их применение.
Немаловажной составляющей успеха хронических имплантаций является использование адекватного метода закрепления имплантата на поверхности черепа животного. При развитии имплантатов для медицины были получены важные данные по взаимодействиям между имплантатом и организмом-донором. Так, имплантаты могут оказывать на организм-донор токсическое, биоактивное или инертное действие (Wintermatel und Ha, 1996). В случае токсического действия в окружении имплантата появляется большое количество мертвых клеток, что вызывает массивный имунный ответ организма и ведет к отторжению конструкции. При биоактивном воздействии между имплантатом и тканью образуются специфические связи, обеспечивающие успешность вживления конструкции в организм. В случае инертных взаимодействий вокруг имплантата образуется соединительнотканная оболочка, обладающая низкой адгезивной способностью. При закреплении электродов с помощью акрилового цемента мы имели дело с токсическим воздействием на ткань. Из литературы известно, что уже через три месяца после имплантации происходит отторжение закрепляющей конструкции (Pfingst et al., 1989). Вероятно, решающую роль при отравляющем воздействии на кость играют как высокотоксические жидкие компоненты акрилового цемента, так и локальное тепловое воздействие при затвердевании акрилата (Albrektsson und Linder, 1984).
Идеальным является использование таких материалов для закрепляющей электроды конструкции, которые обеспечили бы ее надежное соединение с костной тканью. В случае обезьяны N97 сочетание идеально подходящего к поверхности черепа имплантата и использование специального напыления позволили добиться стабильного закрепления имплантата на кости. В дальнейшем именно такие конструкции следует использовать для удержания регистрирующих электродов в стабильной позиции.
Таким образом, нам удалось разработать метод стабильной регистрации активности от отдельного нейрона, который позволяет получать принципиально новые данные о работе нейронных популяций. Комплексный подход делает этот метод уникальным и крайне полезным для исследования свойств нейронных представлений в мозге.
Стабильность представления сложных зрительных объектов в нижневисочной коре обезьян. Одним из главных свойств нервной системы животных является ее способность к обучению. Пластичность нервной системы служит залогом высокой приспособляемости организма к постоянно изменяющимся условиям окружающей среды. Однако, лишь сочетание пластичности и стабильности делает адаптивное поведение животных по-настоящему успешным.
В основе теории сенсорного восприятия лежит понятие внутреннего представления: нейронного отображения объектов и событий внешнего мира. Предполагается, что нейроны головного мозга кодируют информацию об объектах внешнего мира с помощью изменения частоты разряда. Сохранность определенного паттерна импульсации во времени обеспечивает стабильность внутреннего представления, что, в свою очередь, является важной предпосылакой для формирования паттернов адаптивного поведения.
До настоящего времени исследование стабильности нейронного представления были затруднены. Затруднения носили преимущественно технический характер. С одной стороны, необходимо было разработать методы стабильной регистрации клеточного сигнала в мозге, а с другой нужно было выявить критерии оценки стабильности таких отведений. В данной работе нам удалось добиться прогресса в этом направлении. В результате стали возможными долговременные наблюдения за активностью отдельного нейрона.
В ходе наших экспериментов в первую очередь было обнаружено, что нейроны нижневисочной коры, как правило, демонстрируют широкий спектр предпочтений по отношению к зрительным стимулам, которые были использованы в наших экспериментах. Эти данные хорошо согласуются с теорией «экономного кодирования» (Rolls, 1986). В соответствии с данной теорией отдельная нервная клетка может принимать участие в работе разных нейронных ансамблей, которые совокупной активностью кодируют тот либо иной объект внешнего мира. В последнее время появляется все больше подтверждений данной теории, однако вопрос о принципах кодирования остается по-прежнему открытым. Так, продолжается дискуссия о том, какие параметры нейронного разряда являются решающими для кодирования: средняя частота импульсации нейрона, временные модуляции активности или синхронизованная активность нескольких нейронов.
Предшествующие работы показали, что временные модуляции ответа, особенно в последней его фазе, у нейронов нижневисочной коры несут информацию об идентичности зрительного стимула (Sugase et al., 1999). В соответствии с этим, мы обнаружили, что первая фазическая часть ответа нейронов нижневисочной коры часто присутствовала в реакциях клетки на большую часть зрительных стимулов. Любопытно отметить, что в одной из последних работ, касающейся функций слуховой коры высокого уровня, было показано, что только оптимальный для данного нейрона стимул способен вызывать более позднюю тоническую фазу ответа. Можно предположить, что и в зрительной, и в слуховой коре нейроны в начале ответа демонстрируют селективность скорее к относительно широкому диапазону стимулов. После этого популяционные взаимодействия нейронов обеспечивают селективность к небольшому набору внешних воздействий. В настоящей работе, поздняя стадия ответов нейронов часто включала в себя периоды торможения активности, осцилляторных циклов и off-ответы. Таким образом, понятие селективности может обозначать не только определенную частоту разряда в ответ на определенную стимуляцию, но может характеризоваться и тонкими перестройками возбудительно-тормозных паттернов разряда, формируемыми в локальных корковых сетях.
Общепринято, что нейроны нижневисочной коры изменяют характер активности в ответ на предъявление биологически значимых стимулов, что является прямым следствием процессов обучения в данной области. Ответ нейрона может быть усилен или ослаблен в течение одного экспериментального дня (Messinger et al., 2001; Jagadeesh et al., 2001; Erickson & Desimone, 1999) или более длительного периода времени (Leopold et al., 2006; Kiani et al., 2007; Li & DiCarlo, 2008; Baker et al., 2002; Kayaert et al., 2005). Вероятно, такие модификации находятся под влиянием локальных тормозных нейронов, которые, по-видимому, формируют силу и характер временных модуляций селективных ответов (Wang et al., 2000). Устойчивость нейронной специфичности во времени, однако, не была показана напрямую, а эффекты обучения чаще всего демонстрировались на уровне популяций нейронов.
Настоящее исследование доказывает, что селективность индивидуальных нейронов может быть стабильна в течение нескольких дней и даже недель. При этом сохраняется как общий профиль реакций на набор тестовых стимулов, так и тонкие временные модуляции разряда нейронов. Можно предположить, что однажды сформированная чувствительность нейрона к зрительным стимулам остается неизменной, по крайней мере до тех пор, пока не возникает дополнительного обучающего воздействия. Это делает подход к изучению индивидуальных функциональных свойств нейронов в течение длительных промежутков времени с помощью хронически имплантированных электродов весьма актуальным для понимания формирования нейронной чувствительности в процессе накопления зрительного опыта.
Роль прототипа в кодировании индивидуальных лиц нейронами нижневисочной коры обезьян
В соответствии с теорией Д.Марра, завершающей стадией процесса распознавания является категоризация, которая позволяет определить биологический смысл зрительных объектов. Прототип является одним из центральных понятий в некоторых теориях категоризации зрительных объектов (Palmer, 1999). Он представляет собой идеальный образец, вокруг которого группируются все остальные члены той либо иной общности объектов. Таким образом, прототип - наиболее типичный (усредненный) представитель категории. По всей видимости мозг способен формировать для разных категорий объектов отдельные прототипы.
До настоящего времени исследование роли прототипа в процессах восприятия было затруднено. Развитие компьютерной техники и графики позволило разработать целый ряд моделей морфинга зрительный стимулов, центральным звеном в которых является усредненный образец – прототип. В нашей работе были использованы наборы стимулов лиц, у которых уровень индивидуальности мог плавно изменяться от 0 до 160%. Нулевым уровнем индивидуальных черт обладает прототип, увеличение уровня индивидуальности усиливает индивидуальные черты лица вплоть до его карикатуризации. В психофизических экспериментах (Leopold, 2001) были получены данные, демонстрирующие важность прототипа для кодирования информации об индивидуальных лицах.
Эксперименты по психофизическому тестированию распознавания лиц показали принципиальный характер сходства этого процесса у обезьян и человека. Монотонное увеличение уровня индивидуальности лица вызывало улучшение распознавания лиц как обезьянами, так и человеком. Разный угол наклона психометрической функции (у обезьян он был меньше) может свидетельствовать о том, что обезьяны распознают человеческие лица хуже. Возможно, что использование лиц обезьян дало бы другой результат. Тем не менее, эксперимент с адаптацией показал, что и у обезьян кодирование лиц построено, по всей видимости, по оппонентному принципу. Специфическая адаптация к «антилицу» вызывала положительные сдвиги порогов восприятия. Психофизические эксперименты заложили основу последующей нейронной части работы, которую проводили с наборами стимулов-лиц, которые в поведенческих опытах дали положительный результат.
В экспериментах по регистрации активности нейронов детекторов-лиц обнаружено, что настройка нейронов передней нижневисочной коры часто была центрирована относительно усредненного, неопределенного в плане идентичности лица. При этом наблюдали два типа ответов нейронов: а) монотонное увеличение частоты разряда нейрона по мере увеличения уровня индивидуальности лица; б) усиление импульсации в ответ на предъявление усредненного лица – прототипа. Интересно, что кривые настройки нейронов не отличались у двух обезьян, хотя одна из них освоила задачу по активному распознаванию лица, а вторая – нет. Таким образом, можно выдвинуть предположение, что обучение не оказывает прямого воздействия на способность нейронов кодировать информацию об индивидуальных характеристиках лиц.
В данных экспериментах было получено подтверждение в пользу гипотезы «экономичного кодирования». Как правило, нейроны-детекторы лиц фиксировали изменения уровня индивидуальности нескольких лиц одновременно. Поэтому по-прежнему остается открытым вопрос о характеристиках популяционного кода, которые дают возможность однозначно и точно опознавать индивидуальные лица.
Полученные данные позволяют выдвинуть предположение, что характеристики ответов нейронов-детекторов лиц отображают степень различия между информацией о лице индивидуума с хранящимся в памяти образцом усредненного лица, т.е. прототипом. Результаты нейрофизиологических экспериментов связывают ответы нейронов нижневисочной коры и психологические модели различения индивидуальных лиц.
Подводя итоги нашего исследования способов кодирования информации о лицах, можно предположить, что селективные к лицам ответы нейронов могут представлять собой результат внутреннего процесса сравнения текущей и хранящейся в памяти информации, который позволяет мозгу выявлять незначительные различия в форме индивидуальных лиц, что формирует основу для процесса их распознавания. В этом контексте сравнение зрительного образа с неким выработанным ранее прототипом или нормой обеспечивает высокий уровень надежности в работе зрительного анализатора в условиях постоянной трансформации образа, который типичен для сенсорного восприятия.
Наиболее частые трансформации зрительного объекта, такие как изменения в размере изображения, его ракурсе, эмоциональном выражении лица и возрасте субъекта, не обязательно должны быть заучены и приложены для каждой идентичности. Стандартные алгоритмы сравнения с образцом могут обеспечить экономию времени, необходимого для обучения инвариантному восприятию индивидуального лица. Кроме того экономия может затронуть также мощности мозга, задействованные в хранении информации. Механизм опознания, базирующийся на сравнении с прототипом, как нам кажется, может объяснить также, почему процесс опознания лиц происходит настолько быстро, легко и, как правило, подсознательно.
ВЫВОДЫ
1. Оптическое картирование по внутреннему сигналу позволяет быстро и эффективно тестировать особенности функциональной анатомии зрительной коры, связанные с активностью нейронов-детекторов.
2. Функциональные модули, осуществляющие анализ информации о базисных и наклонных ориентациях элементов изображений, занимают равные по площади поверхности первичной зрительной коры.
3. Использование хронически имплантированных микроэлектродов и адекватных критериев оценки параметров нейронной активности позволяют обнаружить и исследовать стабильность селективных свойств нейронов-детекторов.
4. В условиях отсутствия внешних воздействий, вызывающих изменения характеристических нейронных свойств, корковые детекторы способны длительное время сохранять стимул-специфический паттерн активности.
5. Кодирование сложных изображений может осуществляться как за счет увеличения частоты разряда нейронов, так и за счет временных модуляций импульсации клетки.
6. На поведенческом уровне системы распознавания лиц человека и обезьяны принципиально похожи. По всей видимости в основе распознавания лежит принцип сравнения с прототипом.
7. Нейроны-детекторы кодируют информацию об индивидуальных лицах с помощью монотонных изменений частоты импульсации, при этом «нулевая частота» разряда определяется активностью на прототип. Кодирование информации с помощью прототипа может быть одним из универсальных принципов работы сенсорных систем мозга.
СПИСОК СТАТЕЙ ПО ТЕМЕ ДИССЕРТАЦИИ
По теме диссертации опубликовано 9 статей, из них 4 в отечественных и 5 в рецензируемых международных изданиях.
Статьи в рецензируемых научных журналах:
- Иванов Р.С., Лямзин Д.Р., Бондарь И.В., Куликов М.А., Шевелев И.А. Перекрытие зон оптических ответов на крестообразную фигуру и отрезки линий разной ориентации в первичной зрительной коре кошки. Журн. ВНД им. И.П. Павлова, 2010, 60 (2):133-137.
- Bondar IV, Leopold DA, Richmond BJ, Victor JD, Logothetis NK. Long-term stability of visual pattern selective responses of monkey temporal lobe neurons. PLoS One. 2009 Dec 9; 4(12):e8222.
- Лямзин Д.Р., Бондарь И.В., Иванов Р.С. Обработка функциональных карт зрительной коры мозга кошки, полученных по внутреннему оптическому сигналу. Росс. Физиол. Журн. им. И.М. Сеченова, 2009, 95(4): 335-346.
Lyamzin DR, Bondar IV, Ivanov RS. Processing of functional maps of the cat visual cortex obtained using intrinsic optical signals. Neurosci Behav Physiol. 2010 Jun; 40(5):573-81.
- D.A. Leopold, I.V. Bondar, M.A. Giese. Norm-based face encoding by single neurons in the monkey inferotemporal cortex. Nature. 2006 Aug 3;442(7102):572-5.
- Иванов Р.С., Бондарь И.В., Салтыков К.А., Шевелев И.А. Площадь зон оптической активации поля 17 коры мозга кошки при предъявлении решёток разной ориентации. Журн. ВНД им. И.П. Павлова, 2006, 56(4): 516-522.
- Академик Шевелёв И.А., Бондарь И.В., Айзел У., Кисварди З., Бузас П., Иванов Р.С., Салтыков К.А. Наркоз и тангенциальная упаковка в стриарной коре кошки нейронов с чувствительностью к крестообразным фигурам. // Доклады Академии наук, 2005, 402(4):566–570
- J. Kruger, V.A. Lamme, I. Bondar. Colour-related oscillations in the striate cortex of awake monkeys: "reverse" observations. Experimental Brain Research. 139(2):180-7; 2001
- I. Porada, I.V. Bondar, W.B. Spatz and J. Kruger. Rabbit and monkey visual cortex: more than a year of recording with up to 64 microelelctrodes. Journal of Neuroscience Methods. 95(1): 13-28; 2000.
Главы в книгах:
D. A. Leopold & I. Bondar. Adaptation to Complex Visual Patterns in Humans and Monkeys. In: Fitting the Mind to the World Adaptation and After-Effects in High-Level Vision. Eds. Colin W. G. Clifford and Gillian Rhodes, Oxford University Press, 2005
J. Krger, H. Haan, I. Bondar. Recording from foveal striate cortex while the monkey is looking at the stimulus. In Neural Networks: Artificial Intelligence and Industrial Applications Eds. B. Kappen and S. Gielen, Springer, Berlin. pp 33-36, 1995.
Материалы конференций:
И.В. Бондарь, Р.С. Иванов Исследование популяционной активности зрительной коры млекопитающих с помощью оптического картирования по внутреннему сигналу: чувствительность к контрасту, пространственной частоте и ориентации стимула.— IX международная конференция «Прикладная оптика - 2010.» Санкт-Петербург 19-22 октября 2010 г.
И.В. Бондарь Особенности специализации нейронов-детекторов нижней височной коры обезьяны: стимул-специфические модуляции частоты разряда и стабильность профилей селективности. XXI Съезд Физиологического общества имени И.П. Павлова. Калуга.
19-25 сентября 2010 г.
Е.Е. Минакова, Р.С. Иванов, И.В. Бондарь Изучение ретинотопической организации зрительной коры мозга крыс с помощью метода оптического картирования по внутреннему сигналу. Научная сессия НИЯУ МИФИ-2010. Нанофизика и нанотехнологии. Фундаментальные проблемы науки. Москва. 25-31 января 2010 г.
Р.С. Иванов, Д.Р. Лямзин, И.В. Бондарь Паттерн оптической активации нейронов первичной зрительной коры при стимуляции крестообразными фигурами и их составляющими (метод оптического картирования по внутреннему сигналу). ICNC-2009. XV Международная конференция по нейрокибернетике. Ростов-на-Дону. 23-25 сентября 2009 г.
Р.С. Иванов, Д.М. Лямзин, И.В. Бондарь Паттерн оптической активации нейронов первичной зрительной коры при стимуляции крестообразными фигурами и их составляющими.–– ISIR-2008, Научные труды II Съезда физиологов СНГ «Физиология и здоровье человека». Кишинэу, Молдова, 29-31 октября 2008 года. М.-Кишинэу: Медицина–Здоровье, 2008, с. 113-114.
I.V. Bondar, D.A. Leopold, N.K. Logothetis Stability of neuronal representation in inferotemporal cortex: longterm single neurons study by chronically implanted microelectrodes.
Материалы Международного конгресса по психофизиологии (С.Петербург, сентябрь 2008 г.).
Иванов Р.С., Бондарь И.В., Салтыков К.А., Шевелев И.А. Особенности функциональных карт ориентационной чувствительности по данным оптического картирования первичной зрительной коры кошки. Материалы 14-ой международной конференции по нейрокибернетике. Ростов-на-Дону. Т. 1., с. 28-30. Сентябрь 2005
Иванов Р.С., Бондарь И.В., Салтыков К.А., Шевелев И.А. Особенности функциональных карт ориентационной чувствительности по данным оптического картирования первичной зрительной коры кошки. Научные труды 1 Съезда физиологов СНГ. Сочи, Дагомыс, Т. 1, C. 62-63. Август 2005
И.В. Бондарь, И.А. Шевелев, К.А. Салтыков, Р.С. Иванов, А.С. Тихомиров Исследование популяций нейронов первичной зрительной коры кошки методом оптического картирования. Рос. Физиол. журн. им. И.М.Сеченова, т. 90, № 8, стр. 298, 2004.
I.V. Bondar, D.A. Leopold, A.A. Boldyrev, N.K. Logothetis. Long-term stability of complex neural responses in monkey inferotemporal cortex. Society for Neuroscience Abstracts, 2003.
D.A. Leopold, I.V. Bondar, G. Mahlknecht, N.K. Logothetis. Prototype-referenced encoding of faces in the monkey inferotemporal cortex. Society for Neuroscience Abstracts, 2003.
I.V. Bondar, D.A. Leopold and N.K. Logothetis. Evidence for prototype-referenced encoding of faces in the monkey. 3rd Forum of European Neurosciеnce, Paris, 2002
I.V. Bondar, D.A. Leopold, J.M. Pauls, N.K. Logothetis. Neural responses related to face identity in inferotemporal cortex of monkeys measured with 64 implanted electrodes. Society for Neuroscience Abstracts, 2001
I.V. Bondar, N.K. Logothetis. Activity in visual cortex measured with 64 chronically implanted electrodes in monkeys. Society for Neuroscience Abstracts, 2000
I. Bondar, J. Stiller, J. Krger. Recognizing saccades from local field potentials in monkey visual cortex Proceedings of the 26th Gttingen Neurobiology Conference 1998, vol II, p. 482
J. Krger, I. Bondar, M. Fahle, E. Schottmann, X. Schwiete. Kontrollierte Blickrichtung bei wachen Affen ohne Dressur Visuelle Wahrnehmung: Beitrge zur 1. Tbinger Wahrnehmungskonferenz, s. 164
H. Haan, J. Timmer, I. Bondar, J. Krger. Local field potentials (LFPS) and spikes in monkey striate cortex Proceedings of the 24th Gttingen Neurobiology Conference 1996, vol II, p. 394
J. Krger, I. Bondar, B. Reif. Eye movements and multielectrode recordings in an awake monkey Proceedings of the 24th Gttingen Neurobiology Conference 1996, vol II, p. 396
J. Krger, I. Bondar, I. Porada, B. Reif. High-frequency oscillatioons in monkey foveal striate cortex Proceedings of the 23rd Gttingen Neurobiology Conference 1995, vol II, p. 474