

Тоническая регуляция моторных реакций
На правах рукописи
САФРОНОВ
Вадим Александрович
ТОНИЧЕСКАЯ РЕГУЛЯЦИЯ
МОТОРНЫХ РЕАКЦИЙ
Специальность: 03.03.01 - физиология
Автореферат
диссертации на соискание ученой степени
доктора биологических наук
Москва - 2012
Работа выполнена в Федеральном государственном бюджетном учреждении - Научно-исследовательский Институт нейрохирургии им. акад. Н.Н. Бурденко Российской академии медицинских наук
Научный Козловская Инеса Бенедиктовна
консультант: д.м.н., профессор, член-корреспондент РАН, заведую-
щая отделом сенсомоторной физиологии и профилактики
Федерального государственного учреждения науки Государ-
ственный научный центр Российской Федерации - Институт
медико-биологических проблем Российской академии наук
Официальные Шульговский Валерий Викторович
оппоненты: д.б.н., профессор, заведующий кафедрой высшей нерв-
ной деятельности, руководитель лаборатории системно-
го анализа деятельности мозга Биологического факуль-
тета Московского государственного университета имени
М.В. Ломоносова
Сонькин Валентин Дмитриевич
д.б.н., профессор, главный научный сотрудник лабора-
тории мышечной деятельности и физического воспита-
ния Федерального научного учреждения - Институт воз-
растной физиологии Российской академии образования
Фролов Александр Алексеевич
д.б.н., профессор, заведующий лабораторией математи-
чаской нейробиологии обучения Федерального государ-
ственого бюджетного учреждения науки - Институт Выс-
шей нервной деятельности и нейрофизиологии Россий-
ской академии наук
Ведущая Федеральное государственное бюджетное учреж-
организация: дение науки - Институт физиологии им. И.П. Пав-
лова Российской академии наук (Санкт - Петер-
бург)
Защита диссертации состоится « 21 » ноября 2012 года в «10-00» часов на заседании диссертационного совета Д 002.111.01 в Федеральном государственном бюджетном учреждении науки Государственном научном центре Российской Федерации - Институте медико-биологических проблем Российской академии наук по адресу: 123 007, Москва, Хорошевское шоссе, д. 76 А.
С диссертацией можно ознакомиться в библиотеке ГНЦ РФ – ИМБП РАН
Автореферат разослан « » _______ 2012 года
Ученый секретарь диссертационного совета,
доктор биологических наук М.А. Левинских
Актуальность проблемы
Мышечный тонус является важным фактором в организации моторики тела. Локомоции и поддержание позы при выполнении движений обес-печиваются тонической активностью большого числа скелетных мышц. На существенную значимость тонических механизмов для координированного выполнения моторных задач указывал Н.А. Бернштейн. Нарушения мышечного тонуса у неврологических больных коррелируют с такими моторными дисфункциями, как гиперкинезы, ригидность, спастика, дискоординация походки, нарушение статики. Изменения мышечного тонуса имеют место и в условиях гипогравитации (И.Б. Козловская и соавт., 1984; А.И. Григорьев и соавт., 2004).
Понимание механизмов движений, начиная с работ Шеррингтона, базируется на результатах многочисленных исследований произвольного и рефлекторного управления моторными процессами. Однако один из важнейших тонических механизмов - реакции укорочения, поддерживающей напряжение мышц при пассивном сближении их противоположных концов, оставался необъясненным. О механизме реакции укорочения в научной литературе имеется сравнительно мало данных, а имеющиеся - в большой степени противоречивы.
На основании результатов экспериментальных исследований Ron-dot и Matral (1973) заключили, что реакция укорочения (РУ) является аутогенной, что Iа-афференты не принимают участия в её генерации, как не принимают участия в вызове реакции и Ib–афференты антагониста, сомнительно также участие в этих процессах афферентов группы II. Однако, описав основные свойства РУ, исследователи не смогли предложить механизм этой реакции, что является чрезвычайно важным как для понимания процессов регуляции мышечного тонуса, так и для практики клинической и космической медицины.
Цель исследования
- Цель исследования состояла в анализе механизмов формирования тонических моторных реакций.
Задачи исследования
- Исследование механизмов формирования мышечного тонуса как в статике, так и во время движений по показаниям РУ и коленного рефлекса у здоровых людей и больных с нарушениями в центральной нервной системе (ЦНС).
- Исследование стохастической компоненты нервно-мышечных реакций на пассивные перемещения руки.
- Исследование в норме и при моторных дисфункциях стохастической компоненты сухожильных рефлексов с применением различных внешних воздействий, включая прием Ендрассика.
Новизна исследования
- Впервые, с применением новых экспериментальных подходов, выявлен механизм РУ укорочения и показана его роль в формировании и поддержании мышечного тонуса.
- Впервые показана связь РУ со стохастической нейронной активностью -мотонейронов.
- Выявлено сходство свойств РУ с известными нервно-мышечными рефлексами, что ставит механизм этой реакции в ряд спинальных механизмов.
- Показано, что стохастическая активность и РУ в значительной мере ответственны за поддержание тонуса скелетных мышц в статике и во время движений.
- Сконструированы оригинальные устройства для исследования мышечных реакций на пассивные колебательные движения конечности и для исследования коленных рефлексов в изометрическом режиме.
Публикации
По теме диссертации опубликована 41 научная работа, из которых 22 – в журналах, рекомендуемых ВАК РФ. Также по теме диссертации получены авторские свидетельства на 12 изобретений.
Основные положения, выносимые на защиту
- Стохастическая супрасегментарная возбуждающая активность и РУ являются основными механизмами регуляции и поддержания мышечного тонуса как в статике, так и во время движений.
- Стохастическая активность является базовым фактором в механизме РУ.
- Как отсутствие, так и высокая выраженность РУ сопровождают многие формы моторных дисфункций.
Теоретическая и практическая значимость
- В работе представлены экспериментальные подтверждения ведущей роли стохастической активности ЦНС в регуляции мышечного тонуса.
- Показана роль РУ в формировании и поддержании мышечного тонуса как в статике, так и во время движений.
- Выявлено сходство характеристик РУ с характеристиками спинальных рефлексов.
- Показано, что величина спастичности может быть определена количественно по графической зависимости амплитуды коленного рефлекса от энергии удара по сухожилию.
- Исследования, выполненные на клиническом материале, позволили разработать новые технологии как для диагностики, так и для реабилитации неврологических больных.
Структура и объем диссертации
Диссертация состоит из введения, 6 глав собственных исследований, выводов, приложения и списка литературы из 556 источников (156 отечественных и 400 зарубежных). Работа изложена на 207 страницах, иллюстрирована 70 рисунками.
Содержание работы
Методы и материалы исследования
Мышечный тонус определяли, регистрируя механическое сопротивления руки пассивному сгибанию и разгибанию её в локтевом суставе –механомиограму (ММГ) на изменение угла () в локтевом суставе. При проведении исследований испытуемый располагался в кресле с предплечьем одной руки, фиксированном на горизонтальной подвижной плат-форме так, что угол между плечом и предплечьем составлял приблизительно 100°. Руку принудительно сгибали и разгибали в горизонтальной плоскости с амплитудой ±25°. Применение различных скоростей пассивного перемещения руки позволило разделять мышечные реакции на тонические и фазические. Совпадение по фазе ММГ и указывало на то, что пассивная конечность сопротивляется этому перемещению. Величину полного сопротивления (амплитуду между противоположными максимумами ММГ) выражали в единицах момента сил – ньютонометрах (Нм). На разработанные для этих измерений устройства и способы получены 12 авторских свидетельств.
Устройство, компенсирующее на измерительном элементе компоненту, создаваемую моментом инерции подвижной части прибора и перемещаемой конечности, открыло возможности исследования мышечных характеристик в широком диапазоне периодов синусоидальных движений: от 0,5 с до 4 минут (с амплитудой 50о).
При проведении исследовании коленных рефлексов в изометрическом режиме в одномерной системе координат испытуемый располагался в специальном кресле, оборудованном крепежными ремнями, которые обхватывали плечевой пояс и фиксировали испытуемого к креслу на уровне груди, живота и бедер. При необходимости фиксировали также руки и, с помощью специального устройства, голову. Голени испытуемого жестко крепили к двум неподвижным измерительным платформам, исключающим движения в коленных суставах. Измерительные платформы опирались на тензометрические датчики, регистрирующие моменты сил отдельно в каждом коленном суставе. Величину удара по сухожилию определяли по шкале энергий ударов в миллиджоулях (мДж). Электромиограмму (ЭМГ) и ММГ регистрировали с помощью электромиографов фирм “DISA” и "Nicolet". Конструкция устройства обеспечивала воспроизводимость и простоту процедуры измерений.
При исследовании мышечных реакций на однократное линейное пе-ремещение испытуемый располагался в кресле, а его руку закрепляли на платформе, перемещаемой в горизонтальной плоскости, вызывая поворот руки в локтевом суставе на заданный угол и с заданной скоростью.
При исследовании реакций на пассивные колебательные движения руку подвешивали на струнах длиною более 2 м, так что предплечье могло свободно качаться в локтевом суставе в горизонтальной плоскости.
Контингент испытуемых. 402 исследования выполнены у 214 практически здоровых добровольцев. В 9 из них изучали влияние ишемии плеча и предплечья на мышечный тонус верхних конечностей, в 4-х – влияние на мышечный тонус дремотного состояния испытуемых. Ишемию руки создавали при помощи манжетки Рива-Роччи, которую располагали на плече как можно более проксимально. При помощи быстрого сдавливания подключенного к манжетке резинового баллона создавали давление. Окклюзию сосудов осуществляли со скоростью, при которой не имело место нежелательное переполнение системы венозных сосудов. Величину давления в манжетке устанавливали по манометру на уровне, превышающем предварительно измеренное максимальное артериальное давление на 50 - 60%. Исследования мышечных реакций у здоровых испытуемых систематизированы в таблице 1.
Таблица 1.
Виды исследований, выполненных с участием здоровых испытуемых
| № п/п | Способы исследования | Кол-во исследов. |
| 1 | Исследование мышечного тонуса при периоде пассивных перемещений в 14, 6 и 3 с. | 194 70 |
| 2 | Исследование мышечного тонуса при периоде пассив- ных перемещений от 0,5 с до 4 мин. | 105 65 |
| 3 | Исследование коленных рефлексов | 44 40 |
| 4 | Исследования с однократными линейными перемещениями | 43 23 |
| 5 | Исследования маятнико-подобных движений | 16 16 |
| Общее количество исследований и испытуемых | 402 214 |
(В числителе - количество исследований, в знаменателе - количество испытуемых).
В группе здоровых испытуемых исследовали влияния на мышечный тонус облегчающих воздействий - редуцированного приема Ендрассика, звука, света, решения математических задач, эмоциональных нагрузок. Механографические характеристики пассивных регулярных движений руки исследовали в 194 опытах с ограниченным диапазоном скоростей и в 105 - в широком диапазоне скоростей. Результаты этих исследований для правой и левой рук у здоровых людей отличались и часто значительно. Поэтому исследования для правой и левой рук в локтевом суставе при статистической обработке рассматривались как разные случаи. Например, 194 и 105 опытов в таблице 1 реально соответствовали 388 и 210 случаям.
На базе Института нейрохирургии им. акад. Н.Н Бурденко у 392 больных с различными центральными нарушениями выполнено 685 исследований (Таблица 2) и на базе ВНИИ общей и судебной психиатрии им. В.П. Сербского обследовано 30 мужчин в возрасте 20 – 40 лет с реактивными состояниями, сопровождавшимися двигательными расстройствами и изменениями мышечного тонуса вследствие длительно действующего сильного эмоционального стресса.
Таблица 2.
Виды и количество исследований больных людей
| № п/п | Способы исследования | Кол=во исследований | |||||
| Пар. | ДЦП | Моз. | Пер. | Спас. | Реак. | ||
| 1 | Исследование мышечного тонуса при периоде пассивного перемеще- ния 14, 6 и 3 с. | 23 13 | 38 28 | 21 19 | 1 1 | 10 9 | 30 30 |
| 2 | Исследование мышечного тонуса при периоде пассивных перемеще- ний от от 0,5 с до 4 мин. | 136 79 | 223 74 | 20 18 | 101 82 | 46 30 | - |
| 3 | Исследование характеристик коленных рефлексов | 11 7 | 39 19 | 2 2 | - | 10 7 | - |
| 4 | Маятнико-подобное движение | - | - | 4 4 | - | - | - |
| Всего | 170 99 | 300 121 | 47 43 | 102 83 | 66 46 | 30 30 | |
(В числителе показано количество исследований, в знаменателе - количество больных. В таблице применены обозначения: Пар. – больные паркинсонизмом. ДЦП – больные детским церебральным параличом и торсионной дистонией, Моз. –опухоли в задней черепной ямке или в мозжечке. Пер. – нарушениями в периферической системе. Спас. – кривошея или изменение мышечного тонуса по спастическому типу. Реак. – реактивные больные. Общее количество исследований и больных 715/422.)
Большую группу составили больные с экстрапирамидными нарушениями: 170 исследований проведено у 99 больных паркинсонизмом с ригидной, дрожательной, акинетической и смешанными формами заболевания, 300 исследований выполнены у 121 больного детским церебральным параличом (в том числе 17 больных с торсионной дистонией). Эти исследования осуществлялись в ходе выполнения лечебных деструктивных воздействий на различные подкорковые структуры головного мозга, выполнявшихся с целью снижения моторных дисфункций, главным образом, гиперкинезов и спастических компонент мышечного тонуса. 47 исследований были выполнены у 43 больных с верифицированными во время операции поражениями мозжечка.
Методика исследования мышечного тонуса была применена для оценки нарушений в периферической проводниковой системе у больных с травматическим поражением периферических отделов нервной системы на уровне плечевого сплетения и периферических нервных стволов (83 случая), а также при спастической форме нарушения мышечного тонуса сосудистого и опухолевого генеза (46 случаев) и спастической кривошее (12 случаев). Коленный рефлекс исследовали в 35 случаях.
Результаты исследований и их обсуждение
Исследование реакции укорочения
Большинство исследователей согласны с тем, что реакция укорочения (РУ) является одним из важных механизмов поддержания тонического состояния мышц. Однако со времени обнаружения Вестфалем РУ прошло более 100 лет, но достоверного объяснения механизма этой реакции до настоящего времени нет.
На рис.1 представлен результат исследования РУ в голеностопном суставе у больного паркинсонизмом, выполненного в наших экспериментах так же, как это впервые осуществил Вестфаль: угловое пассивное перемещение стопы он осуществлял вручную. Но мы одновременно регистрировали угловое перемещение в голеностопном суставе и электрическую активность сгибателей и разгибателей стопы.

Рис 1. А – пассивное тыльное сгибание стопы, Б – принудительное тыльное разгибание стопы. Обозначения: 1 – ЭМГ m. tibialis ant., 2 - ЭМГ m. gastrocnemius, 3 - угол в голеностопном суставе. Калибровка для каналов 1 и 2 одинакова.
На рис. 1А видно, что пассивное тыльное сгибание стопы сопровождалось выраженной РУ в пассивно укорачиваемой передней большеберцовой мышце. Если после сгибания в голеностопном суставе конечность больного оставляли в покое, стопа сохраняла приданное ей положение многие минуты. При принудительном ступенчатом возвращении через 3 минуты стопы в исходное положение (рис.1Б) активность в растягиваемой передней большеберцовой мышце уменьшалась почти до исходного уровня. При этом каждая последующая ступень растяжения передней большеберцовой мышцы сопровождалась вначале торможением в ней фоновой активности, а затем её восстановлением, но на величину несколько ниже предыдущей.
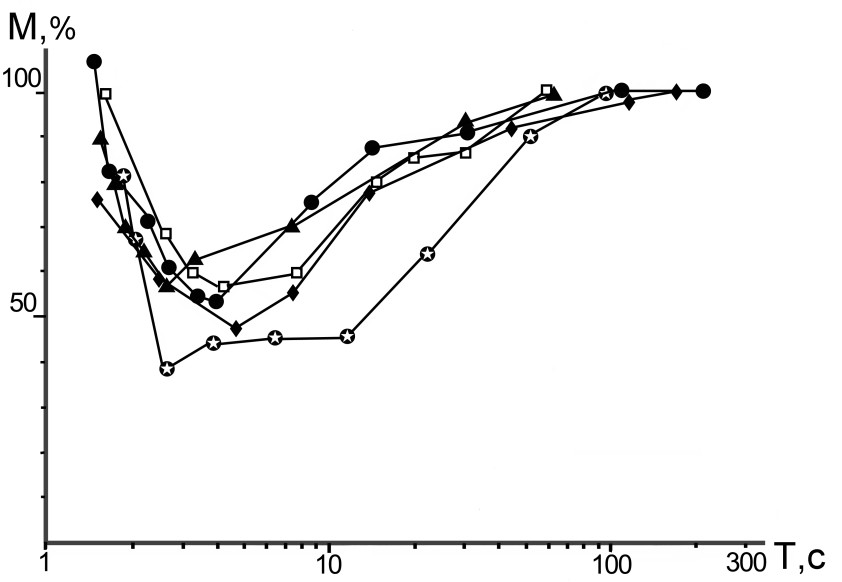
Применение синусоидальных циклических перемещений. На устройстве, позволяющем изменять период синусоидальных перемещений от 0,5 с до 4 мин и амплитудой ±25о, проведены более 100 исследований. На всех полученных графиках в области средних скоростей имело место снижение амплитуд ММГ (рис.2). У здоровых людей в этих экспериментах была обнаружена нелинейная зависимость амплитуд ММГ от периода колебательного цикла: в диапазоне циклических движений от 2 с до 20 с имело место снижение амплитуд ММГ, обусловленное РУ. При дви-жениях с периодом от 100 с и более РУ была невыраженной. При перемещениях с периодом меньшим 2 с в мышцах, напротив, возникала реакция на растяжение и, соответственно, амплитуда ММГ увеличивалась. При ишемии мышц плеча и во время дремотного состояния графики становились параллельными оси абсцисс, а коэффициенты вариации амплитуд ММГ приближались к нулю.
Рис.2. Зависимость амплитуд механо-грамм мышц плеча от величины периода пассивного колебательного перемещения руки у 5 здоровых людей.
По оси абсцисс – период колебательных перемещений в секундах, по оси ординат – амплитуды механограмм в процентах к амплитудам на самой малой скорости пас-сивных движений (при Т 100 с).
У здоровых людей выявлен также широкий разброс форм и величин механического сопротивления пассивным синусоидальным перемещениям конечности. На рис. 3 представлены два крайних типа: А – “пассивный” тип, при котором выраженных реакций на перемещения не обнаруживалось и Б – “активный” тип реакции, при котором ответы в фазу пассивного укорочения мышцы были отчетливо выражены.

Рис.3. Механографическое исследование двух молодых людей 26 лет (А) и 15 лет (Б). Перемещение начиналось с разгибания руки в локтевом суставе. 1 и 2 - ЭМГ сгибателя и разгибателя лок-тевого сустава (50 мкВ для А и 40 мкВ для Б), 3 – угловое движение в локтевом сус-таве, 4 – механическое соп-ротивление пассивной руки перемещению (0,35 Нм для А и 0,65 Нм для Б). В скобках
указана калибровка по раз-
меру "Е".
Все другие типы реакций у здоровых людей располагались между этими формами. Однако даже при пассивном типе реакции коэффициент вариации амплитуд ММГ никогда не был равен нулю. В исследованиях более чем 150 здоровых людей не обнаружено ни одного случая, в котором величины амплитуд ММГ от цикла к циклу были бы постоянны. Исключение составили лишь данные исследований больных с преганг-лионарным отрывом корешков плечевого сплетения от уровня С5 до Th1.
Исследования в изометрии показали, что фоновая активность участвует также и в поддержании мышечного тонуса в статике (рис. 4).

Рис.4. Сопутствующая активность в мышцах плеча у здорового человека, вызванная произвольным прижиманием к большому пальцу указательного паль-ца при двух фиксированных положениях той же руки.
А и Б – регистрации при угловых по-ложениях между плечом и предплечьем 170 и 90 градусов соответственно. 1 и 2 – ЭМГ сгибателя и разгибателя локте-вого сустава соответственно, 3 – произ-вольное усилие в пальцах, сжимающих датчик. Все регистрации ЭМГ выполнены при одном усилении.
В случае, показанном на этом рисунке, исследование осуществляли при двух фиксированных статических угловых положениях между плечом и предплечьем локтевого сустава, равных 170 и 90 градусов. При поддержании изометрического положения мышц плеча в неподвижном состоянии сопутствующая активность оказывалась выше в той мышце, которая была более укорочена. В этом исследовании испытуемый развивал произвольное усилие, прижимая указательный палец этой же руки к большому пальцу. Возникающая сопутствующая активность в двуглавой и трехглавой мышцах плеча оказывалась выше в той мышце, которая была укорочена. Так, когда рука была почти полностью разогнута (рис. 4А), сопутствующая активность возникала в укороченных разгибателях (2-й трек), но не в пассивно растянутых сгибателях (1-й трек). Когда же рука была согнута рис. 4Б) и были пассивно укорочены сгибатели, то именно в них (1-й трек), а не в пассивно растянутых разгибателях (2-й трек) возникала сопутствующая активность. В этом исследовании не было движения. Поэтому облегчение мотонейронов, иннервирующих укороченные мышцы, не может быть следствием афферентного влияния веретенных рецепторов.

Исследования с однократными пассивными перемещениями. В примере рис.5 РУ у здорового человека возникла при пассивном сгибании руки со скоростью 14о/с на угол 15о из углового положения между плечом и предплечьем 100о.
Рис.5. А – испытуемый находится в пассивном состоянии, Б – испытуемый развивает максимальное усилие сжимания динамометра кистью противоположной руки.
1 и 2 – ЭМГ сгибателя и разгибателя локтевого сустава соответственно; 3 – угол в локтевом суставе. Калибровка ЭМГ одинакова для обеих регистраций.
Перемещение вызывало в укорачиваемой двуглавой мышце плеча (1-й трек) РУ, вероятно, вследствие имеющей место подпороговой фоновой активности в -мотонейронах. Из регистрации А в 1-м треке видно, что РУ представлена несколькими двигательными единицами (ДЕ). Когда испытуемый развивал одновременно максимальное усилие сгибателями кисти другой руки, мощность РУ, за счет возникшей сопутствующей активности, возрастала, проявляясь увеличением количества участвующих ДЕ и частоты их импульсации (рис. 5Б), как если бы сопутствующая активность суммировалась с фоновой активностью, увеличивая мощность РУ, или увеличивалась сама фоновая активность. После прекращения движения активность в укороченной мышце некоторое время сохранялась, постепенно уменьшаясь. Причиной этого последействия могло быть следовое возбуждение тонической активности, генерируемой малыми тоническими нейронами и/или реологическими свойствами мышечной ткани.
Если руку перемещали при исходно максимально укороченной мышце, латентное время РУ могло быть достаточно коротким. На рис.6 показан пример РУ в мышцах плеча (1-й трек), возникшей во время пассивного перемещения руки в локтевом суставе со скоростью 250о/с из углового положения между плечом и предплечьем 60о в положение 50о. Для данного испытуемого эти угловые положения локтевого сустава были анатомически максимально возможными. До начала перемещения в двуглавой мышце уже имелась небольшая фоновая активность, возможно, вследствие имевшего место укорочения. Латентное время РУ в этом случае составило около 20 мс. Латентное время одновременно возникшего рефлекса на растяжение в трехглавой мышце плеча было несколько большим. Поскольку угол в локтевом суставе регистрировали устройством из тонкой пружины с наклеенными на нее тензодатчиками и браншами для крепления к руке, в пружине после резкой остановки сгибательного движения возникали небольшие механические автоколебания.

Рис. 6. Реакции в двуглавой и трехгла-вой мышцах плеча на принудительное сги-бание руки в локтевом суставе упругими тя-гами со скоростью 250о/с.
1 - ЭМГ двуглавой мышцы плеча, 2 - ЭМГ трехглавой мышцы плеча, 3 - сгибание руки в локтевом суставе на анатомически допус-тимый угол в 10о. Регистрации ЭМГ осу-ществлены при одинаковом усилении.
В пределах, обычно применяемых диапазонов перемещения конеч-ности, латентное время РУ в наших исследованиях составляло несколько более 40 мс, что совпадало с данными литературы (Foix, Thvnard, 1923). Исследователи полагали, что такое время является результатом реализации РУ через длинные рефлекторные пути. Однако увеличенное латентное время можно объяснить и влиянием реологии мышц. В максимально укороченной позиции этот механический эффект нивелируется рефлекторно возникающим в мышце напряжением, создающим условия для более быстрой трансляции механического воздействия на сухожильные рецепторы. Латентное время РУ при таких условиях оказывается сопоставимым с временем реакций, опосредуемых другими проприорецепторами.
Произвольная активация тестируемой мышцы. Результаты выполненных исследований показали, что выраженность РУ зависит от величины облегчающих влияний. При этом произвольное усилие в мышцах кисти контралатеральной руки оказывало меньший эффект, нежели усилие в мышцах расположенных сравнительно близко. Чем большим было развиваемое испытуемым усилие в кисти, тем большей была и РУ. Максимально возможное облегчение наблюдалось при произвольной активации самой тестируемой мышцы.
В исследовании, данные которого представлены на рис.7, испытуемый исходно удерживал умеренное напряжение в сгибателях и разгибателях плеча. На этом фоне электромеханическое устройство однократно сгибало руку в локтевом суставе по синусоидальному закону (чтобы исключить влияние ускорения) из углового положения между плечом и предплечьем в 100о в положение 75о со средней скоростью 60о/с. В растягиваемой трехглавой мышце (2-й трек) возникало торможение произвольной активности, продолжавшееся еще некоторое время после прекращения растяжения. В одновременно укорачиваемой двуглавой мышце (1-й трек) активность возрастала до 1 мВ.

Рис. 7. Реакции в мышцах плеча у здорового человека на принудительное сгибание руки в локтевом суставе электрическим двигателем во время произвольного напряжения тех же мышц. В скобках указана калибровка по размеру "Е". 1 - ЭМГ двуглавой мышцы плеча (0,3 мВ), 2 – ЭМГ трехглавой мышцы плеча (0,2мВ), 3 – сгибание руки.
Исследование суммации возбуждения при повторяющихся вызовах реакции укорочения. Во время повторяющихся экскурсий за счет активности на мотонейронах, опосредующих РУ, от цикла к циклу накапливается следовая деполяризация, относящаяся к механизму РУ. Схематически этот процесс показан на рис. 8.

Рис.8. Моменты возникнове-ния реакции укорочения в дву-главой мышце плеча от цикла к циклу во время пассивных си-нусоидальных перемещений при трех указанных над графи-ками периодах. По оси орди-нат – угол между плечом и предплечьем (70° соответст-вует положению, когда двугла-вая мышца укорочена). Стрел-ки слева указывают на после-довательность фаз растяже-ния и укорочения двуглавой мышцы плеча.
График свернут в трубку, так что верхняя и нижняя линии “70о ” являются одной и той же линией. Синусоидальное перемещение начиналось с разгибания руки (растяжения двуглавой мышцы) из положения между плечом и предплечьем 70о (нижняя линия). Растяжение заканчивалось при положении локтевого сустава 120о, затем следовало сгибание руки, сопровождающееся пассивным укорочением двуглавой мышцы. В первом цикле РУ по ЭМГ возникла в положении 87о – кружок 1. После достижения 70о пассивное укорочение мышцы завершилось и сменилось фазой растяжения. Во втором цикле РУ возникла уже при 97о (кружок 2). Последовательность моментов возникновения реакций показана линиями, соединяющими кружки. На графике видно, как от цикла к циклу на одной и той же частоте момент возникновения РУ сдвигался к началу фазы укорочения. Сначала он приближался к началу фазы укорочения двуглавой мышцы (укорочение начиналось на линии 120о) и иногда возникал уже в конце предшествующей фазы перемещения, когда мышца еще растягивалась. Возникшая в этой точке “активность на растяжение” в конце периода растяжения сохранялась на всём протяжении в последующей фазе укорочения, исчезая с началом очередной фазы растяжения. И если при периоде колебательного движения в 7 с смещения моментов возникновения реакции происходили сравнительно медленно, то при 0,9 с уже в третьем цикле очередная реакции возникала в конце фазы растяжения мышцы, хотя в следующем 4-м цикле момент начала возбуждения вновь возвращался в фазу укорочения. При этом времена появления активности становились нестабильными, перемежаясь - в конце фазы растяжения мышцы (точками 3, 5, 7, 8) и в начале фазы укорочения (точки 2, 4, 6). Можно предположить, что следовое возбуждение, возникающее в фазу укорочения мышцы и сохраняясь на мотонейроне в слеующую фазу растяжения, облегчало реакцию на растяжение.
Реакция укорочения и двигательные единицы (ДЕ). При выполнении циклических перемещений, начиная с некоторой очередной фазы, последующие реакции укорочения, возникающие с уменьшенной латентностью, начинались с двойных разрядов ДЕ, так называемых “дуплетов” (3-й и 4-й циклы на рис.9). Согласно данным Л.П. Кудиной и Р.Э. Андреевой (2004), дуплеты являются признаком накопления на -мотонейронах следового возбуждения. В нашем случае возникновение дуплетов, совпадавшее со снижением порогов мотонейронов, возникавшим в предыдущую фазу укорочения, также указывало на то, что на -мотонейронах

в фазу укорочения “их мышцы” имело место накопление возбуждения.
Рис.9. Генерация реакции укорочения от цикла к циклу в двуглавой мышце плеча во время пассивных синусоидальных движений с периодом около 7 с и амплитудой 500. 1 – ЭМГ сгибателя локтевого сустава; 2 – угол в локтевом суставе. Движение начиналось с разгибания руки в локтевом суставе. Стрелки указывают дуплеты.
Эффекты эмоциональной напряженности. Эмоциональное состояние человека влияло на выраженность РУ. Если испытуемый перед исследованием был чем-либо взволнован, выраженность РУ была больше, чем в те дни, когда он был спокоен. Если испытуемый, напротив, был вял и сонлив, выраженность РУ значительно снижалась, почти полностью исчезая во время сна. Внешние воздействия в наших исследованиях приводили к увеличению этой реакции (И.Д. Гедельман и др.,1966; В.А. Сафронов, 1970; 1984), что совпадает с данными и других исследователей (Soeken, 1936; Andrews et al., 1973). Особенно высокой была РУ у людей при сильном и длительно действующем эмоциональном стрессе (В.А. Сафронов, Т.Г. Морозова, 1988).
Влияние позной активности на выраженность реакции укорочения. Поддержание прямой позы увеличивает сопутствующий нисходящий возбуждающий фон. РУ увеличивалось при поддержании испытуемым позы на жестком стуле в сравнении с мягким креслом (рис.10). При этом РУ у всех испытуемых возрастала, возможно, за счет сопутствующей активности, вызванной одновременно с познотонической активностью, направленной на поддержание позы в условиях неудобного сидения.

Рис. 10. Исследование реакции мышц на синусоидальные колебательные перемещения руки с периодом 8 с в локтевом суставе здорового испытуемого. А - испытуемый располагается в мягком удобном кресле и Б - на жестком канцелярском стуле. 1 и 2 - ЭМГ сгибателя и разгибателя локтевого сустава (58 мкВ), 3 – механограмма (0,5 Нм), 4 – фазовая кривая (разгибанию соответствует движение кривой вверх). В скобках указана калибровка по размеру “Е”.
Нестабильность реакций у 8 здоровых испытуемых на качества сидений была оценена по коэффициенту вариации амплитуд ММГ и показана в таблице 3.
Таблица 3.
Влияние комфортабельности сидения на величину коэффициента вариации (С)
| Коэфф. вариации ± станд. ошибка | Разница: с1 – с2 | Достовер-ность: Р | |
| Канцел. стул (С1) | Мягкое кресло (С2) | ||
| 14,22 ± 1,87 | 5,14 ± 0,76 | 9,08 ± 2,02 | 0,9999 |
| 8,33 ± 0,72 | 2,57 ± 0,17 | 5,76 ± 0,74 | 0,9999 |
| 3, 04 ± 0,27 | 2,20 ± 0,14 | 0,84 ± 0,31 | 0,9934 |
| 4, 89 ± 0,25 | 2,07 ± 0,13 | 2,82 ± 0,28 | 0,9999 |
| 3, 79 ± 0,24 | 2,71 ± 0,17 | 1,08 ± 0,30 | 0,9999 |
| 20,50 ± 1, 39 | 12,68 ± 0,82 | 7,82 ± 1,62 | 0,9999 |
| 12,46 ± 0,83 | 3,02 ± 0,20 | 9,44 ± 0,86 | 0,9999 |
| 9,90 ± 0,62 | 3,38 ± 0,23 | 6,52 ± 067 | 0,9999 |
Обозначения в таблице: (С1 – С2) – разница между коэффициентами вариации ММГ при сидении испытуемого на канцелярском стуле (С1) и на мягком кресле (С2); Р – достоверность разницы между коэффициентами вариации.
Модель механизма реакции укорочения. Результаты проведенных исследований позволили предположить, что базовым механизмом РУ является фоновая тоническая активность (надпороговая или подпороговая) -мотонейронов, обусловливаемая как афферентными входами, так и супрасегментарными влияниями. Модель механизма РУ представлена на Рис.11.

Рис.11. Обозначения: F – фоновые влияния, поступающие на пул -мотонейро-нов; -МН – пул -мотонейронов; R – клетка Реншоу; G – сухожильный рецептор Гольджи; pp – рецептор длины; В – компонент мышечной вязкости; K1 и K2 – упругие элементы мышцы, последовательные с рецептором Гольджи; K3 – упругий элемент мышцы, соединенный параллельно рецептору Гольджи; S – проксимальное и дистальное сухожилия мышцы; P – силы растяжения, приложенные к мышце. Черные и белые кружки – соответственно тормозные и возбуждающие нейроны и синапсы.
Базовым фактором в этой модели является фоновая активность (F) на входе -мотонейронного пула (В.А. Сафронов,1984). Процесс генерации РУ в модели происходит в следующем порядке: при пассивном сближении сухожилий мышцы (S) освобождаются упругие элементы (k). В результате снижается интенсивность воздействия элемента k1 (и k2) на сухожильный рецептор Гольджи (G) и, тем самым, уменьшается его тормозное влияние на -мотонейрон. Возникает своеобразное “открытие ворот”, и существующая на входе мотонейрона фоновая активность (F) поступает к мышце. В таком “пассивном” состоянии мышца может оставаться довольно продолжительное время.
Если применить к мышце растяжение (Р), то вначале, из-за того, что вязкая компонента B препятствует деформации компоненты k2, произойдет растяжение только упругого элемента k1 (и k3), которое создаст силовое воздействие на рецептор Гольджи и, следовательно, вызовет торможение -мотонейрона. Затем вязкость (B) постепенно отпускает упругий элемент k2, в результате чего деформация перераспределяется между упругими элементами k2 и k1, за счет чего силовое воздействие на рецептор Гольджи снизится, уменьшая интенсивность тормозных влияний на -мотонейрон. Этим событиям соответствует изменение во времени активности мышц на рис.1Б. Согласно взаимодействию между элементами модели, РУ является результатом модуляции фоновой активности мотонейронов тормозной афферентацией от рецепторов Гольджи. Таким образом, РУ обеспечивает приспособление длины мышцы к более укороченной позиции. Заметим, что все характеристики РУ объясняются предложенным механизмом и, следовательно, можно полагать, что механизм этой реакции один и тот же во всех случаях его проявления. Из результатов исследований, аналогичных представленным на рис. 4, следует, что и в условиях изометрии пороги -мотонейронов ниже, когда мышца укорочена по сравнению с тем, когда она растянута, что объясняется снижением тормозных влияний от рецепторов Гольджи укороченной мышцы.
Резюме. При пассивном укорочении мышцы уменьшается гомонимное торможение мотонейронов рецепторами Гольджи, что приводит к открытию “ворот” и трансляции фоновой активности к мышцам. Модель объясняет все свойства РУ, выявленные в наших исследованиях в норме и при патологии, а также в исследованиях, описанных ранее в литературе. Полученные нами результаты подтверждают представления других исследователей, согласно которым РУ является аутогенной реакцией, создающей напряжение в мышцах при их пассивном укорочении. Она в равной степени проявляется в сгибателях и в разгибателях. Она обнаруживает свойство суммации возбуждения, типичное для других спинальных рефлексов, и не является результатом реципрокных отношений между укорачиваемой мышцей и ее антагонистами, как не является и произвольной. Модель, построенная на основании данных исследований, предполагает, что РУ возникает в результате модуляции фоновой тонической активности -мотонейронов афферентными влияниями от сухожильных рецепторов.
Исследования коленных рефлексов
Erb (1875) и Westfal (1875) в одновременно опубликованных ими двух работах положили начало активным исследованиям механизма коленного рефлекса и разработке методов и устройств для его количественного измерения (И.И. Русецкий, 1935; Е.С. Арутюнов, 1953; З.Х. Абазова и др, 2006; Lombard, 1889; Zhang et al, 1999; Mamizuka et al, 2007).
Наиболее распространенным методом в исследовании рефлекторных ответов явилась электромиография. Но электромиографическая регистрация поверхностными биполярными электродами дает сведения только об активности мышечных волокон, расположенных вблизи этих электродов, но не об активности всех мышц данной группы. Поэтому нам представлялось более адекватным оценивать рефлекторные реакции по механическому напряжению мышц.
Величину коленного рефлекса часто определяют по амплитуде рефлекторного движения свободного сегмента конечности (Lombard, 1889; Sommer, 1899; В.М. Бехтерев, 1915; А.Г. Чеченин и др. 2000; Mra-chacz-Kersting, Sinkjaer, 2003; Абазова и др., 2006; Mamizuka et al, 2007). Однако на результаты этих измерений большое влияние оказывают механические помехи: после каждого рефлекторного движения изменяется исходное положение исследуемого звена, перемещается место нанесения удара перкуторным молоточком, а, следовательно, изменяются начальные условия и эффективность удара. На рефлекторный ответ оказывает влияние и реакция одновременно растягиваемого антагониста. Эти недостатки отсутствуют в случае измерения момента сил в изометрических условиях (Clarke, 1965; Davis, Beale, 1970; Dietrichson, Srbye, 1971; Simons, Lamonte, 1971; Simons, Dimitrijevic, 1972; Коц Я.М., Болховских Р.Н. 1975; Sinkjaer et al., 1988; Zhang et al., 2000; Nozaki et al., 2005).
Для измерения рефлекторно возникающего момента сил в изометрическом режиме было сконструировано устройство, защищенное авторским свидетельством (В.А. Сафронов, 1998). Рефлекторный ответ регистрировали одновременно по развиваемому моменту сил в коленном суставе и по ЭМГ. Степень воздействия молоточка на сухожилие калибровали по энергии удара. Стохастические супрасегментарные влияния на рефлекторные ответы оценивали по коэффициенту вариации их вели-чин. Также исследовали влияния внешних воздействий таких, как свет, звук, прием Ендрассика.
Об облегчении сухожильных рефлексов при произвольном напряжении мышц спины и рук впервые сообщил венгерский врач Ендрассик (Jendrassik, 1883). Согласно его данным, в условиях, когда испытуемый произвольно создает усилие растяжения в сцепленных перед грудью руках, имеет место облегчение коленного рефлекса. Мы в своих экспериментах применяли модифицированный прием Ендрассика: для облегчения моторных реакций испытуемые только кистью сжимали динамометр.
Зависимость величины рефлекса от энергии удара. У большинства здоровых людей была выявлена выраженная зависимость амплитуды коленного рефлекса от энергии удара по сухожилию (рис.12). Как следует из рисунка, эти графики, как правило, имели максимумы при энергиях удара от 60 до 90 мДж. При бльших энергиях удара амплитуда ответов снижалась. Подобная зависимость коленного рефлекса от энергии удара сохранялась и при выполнении приема Ендрассика, хотя амплитуда ответов в этих случаях могла возрастать в 10 раз. Снижение амплитуды рефлекса при бльших энергиях ударов, возможно, обусловлено торможением -мотонейронов афферентными влияниями рецепторов Гольджи, так как вследствие механического шунтирования этих рецепторов параллельными экстрафузальными мышечными волокнами тормозной эффект от них, согласно модели (рис. 11), может проявляться лишь при больших энергиях удара по сухожилию. У испытуемых с большой мышеч-ной массой применяемых энергий ударов оказалось недостаточным, что-бы обнаружить подобную нелинейную зависимость – 2 нижних графика.

Рис.12. Зависимость амплитуды коленного рефлекса от энергии удара молоточком по сухо-жилию в группе здоровых испытуемых.
По оси абсцисс – энергия удара в мДж, по оси ординат – амплитуда коленного рефлекса в Нм.

Во время приема Ендрассика, по сравнению со спокойным состоянием человека, коэффициенты вариации оказывались более выраженными и коррелировали с величиной коленного рефлекса (Рис.13).
Рис.13. Зависимость рефлекторного ответа (1 и 2) и коэффициента вариации (кружки) от энергии удара в миллиджоулях (мДж) в спокойном состоянии испы-туемого (1 и светлые кружки) и при выполнении прие-ма Ендрассика (2 и темные кружки). Каждая точка получена усреднением данных не менее 10 проб. По левой оси ординат - амплитуда коленных рефлексов, по правой оси ординат – коэффициенты вариации. По оси абсцисс – энергия удара молоточком.
В этом исследовании при спокойном состоянии человека коэффициент корреляции равнялся 0,1444, что соответствовало по шкале Чеддока слабой тесноте связи, а на фоне приема Ендрассика этот коэффициент увеличивался до -0,8763, соответствуя по этой же шкале высокой, но обратной по знаку тесноте связи: с повышением амплитуды рефлекса коэффициент вариации уменьшался. Вероятно, это связано с тем, что чем выше амплитуда коленного рефлекса, тем больше активируется -мотонейронов, опосредующих этот рефлекс, а значит меньше остается “свободных” от участия в рефлексе -мотонейронов, которые могли бы реализовать разброс рефлекторных ответов.
Влияния на коленные рефлексы звука. Амплитуда коленного рефлекса повышается при предъявлении звука. Так, у испытуемого (28 лет) в спокойном состоянии средняя амплитуда рефлекса на удар молоточка с энергией 76,06 мДж была 4,50 Нм. При подаче звука порядка 100 дб средняя амплитуда рефлекса увеличилась до 6,24 Нм (Рис.14).

Рис.14. 1 – усредненная величина коленного рефлекса при спокойном расслабленном состоянии испытуемого и 2 – при подаче в наушники сильного звука. Достоверность разности средних значений, оцененная по двухвыборочному t-тесту, достаточна: t = 4,34 (при критическом значении t = 2,14). Рас-хождение средних величин значимо с надежностью более 0,95.
У животных неспецифический звуковой раздражитель также повышает возбудимость -мотонейронов по отношению к фону на 180 – 200% (М.Е. Иоффе, 1975). Таким образом, звуки повышают фоновую тоническую активность мотонейронов, иннервирующих скелетные мышцы. Противоположный эффект полного отсутствия звуков описал И.П. Павлов в экспериментах с собаками в “Башне молчания”. В “Башне” собаки засыпали. Анализируя эти данные, И.П. Павлов заключил, что для нормальной работы мозга необходим постоянный приток внешних раздражителей, однообразие же впечатлений снижает тонус коры головного мозга.
Влияния на коленные рефлексы света и темноты. Для исследования влияния освещенности на коленные рефлексы глаза освещали диффузным белым светом различной интенсивности от матовых электрических лампочек. Две лампочки размещали отдельно перед каждым глазом испытуемого в устройстве, которое не пропускало иного света. При выключенном освещении испытуемые не обнаруживали извне ни малейшего проблеска света. Интенсивность освещения была прокалибрована в люксах. Во время исследований испытуемые держали глаза открытыми. Энергию удара выбирали, ориентируясь на наиболее устойчивые рефлекторные ответы. Все исследования выполнены в первую половину дня: от 10 до 14 часов.
Интенсивность освещения сетчатки глаз белым светом, вероятно, активирует супрасегментарные фоновые тонические влияния. С увеличением освещения нелинейно увеличивается амплитуда рефлекса и одновременно снижается коэффициент вариации (рис.15).

Рис.15. Зависимость амплитуды коленного рефлекса (треугольники) и коэффициента вариации (кружки) от величины освещения открытых глаз. Каждая точка получена ус-реднением не менее 10 проб. По левой оси ординат – амплитуда рефлексов, по правой оси ординат – коэффициенты вариации. По оси абсцисс с логарифмической шкалой – величина освещения глаз в люксах [лк]. Энергия удара молоточка равнялась 8,62 мДж.
При возрастании интенсивности света от 85 лк и до максимальной, амплитуда коленного рефлекса увеличилась почти в 10 раз. При этом коэффициент вариации снизился в 5 раз, вероятно вследствие того, что при большой интенсивности света в данном пуле оставалось мало “свободных” -мотонейронов для реализации разброса амплитуд рефлекса.
У здоровых испытуемых в начальном диапазоне от 0 лк (полная темнота) до 85 лк обнаруживалась зависимость, противоположная влиянию больших освещений (рис.16). По мере увеличения освещения от полной темноты до 85 лк амплитуда коленного рефлекса снижалась, и только при увеличении освещения более 85 лк она начинала возрастать как на рис. 15. Статистическая оценка разницы средних значений коленного рефлекса показала, что его величина в точке 3 графика на рис.16 достоверно отличается от значения в точке 1 (Р = 0,991), в точке 4 отличие было еще более достоверным (Р = 0,999) и только для точки 2 эта разница не была достоверной. Заметим, что такая монотонная зависимость обнаруживалась не всегда, так как на регистрируемый процесс обычно накладывается значительная вариабельность, характерная для низкого уровня рефлекторных процессов. Для некоторых испытуемых подъем графика рефлекторных ответов начинался после точки 3, поэтому для оценки влияния темноты на коленные рефлексы сравнение

проводили со значением коленного рефлекса в точке 3 (37 лк).
Рис.16. Зависимость усредненных зна-чений коленного рефлекса от величины слабого освещения открытых глаз.
По оси ординат – средняя амплитуда коленного рефлекса в Нм. По оси абсцисс – величина освещения глаз в люк-сах. Значения освещения даны по ли-нейной шкале. Каждая точка получена усреднением не менее 10 проб. Точка 1 на графике соответствует полной темно-те, точка 2 – освещению 11 лк, точка 3 – 37 лк, точка 4 – 85 лк.
Суммарные результаты всех исследований сведены в таблицу 4. Сравнивание значений коленного рефлекса в точке 2 на рис.16 со зна-чением в темноте не проводили, так как в этой точке величина разности рефлексов была слабо достоверной (1-е исследование в таблице 4).
Таблица 4
Реакции коленного рефлекса здоровых испытуемых на полную темноту
| № | Пол | Е | А% | P(А) | n | № | Пол | Е | А% | P(А) | n | |
| 1 | 11 | +12,32 | 0,724 | 18 | 13 | 37 | +24,5 | 0,997 | 79 | |||
| 2 | 37 | +41,78 | 0,997 | 20 | 14 | 37 | +13,6 | 0,999 | 79 | |||
| 3 | 37 | +16,38 | 0,941 | 30 | 15 | 37 | +44,0 | 0,999 | 79 | |||
| 4 | 37 | +35,20 | 0,992 | 40 | 16 | 37 | +25,0 | 0,995 | 80 | |||
| 5 | 37 | +28,44 | 0,992 | 40 | 17 | 37 | +5,23 | 0,898 | 80 | |||
| 6 | 37 | +4,538 | 0,724 | 84 | 18 | 37 | +8,85 | 0,769 | 81 | |||
| 7 | 37 | +40,84 | 0,995 | 84 | 19 | 37 | +6,49 | 0,989 | 80 | |||
| 8 | 37 | +30,84 | 0,960 | 78 | 20 | 37 | +11,0 | 0,966 | 98 | |||
| 9 | 37 | +5,599 | 0,771 | 80 | 21 | 37 | +27,0 | 0,999 | 80 | |||
| 10 | 37 | +29,90 | 0,995 | 80 | 22 | 37 | +8,69 | 0,905 | 80 | |||
| 11 | 37 | +12,18 | 0,898 | 80 | 23 | 37 | +3,34 | 0,693 | 100 | |||
| 12 | 37 | +12,37 | 0,977 | 80 | 24 | 37 | +2,89 | 0,655 | 80 |
В таблице указаны порядковые номера, пол испытуемого, величина освещения Е в люксах, отношение разности величин коленного рефлекса в темноте и при освещении Е к рефлексу в темноте в процентах (А%), доверительная вероятность P(А) и количество проб (n), использованных в статистической обработке. Разницу А% считали достоверной, если соответствующая величина доверительной вероятности была равна или больше 0,95. Из-за большой вариабельности рефлекторных ответов выполняли по 100 и более проб. Результаты первых 20 проб не учитывали, так как у некоторых испытуемых в начальный период исследования имела место адаптация к условиям эксперимента. В статистическую обработку включали только результаты 80 проб, выполненных попеременно по 20 проб в темноте и с освещением. Из таблицы видно, что у 14 испытуемых амплитуда коленного рефлекса в темноте была достоверно выше амплитуды при освещении. И хотя у 10 испытуемых эта разница была не столь достоверна, она всегда имела положительный знак: не было ни одного случая, в котором величина рефлекса в темноте была бы ниже, чем при освещении в 37 лк. Широкий разброс значений А% обусловлен различием в личностной реакции отдельных испытуемых на темноту, а также, возможно, и специфическим отношением к тому, какая интенсивность освещения для каждого из них являлась наиболее комфортной. Повторные исследования, выполненные у одного и того же человека в отдельные дни, показали, что разность величин коленного рефлекса в темноте и при освещении сохранялась, хотя и была измененной. Устно испытуемые отмечали, что освещение в 37 лкс по отношению к темноте создавало ощущение спокойствия. Темнота же вызывала некоторое состояние тревожности. И хотя не все испытуемые эту разницу ощущали, она, по-видимому, всегда имела место. Этот результат связан, вероятно, с рудиментарной тревожностью на темноту, сохранившейся у человека с первобытных времен.
Влияние позной активности на величину рефлекторных ответов исследовали в экспериментах с разгрузкой шейных мышц. Для этого при-меняли опору для головы, устраняющую необходимость удерживать её произвольно. В этих условиях средняя амплитуда коленного рефлекса значительно снижалась (рис. 17).
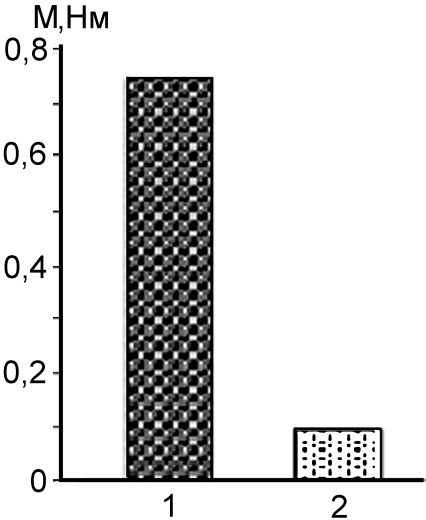
Рис.17. 1 – усредненная величина коленного рефлекса у испытуемого, сидящего в кресле в спокойном расслаблен-ном состоянии, когда его голова находится в привычной вер-тикальной позе; 2 – голова поддерживается опорой так, что для человека исключается необходимость её удерживать. Одновременно наступало расслабление шейных мышц.
Как показывает рисунок, при энергии удара в 49,5 мДж достоверность разности средних величин рефлекса в двух пробах, в одной из которых голова поддерживалась опорой, оцененная по двухвыборочному t-тесту, была высокой: t = 11,83, при критическом значении t = 2,10. Отличие средних значений также было значимо с надежностью более 0,999. Результаты исследований дают основание полагать, что имеющаяся в естественных условиях позная активность сопровождается повышенной сопутствующей тонической активностью многих скелетных мышц, в том числе и мышц, не участвующих в поддержании позы.
Влияние приема Ендрассика на коленный рефлекс. Результаты исследований показали, что степень облегчения коленного рефлекса, обусловливаемого произвольным сжатием динамометра кистью, коррелирует не с активностью произвольно напрягаемых сгибателей кисти, а с величиной суммарной сопутствующей активности других мышц тела.
На рис.18 представлены графики зависимости амплитуды коленного рефлекса: 1 - от суммарной активности сгибателей пальцев и разгибателей кисти, 2 – от усредненной активности мышц тела (прямой мышцы бедра, трапециевидных мышц справа и слева, широчайшей мышцы спи-ны, большой грудной мышцы, дельтовидной мышцы и двуглавой мышцы плеча) и 3 – рефлекса от величины силы сжатия динамометра кистью.
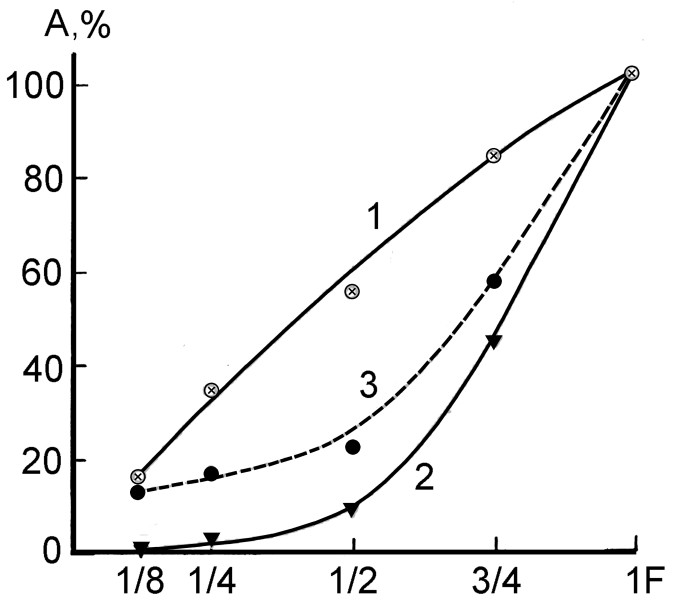
Рис.18. Зависимость амплитуды коленного реф-лекса: 1 – от усредненной амплитуды суммарной активности сгибателей пальцев и разгибателей кисти, 2 – от сумммарной активности других мышц тела, 3 – амплитуды рефлекса от величины силы, развиваемой кистью руки. По оси абсцисс – сила относительно максимальной силы, развиваемой кистью, по оси ординат – величина электрической активности коленного рефлекса в процентах к его величине при максимальной силе развиваемой кистью.
Из сравнения представленных графиков видно, что суммарная активность сгибателей пальцев и разгибателей кисти является приблизительно линейной функцией от силы, развиваемой кистью (график 1). Графики же величины коленного рефлекса (график 3) и суммарной активности других мышц тела (график 2) выражают подобные друг другу приблизительно квадратичные зависимости от силы, развиваемой кистью руки.
Также и у больных с парезами мышц руки вследствие повреждения периферических нервов или после хирургической операции на головном мозге на фоне безуспешной попытки сжимать кисть паретичной руки в кулак в вызванную активность вовлекаются другие мышцы, с сохранной иннервацией. Все эти факты исключают предположение, что коленный рефлекс облегчается афферентацией от проприорецепторов кондиционирующей мышцы и подтверждают, что усиление рефлекторной активности при приеме Ендрассика обусловливается сопутствующей активацией. Это явление похоже на известные в неврологии у больных с центральным парезом распространенные синкинезии, возникающие при попытках движения паретичной конечностью. Однако описываемый нами механизм активации только внешне подобен явлению синкинезии. Сопутствующая активность в данном случае родственна тонической, а не двигательной. Поэтому, по аналогии со словом “синкинезия“, такую активность следует именовать “синтонией” или “синтонической активностью”.
Динамика рефлекторных ответов при приеме Ендрассика. В этом разделе повторены исследования Delwaide и Toulouse (1977; 1980; 1981) временной зависимости облегчения сухожильных рефлексов от начала развития усилия. Однако, в отличие от проведенных этими авторами исследований, временной диапазон процесса облегчения нами был расширен до 25 с (рис.19).
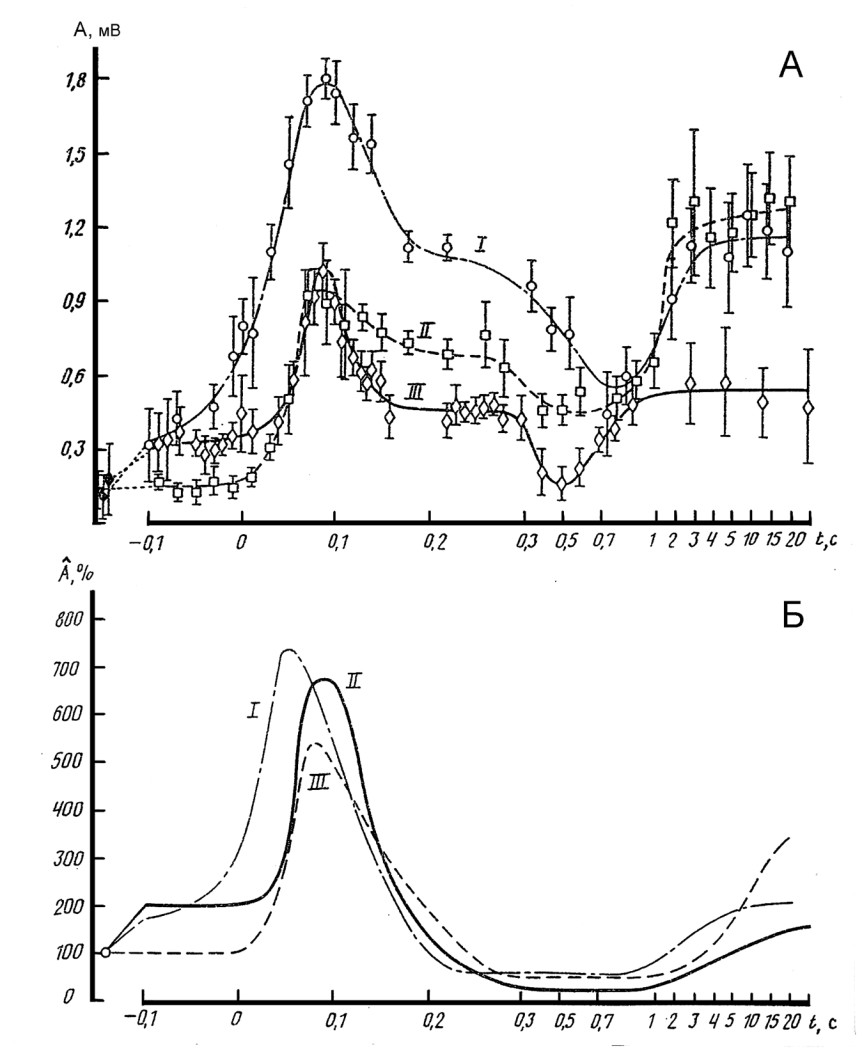
Рис.19. Динамика изменения величины коленных рефлек-сов у трех здоровых испытуемых в процессе развития и поддержания ими произволь-ного усилия мышцами руки.
А - Усредненные амплитуды коленного рефлекса по всем пробам. Б – Нормиро-ванные по произвольному усилию усредненные ампли-туды коленного рефлекса из графики А. Точка “0” соот-ветствует началу развития произвольного усилия.
Эта многофазность описывается аналитическим выражением:
![]()
Согласно этому выражению, амплитуда коленного рефлекса зависит от настроечных влияний S(t) до начала развития произвольного усилия. Затем следует динамическое слагаемое, связанное со скоростью из-менения кондиционирующего усилия ![]() , которое через 100 мс после начала произвольного усилия достигает максимума, а через 300 мс становится пренебрежимо малой величиной. В третью фазу наступает умерен-ное облегчение, генерируемое супрасегментарными влияниями, связанными с поддержанием произвольного усилия, преводящее,вероятно, к утомлению, определяемому слагаемым V(t), при временах
, которое через 100 мс после начала произвольного усилия достигает максимума, а через 300 мс становится пренебрежимо малой величиной. В третью фазу наступает умерен-ное облегчение, генерируемое супрасегментарными влияниями, связанными с поддержанием произвольного усилия, преводящее,вероятно, к утомлению, определяемому слагаемым V(t), при временах ![]() . Помимо этого, облегчение зависит также от квадрата величин произвольного усилия F2 и от некоторой исходной фоновой компоненты B.
. Помимо этого, облегчение зависит также от квадрата величин произвольного усилия F2 и от некоторой исходной фоновой компоненты B.
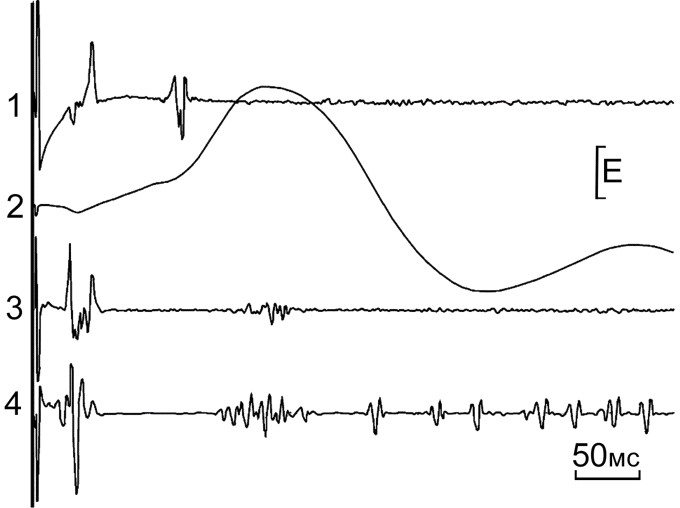
Коленные рефлексы у неврологических больных. Сравнение раз-вернутых во времени рефлекторных ответов здоровых людей при выполнении ими приема Ендрассика (рис. 20) с рефлекторными ответами больных ДЦП (рис.21) не выявило выраженных отличий в ответах здорового и больного человека.
Рис.20. Коленный рефлекс у здорового испытуемого 23 лет на удар молоточком с энергией 49,5 мДж по сухожилию левой ноги при выполнении приема Ендрассика.
1 – поверхностное отведение с m. biceps femoris (50мкВ), 2 – ММГ коленного рефлекса (1,84 Нм), 3 и 4 – поверхностное отведение с m. vastus lateralis и с m. vastus medialis (200 мкВ и 500 мкВ соответственно). В скобках указана калибровка по размеру “Е”.
У больных при вызове рефлекса часто наблюдались вспышки активности клонического типа (3-й трек, рис. 21). У всех больных при бльших энергиях удара возникали также перекрестные реакции в форме тонической активности в мышцах бедра контрлатеральной ноги.

Рис.21.Коленный рефлекс в левой ноге у больной ДЦП 16 лет (спастический па-рапарез) на удар молоточком с энергией 4,07 мДж. 1 – поверхностное отведение с m. biceps femoris (100 мкВ), 2 – механо-грамма коленного рефлекса (3,7 Нм), 3 – поверхностное отведение с m. rectus fe-moris (200мкВ). В скобках указана калиб-ровка по размеру “Е”.

Подобные особенности рефлекторной активности наблюдались и у здоровых испытуемых при выполнении приема Ендрассика с максимальным усилием (рис. 22).
Рис.22. Коленный рефлекс у здорового испы-туемого 27 лет на удар молоточком по сухожи-лию левой ноги с энергией 76 мДж при выполне-нии приема Ендрассика. 1 и 4 – поверхностное отведение с m. biceps femoris правой и левой ноги (40 и 160 мкВ соответственно), 2 и 5 – механо-граммы моторных ответов в правой и в левой но-ге (0,82 и 2,49 Нм соответственно), 3 и 6 – повер-хностное отведение с m. rectus femoris правой и левой ноги (50 и 200 мкВ соответственно). В скобках указана калибровка по размеру "Е".
“Насыщение” рефлекторных путей. В отличие от здоровых людей, у которых при увеличении энергии стимуляции амплитуда коленного рефлекса нарастала и, достигнув максимума, затем снижалась (рис. 12 и 13), у тяжелых больных ДЦП амплитуда коленного рефлекса достигала максимума уже при энергиях удара 20 - 40 мДж (рис. 23) и при дальнейшем увеличении энергии удара не изменялась.

Рис.23. Величина коленного рефлекса и коэффи-циента вариации в зависимости от энергии удара молоточком у больной ДЦП (спастический парапарез). Каждая точка получена усреднением не менее 10 проб. По левой оси ординат – амплитуда коленных рефлексов, по правой оси ординат – коэффициенты вариации. По оси абсцисс – энергия удара молоточка. Вертикальные линии соответствуют величине гене-рального стандартного. Малые кружки – коленный рефлекс в правой ноге, малые квадратики – в левой ноге. Большие кружки – коэффициент вариации при вызове рефлекса в правой ноге, треугольники – в ле-
вой ноге.
График “стимул-ответ”, достигнув уровня плато, оставался параллельным оси абсцисс. Это явление, похожее на “насыщение” в опосредующих рефлекс нейронных путях, вероятно, являлось результатом активации большинства -мотонейронов уже при малых энергиях ударов по сухожилию. По этой же причине, вследствие высокого фонового супраспинального возбуждения, вызывающего “насыщение”, снижались и коэффициенты вариации.
Механизм облегчения коленных рефлексов. Инвазивные исследования механизма облегчения при приеме Ендрассика у здоровых испытуемых не обнаружили изменений в фоновых разрядах первичных и вторичных афферентов срединного, малоберцового и большеберцового нервов (Hagbarth et al.,1975). Эксперименты на животных также подтвердили, что повышение возбудимости в системе сухожильных рефлексов при этом не связано с активацией фузимоторной системы (Morgan et al, 1984). Это позволило предположить, что наиболее вероятным механизмом облегчения рефлексов при приеме Ендрассика является снижение порогов -мотонейронов, опосредуемое фоновой возбуждающей активностью. В пользу этого утверждения свидетельствует также то, что аналогичное облегчение сухожильных рефлексов наблюдается и при слабом произвольном сокращении тестируемой мышцы, то есть при произвольно вызванной активации -мотонейронов непосредственно этой же мышцы (Butler et al., 1993; Zehr, Stein, 1999; Mrachacz-Kersting, Sin-kjaer, 2003). По нашим наблюдениям, прием Ендрассика может вызывать активность и в тестируемой четырехглавой мышце, непосредственно повышая амплитуду рефлекторного ответа.
Резюме. Вызванная активность возникает в случае, когда испытуемый произвольно напрягает отдаленные мышцы (В.А. Сафронов, А.М. Эльнер, 1991; В.А. Сафронов, 2006) или произвольно активирует мышцу, в которой вызывается рефлекс. Подобного рода активность наблюдалась во многих мышцах при выполнении приема Ендрассика и при произвольном сокращении других мышц (А.М. Эльнер, В.А. Сафронов, 1994). Тот факт, что сумма вызванной активности многих мышц коррелировала с величиной облегчения сухожильного рефлекса, позволил предположить, что источник вызванной активности и фоновых облегчающих влияний един. Это заключение подтверждает также и наблюдавшееся нами облегчение рефлексов при попытках больного сокращать мышцы обездвиженной (паретичной) руки. Повышение фоновой активности, кроме приема Ендрассика, вызывают воздействия звука и света.
Уменьшение фоновой активности было отмечено нами во время дремотного состояния испытуемых, при ишемии исследуемой конечности, а также у больных с параличами и парезами периферического и центрального генеза различных уровней, у которых моторная активность бы-ла снижена. При заболевании ДЦП, напротив, имело место повышенное супраспинальное фоновое возбуждение, которое активирует многие спинальные нейронные пути. При этом, как указывали Л.В. Антонова и А.Л. Куренков (1999, с. 304), “часто отмечается активация антагонистов (ко-контракция) и отдаленных мышц, не имеющих к данному движению никакого отношения (патологические синергии)”. Одновременно имеет место и выраженное повышение сухожильных рефлексов.
Вызванную приемом Ендрассика сопутствующую активность у здоровых людей можно считать “моделью” патологической моторной гиперактивности у больных ДЦП. Патологический супрасегментарный фон, вероятнее всего, отличается от нормальных условий только количественно и не имеет избирательных влияний на спинальные структуры. Поэтому искусственное повышение супрасегментарного фона приближает моторный паттерн периферической активности у здоровых людей к особенностям нарушений в моторике у неврологических больных, опосредуемым повышенной нисходящей активностью.
Обсуждение результатов
Результаты проведенных исследований свидетельствуют о том, что стохастическая активность играет важную роль в организации движений. Особенно значимо её участие в поддержании мышечного тонуса, как осуществляемое непосредственно через нервно-мышечные механизмы, так и опосредованно через механизм РУ. Представление о том, что исходная стохастическая активность мотонейронов у человека является фактором в формировании мышечного тонуса разделялось многими исследователями (Rudomin, Dutton, 1967, 1969–a,-b; Chang et al., 1994; Gossard et al., 1994; Matthews, 1996).
В организации РУ принимают участие три фактора: фоновая возбуждающая активность пула тонических мотонейронов, аутогенное торможение этих мотонейронов афферентами сухожильных рецепторов и реология мышцы. Ведущим фактором является фоновая (надпороговая или подпороговая) возбуждающая активность пула тонических мотонейронов.
Выполненные исследования показали, что РУ обладает свойством суммации возбуждения, характерным для спинальных рефлексов. Уменьшение времени возникновения РУ при последовательных циклических перемещениях звена конечности, а также возникновение двойных разрядов двигательных единиц («дуплетов»), выявленные в фазу укорочения, подтверждают наличие процесса накопления возбуждения на мотонейронах в фазу укорочения иннервируемой ими мышцы. Это заключение согласуется с выводом работы Л.П. Кудиной и Р.Э. Андреевой (2004), что дуплеты генерируются при достижении некоторой величины “накопленного” на мотонейронах возбуждения.
Обнаруженный нами факт корреляции облегчения коленного рефлекса при произвольном сжатии кистью динамометра не с активностью произвольно напрягаемых сгибателей, а с величиной суммарной сопутствующей активности других мышц тела, позволяет полагать, что с началом произвольной активности в кондиционирующих мышцах высшие моторные отделы ЦНС во время выполнения движения активируют многие скелетные мышцы тела, возможно, для поддержания позы.
Из результатов проведепнных исследований следует также, что основной причиной нестабильности рефлекторных реакций являются шумовые процессы, порождаемые флюктуацией активности в нейронах нервной системы, в моторной коре и в корково-спинальных путях. Это заключение совпадает с представлениями, развиваемыми рядом других исследователей (Ellaway et al., 1998; Manjarrez et al., 2000; Baldissera et al., 2002). Так, при регистрации внутриклеточных потенциалов Phillips (1959) и Creutzfeldt et al. (1966) установили, что под воздействием синаптической бомбардировки мембранный потенциал пирамидных нейронов непрерывно флюктуирует, разряжаясь потенциалом действия при превышении порогового уровня. Согласно Эвартсу (Evarts, 1965), стохастической активностью обладают, главным образом, нейроны пирамидного тракта, медленно проводящие аксоны которых относятся к тонической системе. Brouwer и Ashby (1992) определили источники шума в нисходящих корково-спинальных трактах. Наличие связи между корковой и корково-спинальной активностью выявили Doyon и Benali (2005). Тоническая активность, существующая в корковых моторных отделах, отличается наличием стохастической компоненты, которая создает высокую вариативность в рефлекторных ответах, зависящих от уровня бодрствования, от воздействий внешних раздражителей, таких как звук, свет, от степени внутренней напряженности, тревоги или эмоциональных переживаний. Все сказанное позволяет заключить, что основной вклад в шумовую активность мотонейронов вносят стохастические влияния преимущественно корковых уровней, а также, вероятно, мозжечковых, рубральных и ретикулярных систем.
Выводы
1. Разработаны устройства для исследования мышечных реакций на пас-сивные однократные и на колебательные движения конечности, а также для исследования коленных рефлексов в изометрическом режиме.
2. Реакция укорочения является тонической аутогенной реакцией; как и другие спинальные мышечные реакции она обнаруживает свойство суммации возбуждения и короткое латентное время.
3. Реакция укорочения является механизмом поддержания тонуса мышц при движениях и может быть индикатором уровня фоновой активности мотонейронов и относительной мерой её величины.
4. Предложена модель механизма реакции укорочения с учетом того, что облегчающие влияния как сухожильных рефлексов, так и реакции укорочения опосредуются тонической активностью.
5. В динамике развития облегчающих влияний на сухожильные рефлексы, регистрируемые при произвольном напряжении различных мышечных групп, выделяются четыре фазы: фаза преднастройки, динамическая фаза, зависящая от скорости нарастания напряжения кондиционирующих мышц, фаза умеренного облегчения, отражающая процесс поддержания усилия, и фаза супраспинальных облегчающих влияний, обусловливаемых, вероятно, процессом утомления.
6. Величина сухожильного рефлекса при дополнительном произвольном усилии (прием Ендрассика) коррелирует не с величиной активности кондиционирующих мышц, а с сопутствующей активностью мышц тела.
7. Изменение мышечного тонуса по спастическому типу характеризуется повышением реакции мышц на их растяжение с большой скоростью.
8. Активность, вызванная приемом Ендрассика у здоровых людей, может быть “моделью” моторной гиперактивности патологического генеза.
9. Совокупность результатов проведенных исследований указывает на то, что поддержание мышечного тонуса как при движениях, так и в статике опосредуется стохастической активностью ЦНС.
Список публикаций по теме диссертации
- Гурфинкель В.С., Сафронов В.А. Устройство для исследования тонуса мышц в динамике //Изобр., пром. образцы, тов. знаки. 1965. № 1. А.С. 177586.
- Гедельман И.Д., Сафронов В.А., Хомская У.Д. К вопросу о произвольном управлении величиной мышечного тонуса //Новые исследования в педагогических науках. 1966. T.8. C. 163 – 169.
- Гурфинкель В.С., Сафронов В.А. Способ исследования тонуса мышц в динамике //Изобр., промыш. образцы, тов. знаки. 1966. № 10. А.С.181777.
- Гурфинкель В.С., Сафронов В.А. Устройство для исследования тонуса мышц в динамике //Изобр., промыш. образцы, тов. знаки. 1966. № 10. А.С. 181778.
- Сафронов В.А. Устройство для исследования тонуса мышц в динамике //Изобр., промыш. образцы, тов. знаки. № 18. А.С. 219079.
- Сафронов В.А. Устройство для исследования тонуса мышц в динамике //Изобр., промыш. образцы, тов. знаки. 1968. № 30. А.С. 227499.
- Гурфинкель В.С., Сафронов В.А. Устройство для измерения моментов инерции различных частей тела человека //Изобр., промыш. образцы, тов. знаки. 1969. № 33. А.С. 255483.
- Винокурский С.А., Гинзбург Х.Б., Гурфинкель В.С., Сафронов В.А., Будник В.М. Устройство для изучения тонуса мышц в динамике //Изобр., промыш. образцы, тов. знаки. 1969. № 28. А.С. 314517.
- Сафронов В.А. О механографической регистрации мышечного тонуса //Вопр. Психологии. 1969. № 3. С. 156 – 160.
10.Сафронов В.А. Характеристика мышечного тонуса при паркинсонизме //Матер. Объед. Конф. Нейрохирургов, посвящ. 100-летию В.И. Ленина. Ростов н/Д. 1969. С. 282 – 284.
11.Сафронов В.А. К вопросу о регуляции мышечного тонуса //Биофизика. 1970-а. Т. 15. Bып. 6. С. 1101 - 1109.
12.Сафронов В.А. Мышечный тонус при паркинсонизме //Вопр. Нейрохир.
1970-б. № 3. С. 11 – 17.
13.Гурфинкель В.С., Сафронов В.А. Мозжечковая гипотония у человека. В кн.: «Материалы 2-го Всесоюзного Симпозиума». Л. 1971. С. 131 – 135.
14.Сафронов В.А. (Выступление на съезде нейрохирургов) //Первый всесоюзный съезд нейрохирургов. Москва. 1972. Т.5. С. 210 – 213.
15.Васин Н.Я., Сафронов В.А., Ильинский И.А. Клинический анализ с спектромеханомиографическим контролем эффективности лечения Л-Допа больных паркинсонизмом //Клиническое значение препарата Л-ДОПА. М. 1973. С. 91 – 102.
16.Сафронов В.А. Спектромеханографическое исследование мышечного тонуса при акинетической форме паркинсонизма //Ж. Вопросы нейрохирургии. 1974. N1. С. 29 - 34.
17.Васин Н.Я., Сафронов В.А., Ильинский И.А. О Л-дофатерапии паркинсонизма и ее соотношение с хирургическим методом лечения //Ж. Невропатологии и психиатрии. 1974. Вып.5. С. 701-706.
18.Сафронов В.А., Кандель Э.И. Рефлекс на укорочение (феномен Вестфаля) при деформирующей мышечной (торсионной) дистонии //Ж. Невропатол. Психиат. им. С.С. Корсакова. 1975. Т. 75. Вып.8. С. 1495 – 1500.
19.Safronov V.A. Comparison of mechanomyographic characteristics of spasticity and rigidity //III Symposium International de Posturographie. Abstracts. Paris. 23-26 september. 1975. P. 73.
20.Safronov V.A. Muscle reaction to shortening in nirmal man //Third internati-
onal symposium on motor control. Bulgaria. Albena. 26 – 29 May. 1976. P.60.
21.Сафронов В.А. Электромиографическое исследование реакции на укорочение у больных паркинсонизмом //Материалы 2-го Всесоюзного симпозиума по электромиографии. Тбилиси. 1976. С. 146 – 147.
22. Safronov V.A. Comparison of mechanomyographic characteristics of spasticity and rigidity //Agressologie. 1977. V. 18 A. P. 23 - 29.
23.Сафронов В.А. «Гамма-ригидность» при паркинсонизме //Ж. Невропатологии и психиатрии им. С.С.Корсакова. 1977. Т.77. Вып.11. С. 1673 –77.
24.Сафронов В.А., Васин Н.Я., Лесов Н.С. Влияние дентатотомии на мышечный тонус при детском церебральном параличе //Вопросы нейрохирургии 1978. N 1. С. 24 - 30.
25.Сафронов В.А., Васин Н.Я., Таламбум Е.А. Особенности мышечного тонуса при ригидности и спастичности //Вопросы нейрохирургии. 1978. № 5. С. 27 – 33.
26.Сафронов В.А. ![]() - ригидность при паркинсонизме //Ж. Невропатологии и психиатрии им. С.С. Корсакова. 1979. Т.79. С. 164 – 168.
- ригидность при паркинсонизме //Ж. Невропатологии и психиатрии им. С.С. Корсакова. 1979. Т.79. С. 164 – 168.
27.Safronov V.A. On the mechanism of the shortening reaction //Fourth International Symposium on motor control. Varna. 8 – 12 June. 1981. P. 74.
28.Сафронов В.А. О некоторых свойствах реакции мышцы на пассивное укорочение //Физиология человека. 1984. Т.10. N4. С. 647 - 654.
29.Safronov V.A., Shevelev I.N., Ragukas R.K. The shortening reaction under conditions of antagonist muscle denervation after partial brachial plexus palsy //Fifth Internat. Symp. on Motor Control. Varna. 10 – 15 June. 1985. P. 122.
30.Сафронов В.А., Шевелев И.Н. Способ определения функционального состояния нервно-мышечной системы при травматических поражениях периферических нервов //Изобр., пром. образцы, тов. знаки. 1988. № 43. А.С. 1438695.
31.Сафронов В.А., Морозова Т.Г. Нервно-мышечные реакции на пассивные движения руки у больных с реактивными состояниями //Ж. Невропа-
тологии и психиатрии им. С.С. Корсакова. 1988. Т.88. Вып.10. С. 41-46.
32.Сафронов В.А., Шевелев И.Н., Ражукас Р.К. Исследование двигательной функции при травматических поражениях плечевого сплетния //Вопр. Нейрохир. 1988. N5. С. 22 – 25.
33.Safronov V.A., Elner A.M. The mechanism of Tendon reflexes by voluntary tension in remote muscles //Sixth Internat. Symp. on Motor Control. Varna. 3 – 7 Jule. 1989. P. 74.
34.Safronov V.A., Elner A.M. The development of facilitation of the tendon reflex by Jendrassik manoeure //Sixth Internat. Symp. on Motor Control. Varna.
3 – 7 Jule. 1989. P. 181.
35.Safronov V.A., Shabalov V.A. The participation of subcortical structures in the regulation on muscle tonus in cerebral palsy patients //Sixth Internat. Symp. on Motor Control. Varna. 3 – 7 Jule. 1989. P. 182.
36.Сафронов В.А. Возможности механографического способа исследования мышечного тонуса у больных с различными поражениями ЦНС //Третий Тбил. Межд. Симп. «Функциональная нейрохирургия» 28 – 30 мая 1990г. С. 261 – 263.
37.Сафронов В.А. Устройство для исследования тонуса мышц в динамике //Изобр., промыш. образцы, тов. знаки. 1990. № 16. А.С. 1560095.
38.Сафронов В.А. Устройство для измерения объемов сегментов тела человека //Изобр., промыш. образцы, тов. знаки. 1990.№ 24. А.С.1574210.
39.Сафронов В.А. Способ определения реакции человека на комфортабельность сидения //Изобр., промыш. образцы, тов. знаки. 1990. № 39. А.С. 1600699.
40.Safronov V.A. Muscle tonus of human subjects. //9th European Congress on Neurosurfery. Moscow. USSR. 1991. June 23–28. S. 314.
41.Safronov V.A. The rigidity state of muscles //Constituent congress international society for pathophysiology. M. 1991. P. 59.
42.Сафронов В.А., Эльнер А.М. Анализ процессов облегчения сухожиль-ного рефлекса приемом Ендрассика //Физиология человека. 1991. Т. 17.
№ 2. С. 73 – 80.
43.Эльнер А.М., Сафронов В.А. Облегчение коленного сухожильного рефлекса при напряжении мышц кисти //Физиология человека. 1994. Т. 20. № 3. С. 123 – 129.
44.Сафронов В.А., Шевелев И.Н., Шабалов В.А., Степаненко А.Ю., Колокольчикова О.С. Рефлекторная дуга коленного рефлекса у больных детским церебральным параличём //1 Российский конгресс по патофизиологии. М. 1996. С. 39.
45.Сафронов В.А. Устройство для измерения и анализа мышечных реак-
ций человека //Изобр., промыш. образцы, тов. знаки. 1998. № 13. А.С.
2110213.
46.Сафронов В.А. Исследование коленных рефлексов в изометрических условиях на электромиографе фирмы “NICOLET” //Материалы VI Международного симпозиума “Современные минимально-инвазивные технологии”. СПб. 2001. С. 68 – 71.
47.Сафронов В.А., Шевелев И.Н. Предоперационное исследование сохранности нервов-невратизаторов //Вопросы Нейрохирургии им. Н.Н. Бурденко. 2005. № 3. С. 17 – 20.
48.Сафронов В.А. Методика исследования коленного рефлекса в изометрических условиях //Физиология человека. 2005. Т.31.№ 6.С.116 –119.
49.Сафронов В.А. Коленный рефлекс. (Сообщение 1-е). Исследование здоровых людей //Физиология человека. 2006-а. Т. 32. № 2. С. 39 - 42.
50.Сафронов В.А. Коленный рефлекс. (Сообщение 2-е). Сопутствующая активация //Физиология человека. 2006-б. Т. 32. № 4. С. 83 – 88.
51.Сафронов В.А. Коленный рефлекс. (Сообщение 3-е). Сравнение рефлексов при нормальной и патологической фоновой активации //Физио-логия человека. 2007. Т. 33. № 1. С. 87 – 91.
52.Сафронов В.А. Хаотичные и упорядочные процессы, опосредующие коленный рефлекс //Физиология человека. 2009-а. Т. 35. № 3. С. 53 – 63.
53.Сафронов В.А. Влияние темноты на коленные рефлексы //Физиология
человека. 2009-б. Т. 35. № 5. С. 79 - 82.
С чувством глубокого уважения и искренней признательности благодарю директора Института нейрохирургии, академика РАН и РАМН, заслуженного деятеля науки России, ДМН, профессора Александра Николаевича Коновалова за предоставленную мне возможность осуществить на базе Института нейрохирургии комплекс исследовательских работ.
Приношу глубокую признательность за ценную помощь, оказанную мне в процессе подготовки работы, д.м.н., заслуженному деятелю науки России, профессору Иван Николаевичу Шевелеву.
Выражаю особую признательность и искреннюю благодарность чл. корр. РАН, заслуженному деятелю наук России, д.м.н., профессору Инесе Бенедиктовне Козловской за бесценную научную консультативную и редакционную помощь при написании работы.
Бесконечно признателен д.б.н., главному научному сотруднику отдела сенсорной физиологии и профилактики ГНЦ РФ – ИМБП РАН – Алексею Алексеевичу Шипову за всестороннюю и многогранную помощь, оказанную в процессе подготовки и оформлении работы.
Глубоко признателен моему первому научному руководителю, академику РАН, профессору Виктору Семеновичу Гурфинкелю.