

Роль p2у – рецепторов в регуляции сократимости миокарда крыс в постнатальном онтогенезе
На правах рукописи
Анисимова Ирина Николаевна
РОЛЬ P2У – РЕЦЕПТОРОВ
В РЕГУЛЯЦИИ СОКРАТИМОСТИ МИОКАРДА КРЫС
В ПОСТНАТАЛЬНОМ ОНТОГЕНЕЗЕ
03.03.01-физиология
Автореферат
диссертации на соискание учёной степени
кандидата биологических наук
Казань – 2011
Работа выполнена на кафедре анатомии, физиологии и охраны здоровья человека государственного образовательного учреждения высшего профессионального образования «Татарский государственный гуманитарно-педагогический университет»
Научный руководитель: доктор биологических наук, профессор
Аникина Татьяна Андреевна
Официальные оппоненты: доктор биологических наук, профессор
Гайнутдинов Халил Латыпович
доктор биологических наук, профессор
Ситдикова Гузель Фаритовна
Ведущая организация: государственное образовательное учреждение высшего профессионального образования «Ульяновский государственный университет»
Защита состоится «15» ноября 2011г. в ___ часов на заседании Диссертационного совета Д 212.078.02. при ФГАОУВПО «Казанский (Приволжский) федеральный университет» по адресу: 420008, г. Казань, ул. Левобулачная, д. 44
С диссертацией можно ознакомиться в научной библиотеке им. Н.И. Лобачевского при ФГАОУВПО «Казанский (Приволжский) федеральный университет» по адресу: 420008, г. Казань, ул. Кремлевская, 35.
Электронная версия автореферата размещена на официальном сайте ФГАОУВПО «Казанский (Приволжский) федеральный университет» www.ksu.ru.
Автореферат разослан «___» ___________ 2011 г.
Ученый секретарь
диссертационного совета

д.м.н., профессор Зефиров Т.Л.
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность исследования
Внутриклеточные пуриновые и пиримидиновые нуклеотиды (АТФ и УТФ) выполняют роль источника энергии, участвуют в биосинтезе рибонуклеиновых кислот, обеспечивая жизнедеятельность клетки. Установлено, что АТФ и уридин 5'-трифосфорная кислота (УТФ) находятся в везикулах вместе с основным медиатором и участвуют в передаче нервных импульсов, выполняя функции котрансмиттера (Burnstock and Kennedy, 1986; Burnstock, 1989, 1990, 2006, 2008, 2009; Kugelgen, 1991).
Свое влияние АТФ и УТФ реализуют через P2X и P2У-рецепторы, разнообразие которых превышает все известные подтипы рецепторов для «классических» нейромедиаторов и гормонов (Зиганшин и др., 1999, 2007; Гиниатуллин, 1998; Гиниатуллин и др., 2005; Аникина, Ситдиков, 2011. Yang et al., 1994; Ralevic and Burnstock, 1998; Ralevic, 2008; Domingo et al., 2010). Пуриновые и пиридиновые нуклеотиды как сигнальные молекулы при взаимодействии с рецепторами осуществляют пролиферацию, дифференциацию, развитие, регенерацию и апоптоз клеток (Burnstock., 2009).
Присутствие внеклеточной УТФ в сердце считается доказанным, и несколько подтипов Р2У- рецепторов активируются пуриновыми и пиримидиновыми нуклеотидами (Vassort, 2001). Метаболизм пиримидиновых нуклеотидов в сердце, к которым относится УТФ, изучен мало. Известно, что содержание пиримидиновых нуклеотидов в сердце ниже по сравнению с пуриновыми нуклеотидами и соотношение АТФ и УТФ составляет 1:10 (Vassort, 2001) или 1:16 (Lazarowski, 2003). Показано, что уровень АТФ и УТФ в плазме значительно коррелирует между собой, что указывает на совместное освобождение этих нуклеотидов (Wihlborg et al., 2006).
В литературе показано разнонаправленное влияние УТФ на деятельность сердца. Отмечено снижение и увеличение силы сокращения, появление двойного эффекта сократимости миокарда при добавлении УТФ (Рычков, 2007; Froldi et al., 1995; Vassort, 2001; Kugelgen, 2006; Talasila, 2009).
Онтогенетический аспект пуринергической регуляции сердца рассматривался только на взрослых организмах и в период их неонатального развития (Webb et al., 1996; Bogdanov et al., 1998; Hansen et al., 1999; Cheung et al., 2003). Экспрессия Р2У-рецепторов определяется на 11-й день эмбрионального развития крыс в сердце, скелетной мускулатуре, периферической и центральной нервной системе, легких, печени, когда большинство органов еще не сформированы и функционально не активны (Cheung and Burnstock, 2003). Показано, что у крыс УТФ вызывает рост неонатальных кардиомиоцитов (Pham et al., 2003).
Многие аспекты функционирования Р2У-рецепторов на важнейших этапах раннего постнатального онтогенеза остаются малоизученными, а эффекты УТФ отличаются в различных тканях и на разных этапах онтогенеза (Vassort, 2001). Методами иммуногистохимии и ПЦР показано присутствие Р2У1,2,4,6,11,13,14 - подтипов рецепторов в целом сердце, на отдельных кардиомиоцитах (Webb et al., 1996; Vassort, 2001; Burnstock, 2009), сердечных миофибробластах (Zheng et al., 1998; Talasila et al., 2009), эндотелии и гладкомышечных стенках сосудов (Wang et al., 2002). Экспрессия Р2У-рецепторов в миокарде меняется в онтогенезе и усиливается для Р2У1,2,6- подтипа рецепторов (Webb et al., 1996).
Однако, нет данных, подтверждающих участие P2У – рецепторов в регуляции сократимости миокарда крыс на ранних этапах постнатального онтогенеза в условиях незрелости регуляторных влияний на сердце и разных этапов биологического созревания организма.
Актуальность проблемы возрастает и в связи с установленным кардиопротектерным влиянием АТФ и УТФ при гипоксии, инфаркте миокарда, аритмии, ишемии сердца, сопровождающихся гибелью клеток и выходом АТФ и УТФ во внеклеточную среду с последующей активацией Р2- пуринорецепторов. (Ninomiya et al., 2002b; Wee et al., 2007). АТФ и УТФ оказывают влияние на миофибробласты, которые играют важную роль в процессах восстановления миокарда при сердечно-сосудистых заболеваниях (Yitzhaki et al., 2005, 2006; Talasila et al., 2009).
Цель и задачи исследования
Цель: исследовать роль УТФ и разных подтипов P2У - рецепторов в регуляции сократимости миокарда крыс в постнатальном онтогенезе.
В соответствии с этой целью были сформулированы следующие задачи:
1. Изучить влияние УТФ разной концентрации на сократительную активность миокарда предсердий и желудочков у крыс 7-, 21- и 100-суточного возраста.
2. Определить влияние неселективного блокатора P2У - рецепторов PPADS на сократимость миокарда предсердий и желудочков крыс 7-, 21- и 100-суточного возраста
3. Изучить влияние неселективной блокады P2У - рецепторов PPADS на эффект действия УТФ у крыс 7-, 21- и 100-суточного возраста.
4. Определить влияние селективного блокатора реактива голубого-2 на сократимость миокарда у крыс 7-, 21- и 100-суточного возраста.
5. Изучить влияние селективной блокады Р2У4 подтипа рецепторов реактивом голубым-2 на эффект действия УТФ у крыс 7-, 21- и 100-суточного возраста.
Основные положения диссертации, выносимые на защиту:
1. УТФ оказывает отрицательный инотропный эффект на миокард предсердий и желудочков крыс 7-, 21- и 100-суточного возраста. В наибольшей степени ингибирующий эффект УТФ на сократимость миокарда наблюдается у 21-суточных крысят.
2. Ингибирующий инотропный эффект УТФ у 7-, 21-суточных животных реализуется через P2У4 рецепторы, а у 100-суточных животных через P2У4 и P2У2 рецепторы.
Научная новизна
В диссертационной работе впервые проведено изучение влияния УТФ на сократимость миокарда крыс в раннем постнатальном онтогенезе. В работе впервые было установлено, что агонист P2У2,4,6 подтипов рецепторов УТФ вызывает уменьшение силы, скорости сокращения и расслабления миокарда предсердий и желудочков крыс 7-100-суточного возраста. Внеклеточная УТФ оказывает модулирующее влияние на сократимость миокарда в микро- и наномолярных концентрациях.
Получены новые данные о возрастных особенностях инотропии миокарда на действие УТФ. У 21-суточных крысят реакция силы, скорости сокращения и расслабления на влияние агониста больше по сравнению с новорожденными животными.
Впервые выявлено наличие собственного действия блокаторов P2У-рецепторов PPADS и реактива голубого-2 на амплитудно-временные характеристики сократимости миокарда крыс разного возраста. PPADS оказывает разнонаправленное действие на исследуемые параметры сократимости миокарда крыс 7-, 21 и 100-суточного возраста. У 21-суточных крысят в миокарде желудочков PPADS вызывает положительный инотропный и люзитропный эффекты в сократимости миокарда.
Реактив голубой-2 увеличивает амплитудно-временные характеристики сократимости миокарда всех возрастных групп, в большей степени у 21-суточных крысят.
Проведенные исследования с использованием разнообразных блокаторов P2У-рецепторов позволили установить подтипы рецепторов, участвующих в ингибирующем действии УТФ. У 100- суточных животных инотропная реакция миокарда на добавление УТФ реализуется при участии Р2У2 подтипа рецепторов.
Доказано, что P2У4 подтип рецепторов участвует в реализации отрицательного инотропного эффекта УТФ во всех возрастных группах и имеет более выраженное модулирующее влияние на сократимость миокарда крыс 100-суточного возраста.
Научно - практическая ценность
Полученные экспериментальные данные значительно расширяют и дополняют наши представления о пуринергической регуляции инотропной функции сердца, возрастных особенностях функциональной активности разных подтипов Р2У-рецепторов в различные периоды постнатального онтогенеза.
Полученные данные можно использовать при трактовке результатов исследований по физиологии сердца в зависимости от возраста животных. Результаты работы можно использовать при чтении лекций и факультативных курсов по физиологии. Материал исследований представляет интерес для специалистов по физиологии и кардиологии.
Личный вклад диссертанта
Приведенные в работе данные получены при личном участии соискателя на всех этапах работы, включая составление плана исследования, проведение экспериментов, обработку экспериментальных данных и оформление публикаций.
Достоверность полученных данных
Достоверность полученных данных подтверждалась использованием достаточного объема экспериментальных исследований, конкретной постановкой и решением поставленных задач и статистической обработкой полученных результатов.
Апробация работы
Основные результаты диссертационной работы доложены на следующих конференциях и форумах: VII Всероссийской конференции с международным участием «Механизмы функционирования висцеральных систем» (Санкт-Петербург, 2009); Международной конференции «Физиология развития человека» Института возрастной физиологии РАО (Москва, 2009); научных конференциях молодых ученых и специалистов ТГГПУ (Казань, 2009-2011); ХХI Съезде Физиологического Общества им. И.П. Павлова (Калуга, 2010); X Всероссийской научно-теоретической конференции «Физиологические механизмы адаптации растущего организма» (Казань, 2008, 2010); Всероссийской конференции с международным участием «Механизмы регуляции физиологических систем организма в процессе адаптации к условиям среды» (Санкт-Петербург, 2010); конференции молодых ученых «Механизмы адаптации физиологических систем организма к факторам среды» (Санкт-Петербург, 2010); XVIII Международной конференции студентов, аспирантов и молодых учёных «Ломоносов 2011» (Москва, 2011), на заседаниях кафедры анатомии, физиологии и охраны здоровья человека ТГГПУ (Казань, 2008-2011).
Реализация результатов исследования
По материалам диссертации опубликовано 18 научных работ, в том числе 2 статьи в рецензируемых журналах, рекомендованных ВАК.
Структура и объем диссертации
Диссертация объемом 114 страниц состоит из введения, обзора литературы, описания методики исследования, полученных результатов, их обсуждения, заключения, выводов и списка цитируемой литературы. Диссертация содержит 16 таблиц и 27 рисунков. Список цитируемой литературы включает 231 источников, из них 42 отечественных и 189 иностранных авторов.
МЕТОДЫ ИССЛЕДОВАНИЙ
Исследования проведены на 270 разнополых лабораторных беспородных белых крысах стадного разведения. Исследовали 3 возрастные группы животных: 7-, 21- и 100-суточного возраста.
Сократительную активность миокарда в эксперименте in vitro изучали на полосках предсердий и желудочков. Определение реакции сократительной функции миокарда на действие агонистов проводили на установке "PowerLab" ("ADInstruments") с датчиком силы "MLT 050/D" ("ADInstruments"). Запись кривой регистрировали на персональном компьютере при помощи программного обеспечения "Chart 5.0". Рассчитывали абсолютные значения силы сокращения в граммах и общую длительность изометрического сокращения полосок миокарда в секундах, а также оценивали процент изменения на действие агониста в процентах от исходных показателей, которые принимали за 100%.
Из правого желудочка и предсердия при помощи специальных ножниц и пинцетов препарировали полоски. Длина полосок составляла 1,5-2 мм, ширина не превышала 1мм. Препарат фиксировали вертикально одним концом к датчику силы, другим — к точке опоры, затем каждый препарат погружали в отдельный резервуар объемом 5 мл, в который подавался рабочий раствор. Состав раствора Кребса (в моль/л): NaСl- 8г; KCl- 0,3г; CaCl2- - 3 мл; MgSO4 –0,5 мл; NaH2PO4- 0,04г; глюкоза – 2г; Trizma HCl- 2.4-3.9 моль/л; Trizma base- 0.25 моль/л (Sigma). Раствор постоянно аэрировали карбогеном 95% O2 и 5% CO2, рН поддерживали в пределах 7.3-7.4.
После погружения препаратов в резервуары следовал "период приработки" в течение 40-60 мин, в ходе которой мышечным полоскам постепенно придавалось оптимальное натяжение. По окончании приработки 10 мин регистрировали исходные параметры сокращения. Затем в течение 30 мин. Регистрировали сократимость миокарда при добавлении в рабочий раствор агониста одной из концентраций. Агонист Р2У-рецепторов УТФ добавляли в различных концентрациях и оценивали изменения сокращений полосок миокарда крыс. По окончании стимуляции агонистами препараты трехкратно отмывали рабочим раствором в течение 10 мин, затем регистрировали исходные показатели для каждой последующей дозы. Для изучения влияния блокаторов Р2У – рецепторов PPADS и реактива голубого-2 оценивали изменения сокращений полосок миокарда в течении 1 часа. Влияние УТФ на амплитуду сокращения при блокаде Р2У – рецепторов записывали в присутствии PPADS или реактива голубого-2, затем добавляли на 20 минуте УТФ и записывали 30 минут.
РЕЗУЛЬТАТЫ СОБСТВЕННЫХ ИССЛЕДОВАНИЙ
Влияние УТФ на сократимость миокарда крыс в постнатальном онтогенезе.
У каждого подтипа Р2-рецепторов существуют наиболее и наименее чувствительные к нему агонисты. Известно, что УТФ является агонистом P2У2,4,6 подтипов пуринорецепторов.
Определение концентрации внеклеточной УТФ показало, что в сердце содержание УТФ в 10-16 раз меньше, чем АТФ (Vassort, 2001; Lazarowski et al., 2003). Известно, что концентрация внеклеточной АТФ соответствует микромолярной, следовательно, содержание внеклеточной УТФ составляет 10µМ. Поэтому для изучения дозозависимого эффекта мы выбрали диапазон концентраций УТФ от 10-6 до 10-10М.
Экзогенная УТФ в концентрации 10-10-10-6М вызывает отрицательный инотропный эффект у 7-, 21- и 100- суточных животных, как в предсердиях, так и в желудочках (рис. 2).
У 7 –суточных крыс УТФ в концентрации 10-6М уменьшает силу сокращения миокарда на 5,9±1,4% в предсердиях и на 8,4±2,4% в желудочках, что является достоверным по отношению к контролю (р<0,05). В концентрации 10-7М при действии агониста сила сокращения миокарда уменьшается на 7,6±2,3% в предсердиях и на 6,9±1,3% в желудочках (р<0,05). УТФ в концентрации 10-8М в этом же возрасте снижает силу сокращения миокарда на 2,6±1,1% в предсердиях и на 6,0±1,1% в желудочках (р<0,05). УТФ в концентрации 10-10М снижает силу сокращения миокарда на 5,2±0,8% в предсердиях и на 6,3±1,3% в желудочках (р<0,05) (табл. 1А, 1Б).
У 21-суточных крысят УТФ в концентрации 10-10М уменьшает силу сокращения препарата на 19,4±2,3% (р<0,05) в предсердиях и на 22±3,4% в желудочках по сравнению с исходной (р<0,05). Агонист в концентрации 10-8М, 10-7М и 10-6М в предсердиях снижает силу сокращения миокарда на 17,3±2,2%, 20,0±1,7% (р<0,05) и 15,0±2,7% соответственно (р<0,05). УТФ в концентрации 10-8М и 10-7М в желудочках уменьшает сократительную активность миокарда на 17,7±2,7% и 13,4±2,1% соответственно (р<0,05). А концентрация 10-6М вызывает незначительные изменения силы сокращения (рис. 1) (табл. 1А, 1Б).
У 100- суточных крыс УТФ выше названных концентраций также приводит к развитию отрицательной инотропии. Максимальный сократительный эффект УТФ у взрослых крыс возникает при концентрациях 10-10М и 10-6М в предсердиях на 20,6±5,4% и 19,3±4,5% соответственно (р<0,05). В желудочках 10-6М снижает силу сокращения на 15,7±1,4 (р<0,05), а концентрация УТФ 10-10М в желудочках не изменяет силу сокращения. Агонист в концентрации 10-8М снижает силу сокращения в предсердиях на 13,0±1,4%, а в желудочках на 12,8±1,9% (р<0,05). УТФ в концентрации 10-7М уменьшает сократительную активность миокарда предсердий на 11±2,9%, а в желудочках на 15,4±1,5% (р<0,05) (табл. 1А, 1Б).
Таким образом, физиологические концентрации внеклеточной УТФ (10-8 и 10-7М) вызывают достоверно больший ингибирующий эффект в сократительной активности миокарда предсердий и желудочков крыс 21 – и 100- суточных животных, по сравнению с 7-суточными (р<0,001).

Рис.1. Влияние УТФ на силу сокращения миокарда предсердий и желудочков крыс 21-суточного возраста.
Примечание:* - достоверность по сравнению с исходным значением *р<0,05

Рис. 2. Влияние УТФ на силу сокращения миокарда крыс 21-суточного возраста
Таблица 1А
Влияние УТФ на амплитудно-временные показатели сократимости миокарда 7-, 21- и 100- суточных крыс
| Возраст | Концентрация агониста, М | Сила сокращения, исх. (г) | Сила сокращения, мах. (г) | Длительность сокращения, исх. (с) | Длительность сокращения, мaх. (с) | Время сокращения, исх. (с) | Время сокращения, мах. (с) | |
| 7 дней | УТФ 10-7 | предсердие | 0,181±0,063 | 0,169±0,060* | 1,413±0,106 | 1,398±0,101 | 0,479±0,015 | 0,467±0,014 |
| желудочек | 0,204±0,023 | 0,191±0,022* | 1,305±0,031 | 1,294±0,026 | 0,480±0,006 | 0,483± 0,003 | ||
| УТФ 10-8 | предсердие | 0,162±0,032 | 0,156±0,031* | 1,146±0,107 | 1,128 ±0,108 | 0,454±0,009 | 0,447±0,009 | |
| желудочек | 0,242±0,031 | 0,228±0,032* | 1,277±0,020 | 1,294±0,027 | 0,402±0,062 | 0,392±0,060 | ||
| 21 день | УТФ 10-7 | предсердие | 0,077±0,021 | 0,063±0,018* | 0,683±0,113 | 0,685±0,118 | 0,322±0,034 | 0,348±0,053 |
| желудочек | 0,091±0,023 | 0,079±0,021* | 0,909±0,106 | 0,896±0,104 | 0,397±0,012 | 0,388±0,009 | ||
| УТФ 10-8 | предсердие | 0,172±0,067 | 0,147±0,060* | 0,696±0,084 | 0,703±0,088 | 0,335±0,028 | 0,343±0,028 | |
| желудочек | 0,130±0,042 | 0,109±0,036* | 0,922±0,105 | 0,902±0,103 | 0,416±0,024 | 0,339±0,021 | ||
| 100 дней | УТФ 10-7 | предсердие | 0,128±0,023 | 0,116±0,022* | 0,678±0,082 | 0,668±0,080 | 0,328±0,10 | 0,318±0,008 |
| желудочек | 0,109±0,019 | 0,094±0,018* | 1,224±0,229 | 1,193±0,224 | 0,425±0,021 | 0,417±0,019 | ||
| УТФ 10-8 | предсердие | 0,170±0,024 | 0,148±0,022* | 0,601±0,005 | 0,597±0,002 | 0,312±0,004 | 0,308±0,002 | |
| желудочек | 0,155±0,037 | 0,134±0,032* | 1,300±0,109 | 1,260±0,102 | 0,423±0,007 | 0,413±0,006 |
Примечание: * - достоверность по сравнению с исходным значением: р<0,05
Таблица 1Б
Влияние УТФ на амплитудно-временные показатели сократимости миокарда 7-, 21- и 100- суточных крыс
| Возраст | Концентрация агониста, М | Время расслабления, исх. (с) | Время расслабления, мах. (с) | Скорость сокращения, исх. (с/t) | Скорость сокращения, мах. (с/t) | Скорость расслабления, исх. (с/t) | Скорость расслабления, мах. (с/t) | |
| 7 дней | УТФ 10-7 | предсердие | 0,948±0,192 | 0,929±0,083 | 0,192±0,034 | 0,186±0,033* | 0,389±0,109 | 0,355±0,100* |
| желудочек | 0,826±0,024 | 0,825±0,025 | 0,219±0,031 | 0,202±0,027* | 0,407±0,050 | 0,380±0,045* | ||
| УТФ 10-8 | предсердие | 0,834±0,017 | 0,825±0,016 | 0,172±0,023 | 0,155±0,022* | 0,288±0,027 | 0,258±0,024* | |
| желудочек | 0,630±0,149 | 0,620±0,146 | 0,195±0,034 | 0,183±0,032* | 0,353±0,064 | 0,333±0,058* | ||
| 21 день | УТФ 10-7 | предсердие | 0,376±0,096 | 0,377±0,098 | 0,146±0,016 | 0,128±0,016* | 0,161±0,021 | 0,131±0,018* |
| желудочек | 0,487±0,112 | 0,473±0,107 | 0,187±0,023 | 0,168±0,020* | 0,212±0,030 | 0,192±0,026* | ||
| УТФ 10-8 | предсердие | 0,451±0,099 | 0,442±0,095 | 0,264±0,092 | 0,232±0,089* | 0,290±0,092 | 0,244±0,072* | |
| желудочек | 0,569±0,093 | 0,546±0,086 | 0,135±0,020 | 0,115±0,018* | 0,169±0,017 | 0,137±0,013* | ||
| 100 дней | УТФ 10-7 | предсердие | 0,355±0,072 | 0,353±0,071 | 0,541±0,079 | 0,472±0,072* | 0,522±0,058 | 0,477±0,056* |
| желудочек | 0,799±0,210 | 0,769±0,201 | 0,205±0,051 | 0,181±0,047 | 0,317±0,070 | 0,287±0,070 | ||
| УТФ 10-8 | предсердие | 0,289±0,001 | 0,288±0,001 | 0,527±0,070 | 0,464±0,053* | 0,488±0,062 | 0,430±0,049* | |
| желудочек | 0,877±0,104 | 0,853±0,099 | 0,272±0,058 | 0,258±0,054* | 0,517±0,102 | 0,448±0,079* |
Примечание: * - достоверность по сравнению с исходным значением: р<0,05
Влияние PPADS на амплитудно-временные характеристики сократимости миокарда крыс 7-100 суточного возраста.
Фармакологические исследования с использованием различных агонистов и антагонистов рецепторов позволят идентифицировать подтип P2У - рецепторов.
Существуют противоречивые данные о селективности блокаторов P2У-рецепторов. По мнению многих исследователей, блокаторы P2У - рецепторов обладают видовой и тканевой специфичностью. В настоящее время установлено, что PPADS является антагонистом Р2У1 и возможно, P2У6 подтипов рецепторов (Kugelgen et al., 2000, Kugelgen, 2006; Wee et al., 2007). В литературе имеются данные, где показано отсутствие влияния PPADS на P2У4 подтип рецепторов крыс (Collo et al., 1996; Townsend-Nicholson et al., 1999; Wildman et al., 2003; Kugelgen, 2000, 2006). По данным других авторов PPADS умеренно блокирует P2У2,6,11,13 подтипы рецепторов (Vassort, 2001; Kugelgen, 2005).
Изучали влияние PPADS на параметры сократимости миокарда предсердий и желудочков крыс 7-, 21- и 100-суточного возраста. Блокатор добавляли в концентрации 30µМ (Burnstock, 2006).
Регистрацию действия препарата на амплитудно-временные характеристики сократимости миокарда изучали в течение 60 минут. Стабилизация показателей сократимости после добавления блокатора наступает к 10 минуте и не меняется в последующие 50 минут.
У новорожденных крысят добавление PPADS вызывает положительный инотропный эффект, который в предсердиях составляет 7,3± 0,7%, а в желудочках 5,3± 1,0% (р<0,05).
У 21-суточных крыс в предсердиях исследуемый блокатор снижает амплитуду сокращения на 7,4±1,4% (р<0,05). После инкубации миокарда желудочков с PPADS наблюдается увеличение амплитуды сокращения на 47,2±4,7% (p<0,05) по сравнению с исходной. Из исследуемых временных параметров инотропии миокарда, в большей степени увеличиваются скорость сокращения и скорость расслабления миокарда желудочков на 44,2±3,5% и 40,8±8,9 соответственно (р<0,05). Такие показатели как длительность, время сокращения и расслабления изменяются не значительно.
У взрослых крыс блокатор оказывает отрицательный инотропноый эффект. В предсердиях сила сокращения уменьшается на 8,9±1,7%, а в желудочках на 6,8±1,1% по сравнению с исходной (р<0,05) (рис.3).

Рис.3. Влияние PPADS на силу сокращения миокарда предсердий и желудочков крыс 7-, 21- и 100 - суточного возраста
Примечание:* - достоверность по сравнению с исходным значением *р<0,05

Рис. 4. Влияние PPADS на амплитудно-временные характеристики сокращения миокарда желудочков крыс 21-суточного возраста.
Следовательно, PPADS оказывает разнонаправленное действие на параметры инотропии миокарда крыс разного возраста. У новорожденных крысят, как в предсердиях, так и в желудочках блокатор оказывает положительный инотропный эффект. У 21-суточных крыс антагонист приводит к развитию положительного инотропного и люзитропного действия на миокард желудочков. У 100- суточных животных наблюдается уменьшение силы сокращения миокарда предсердий и желудочков.
Влияние УТФ на фоне PPADS на сократимость миокарда крыс 7-, 21- и 100-суточного возраста.
Для определения подтипа P2У-рецепторов, участвующих в эффектах УТФ провели серию экспериментов по определению влияния УТФ на фоне блокатора РРАDS. Исследовали влияние наиболее эффективных концентраций УТФ на силу сокращения миокарда. Для 7- и 100-суточных крысят добавляли УТФ в концентрации 10-8М, а для 21-суточных - 10-7М.
У новорожденных крысят в контроле агонист оказывает отрицательный инотропный эффект, который составляет в предсердиях 7,0±0,8%, а в желудочках - 4,3± 0,7%. На фоне блокады P2У2,6 подтипов рецепторов отрицательный инотропный эффект УТФ сохраняется. В предсердиях он составляет 4,9±0,8%, в желудочках 2,3±0,5%.
У 21-суточных крысят УТФ в контроле снижает силу сокращения миокарда предсердий на 15,0±1,4%, а в желудочках на 15,7±0,8%. После инфузии с антагонистом добавление УТФ не изменяет амплитуду сокращения миокарда (рис. 5).
У 100-суточных крыс в контроле УТФ уменьшает силу сокращения желудочков на 14,5±1,7%, и предсердий на 16,6±0,8%. При блокаде Р2У2,6 подтипов рецепторов в предсердиях ингибирующий эффект УТФ также сохраняется (рис.6).
Таким образом, при блокаде Р2У2,6 подтипов рецепторов отрицательный эффект УТФ сохраняется в миокарде предсердий и желудочков всех возрастных групп. Однако, у 100- суточных крыс блокатор РРАDS снижает эффект УТФ в миокарде желудочков с 16 до 6% (р<0,05). Следовательно, ингибирующий эффект УТФ у 100-суточных животных в миокарде желудочков реализуется через P2У2 подтип рецепторов.

Рис. 5. Влияние УТФ в контроле и при блокаде PPADS на силу сокращения миокарда предсердий и желудочков у 21- суточных животных.
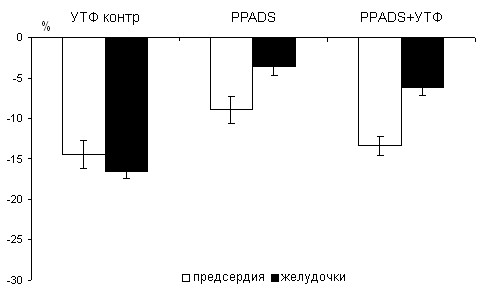
Рис. 6. Влияние УТФ в контроле и при блокаде PPADS на силу сокращения миокарда предсердий и желудочков у 100- суточных животных.
Влияние реактива голубого-2 на сократимость миокарда крыс в онтогенезе. Известно, что у крыс реактив голубой-2 блокирует P2У4 подтип рецепторов в сердце (Bogdanov et al., 1998; Wildman et al., 2003). У крыс этот подтип рецепторов не чувствителен к сурамину и PPADS (Burnstock, 2009). По данным исследования Wee и его сотрудников реактив голубой-2 является эффективным антагонистом, как при низких, так и высоких концентрациях. Реактив голубой -2 добавляли в концентрации 15 микроМ (Burnstock, 2006; Wee et al., 2007).
Реактив голубой является селективным блокатором для P2У4 подтипа рецепторов в сочетании с УТФ как агонистом этих рецепторов.
В данной серии экспериментов изучали влияние реактива голубого-2 на силу сокращения предсердий и желудочков всех возрастных групп. Инфузия блокатора вызывает положительный инотропный эффект миокарда у 7-100-суточных животных. Увеличение силы сокращения изолированных полосок миокарда на действие препарата происходит постепенно, и стабилизация исследуемых параметров сократимости наблюдается к 15-20 минуте.
У 7-суточных крысят реактив голубой-2 вызывает положительный инотропный эффект в предсердиях на 16,3±5,1%, а в желудочках на 13,3±2,3% (р<0,05). У 21-суточных животных наибольший эффект на добавление реактива голубого-2 наблюдался в желудочках и составил 20,0+3,9% (р<0,05).В предсердиях он достигал 15,9+2.6% (р<0,05). У взрослых крыс антагонист также увеличивает силу сокращения миокарда, но в меньшей степени. В предсердиях на 5,9±1,13% и в желудочках на 6,0±1,39% по сравнению с исходной (p<0,05) (рис. 7).
Изучение влияния реактива голубого-2 на сократительную активность миокарда крыс выявило возрастные особенности. Так, у 7- и 21-суточных крысят сила сокращения полосок миокарда предсердий и желудочков на действие блокатора достоверно выше, чем у 100-суточных животных (p<0,05).

Рис. 7. Влияние реактива голубого-2 на силу сокращения миокарда предсердий и желудочков крыс 7-, 21- и 100 - суточного возраста
Примечание:* - достоверность по сравнению с исходным значением *р<0,05
Влияние УТФ на фоне реактива голубого-2 на сократимость миокарда крыс 7-, 21- и 100-суточного возраста.
Для определения подтипа рецепторов, участвующих в ингибирующих эффектах УТФ провели серию экспериментов по изучению влияния УТФ при блокаде P2У4 подтипа рецепторов реактивом голубым-2.
Для эксперимента выбрали такие концентрации УТФ, которые вызывают достоверно значимые изменения показателей сократимости, и соответствуют содержанию внеклеточной УТФ.
У 7-суточных крысят в контроле УТФ уменьшает силу сокращений предсердий на 4,6±0,8%, а желудочков на 6,4+1,2% (р<0,05). При блокаде P2У4 подтипа рецепторов отрицательный инотропный эффект УТФ не сохраняется (р<0,05) (рис.8А, 8Б).
У 21-суточных животных УТФ в контроле сила сокращения снижается в предсердиях на 29,1±4,8% (р<0,05), в желудочках на 22,7±2,9% (р<0,05). При блокаде P2У4 подтипа рецепторов ингибирующий эффект УТФ также не сохраняется (р<0,05) (рис. 9А, 9Б).
У 100-суточных крыс на полосках миокарда предсердий сила сокращения уменьшилась на 15,9+3,5% (р<0,05), а желудочков на 20,3+3,7% (р<0,05). При блокаде P2Y4 подтипа рецепторов ингибирующий эффект УТФ также не сохраняется (р<0,05). Введение УТФ на фоне предварительного добавления реактива голубого-2 сохраняет положительный инотропный эффект блокатора (р<0,05) (рис. 10).
Таким образом, у 7-, 21- и 100- суточных животных после инкубации миокарда с реактивом голубым-2 отрицательный инотропный эффект УТФ не сохраняется.
У 7- и 21-суточных крысят на фоне блокады реактива голубого-2 введение УТФ приводит к некоторому снижению силы сокращения миокарда предсердий и желудочков, которое однако не достигает контрольных значений УТФ (р<0,05).
Таким образом, реализация ингибирующего эффекта УТФ в сократимости миокарда осуществляется через Р2У4-подтип рецепторов у крыс 7-, 21- и 100-суточного возраста. У 100-суточных животных преобладают влияния УТФ с участием P2У4 подтипов рецепторов.
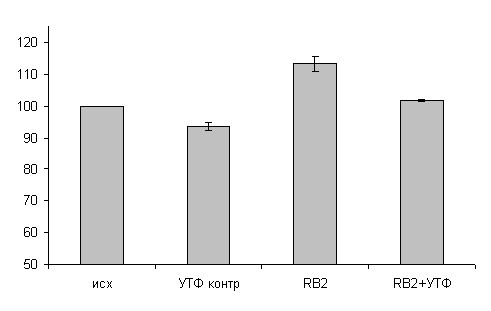

Рис.8А. Влияние УТФ на силу сокращения миокарда желудочков 7- суточных животных в контроле и при блокаде реактивом голубым-2.
Рис. 8Б. Реакция силы сокращения миокарда желудочков на действие УТФ в контроле и при блокаде реактивом голубым-2 у 7-суточных животных.

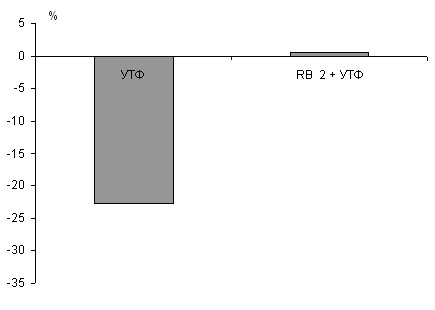
Рис.9А. Влияние УТФ на силу сокращения миокарда желудочков 21- суточных животных в контроле и при блокаде реактивом голубым-2.
Рис. 9Б. Реакция силы сокращения миокарда желудочков на действие УТФ в контроле и при блокаде реактивом голубым-2 у 21-суточных животных.
Примечание:* - достоверность по сравнению с контролем *р<0,05,

Рис.10. Влияние УТФ на силу сокращения миокарда желудочков в контроле и при блокаде реактивом голубым-2 у 100- суточных животных.
ВЫВОДЫ
1. УТФ в концентрации 10-6-10-10М оказывает отрицательный инотропный эффект на миокард предсердий и желудочков крыс 7-, 21- и 100-суточного возраста.
2. Реакция миокарда предсердий и желудочков на УТФ у 21-суточных крысят более выражена по сравнению с новорожденными.
3. УТФ в концентрации 10-8М у 7- и 100-суточных и в концентрации 10-7М у 21-суточных уменьшает скорость сокращения и расслабления миокарда предсердий и желудочков.
4. PPADS вызывает разнонаправленное действие на силу сокращения миокарда крыс разного возраста. У 7-суточных крысят блокада Р2У - рецепторов увеличивает, у 100- суточных животных снижает силу сокращения миокарда, у 21-суточных крыс приводит к развитию положительного инотропного и люзитропного эффекта в миокарде желудочков.
5. PPADS не оказывает влияния на отрицательный инотропный эффект УТФ в миокарде предсердий и желудочков всех возрастных групп. У 100-суточных животных PPADS снижает реакцию сократимости миокарда желудочков на УТФ.
6. Реактив голубой-2 оказывает положительный инотропный эффект на миокард предсердий и желудочков 7-, 21- и 100-суточных животных. Реакция сократимости миокарда на антагонист наиболее выражена у 21-суточных крысят.
7. Реактив голубой-2 предотвращает ингибирующий эффект УТФ на сократимость миокарда предсердий и желудочков 100-суточных животных и уменьшает его у 7-и 21-суточных крысят.
Список работ, опубликованных по теме диссертации
- Ситдиков Ф.Г., Аникина Т.А., Трофимова И.Н. Возрастные особенности влияния УТФ на сократительную активность миокарда крыс в раннем постнатальном онтогенезе. Вестник ТГГПУ, №3 (21), 2010.- С. 111-115.
- Анисимова И.Н., Аникина Т.А., Ситдиков Ф.Г. Влияние блокады P2У- пуринорецепторов на инотропную функцию сердца в онтогенезе. Вестник ТГГПУ, 2011, № 3 (25).- С. 41-45.
- Аникина Т.А., Зверев А.А., Ситдиков Ф.Г., Трофимова И.Н. АТФ модулирует холинергические влияния на сократимость миокарда в зависимости от возраста крыс. IХ Всероссийская конференция «Физиологические механизмы адаптации растущего организма», посвященная юбилею проф. Ф.Г. Ситдикова., 2008, с. 17-18
- Аникина Т.А., Трофимова И.Н. Влияние УТФ на сократимость миокарда крыс. Вестник ТГГПУ, 2008, № 4 (15), с. 58-60
- Аникина Т.А., Зверев А.А., Ситдиков Ф.Г., Трофимова И.Н. Пуринорецепторы сердца. Вестник ТГГПУ, 2008, № 4 (15), с. 53-57
- Ситдиков Ф.Г., Аникина Т.А., Зверев А.А., Трофимова И.Н. Участие P2X и P2У- рецепторов в сократимости миокарда крыс. Всероссийская научно-практическая конференция «Инновационные подходы к естественнонаучным исследованиям и образованию», посвященная юбилею ЕГФ, Казань, 2009, с. 21-23
- Аникина Т.А., Ситдиков Ф.Г., Зверев А.А., Трофимова И.Н. Пуринергическая регуляция сердца в постнатальном онтогенезе. Материалы международной конференции «Физиология развития человека». Москва.- 2009.- С. 8-9.
- Аникина Т.А., Ситдиков Ф.Г., Зверев А.А., Трофимова И.Н. Пуринорецепторы сердца крыс в онтогенезе. VII Всероссийская конференция с международным участием “Механизмы функционирования висцеральных систем”, посвященная 160-летию со дня рождения И.П. Павлова Санкт-Петербург, 2009, с.29-30
- Ситдиков Ф.Г., Аникина Т.А., Зверев А.А., Трофимова И.Н. Влияние УТФ на сократимость миокарда крыс в постнатальном онтогенезе. Тезисы Всероссийской научной конференции с международным участием «Теоретические основы физической культуры». Казань: ТГГПУ, 2009.- С. 155-157
- Трофимова И.Н. Влияние УТФ на сократимость миокарда крыс в раннем постнатальном онтогенезе. Сборник тезисов лучших докладов ХХХ научной конференции молодых ученых и специалистов, посвященный Году учителя в РФ и РТ «Молодые ученые в научном поиске: выпуск 1. Казань.- ТГГПУ.- С. 79-83.
- Трофимова И.Н., Аникина Т.А., Ситдиков Ф.Г., Константинова И.В. Участие P2У- пуринорецепторов в сократимости миокарда. Материалы Х юбилейной Всероссийской научной конференции с международным участием «Физиологические механизмы адаптации растущего организма».- Казань: ТГГПУ, 2010.- С. 183-185.
- Трофимова И.Н., Аникина Т.А. Влияние АТФ и УТФ на сократимость миокарда крыс в постнатальном онтогенезе. Тезисы докладов ХХI съезда физиологического общества им. И.П.Павлова.- М.-Калуга, 2010.- С. 619.
- Аникина Т.А., Ситдиков Ф.Г., Билалова Г.А., Трофимова И.Н. АТФ как модулятор эффектов классических медиаторов в постнатальном онтогенезе. Тезисы докладов ХХI съезда физиологического общества им. И.П.Павлова.- М.-Калуга, 2010.- С. 759-760.
- Аникина Т.А., Ситдиков Ф.Г., Трофимова И.Н. Участие P2У- пуринорецепторов в сократимости миокарда крыс в раннем постнатальном онтогенезе. Тезисы докладов Всероссийской конференции с международным участием «Механизмы регуляции физиологических систем организма в процессе адаптации к условиям среды», посвященной 85-летию со дня основания Института физиологии им. И.П. Павлова РАН. - Санкт-Петербург, 7–9 декабря 2010.- С. 16.
- Анисимова И.Н. Влияние уридин-5‘-трифосфата на сократимость миокарда крыс в постнатальном онтогенезе. Тезисы докладов Конференции молодых ученых, посвященной 85-летию со дня основания Института физиологии им. И. П. Павлова РАН «Механизмы адаптации физиологических систем организма к факторам среды».- СПб.: Институт физиологии им. И. П. Павлова РАН, 2010. – С. 7-8.
- Анисимова И.Н. Участие разных подтипов P2У-рецепторов в сократимости миокарда крыс в постнатальном онтогенезе. Тезисы докладов XVIII международной научной конференции студентов, аспирантов и молодых ученых «Ломоносов-2011».- Москва, МГУ.-2011.- с. 237-238.
- Аникина Т.А., Ситдиков Ф.Г., Анисимова И.Н., Лохотская Л.А. Роль пуринорецепторов в регуляции деятельности сердца крыс. Тезисы докладов III Съезда физиологов СНГ, Ялта, 2011.- С.
18. Аникина Т.А., Ситдиков Ф.Г., Зверев А.А., Анисимова И.Н. Пуринергическая регуляция сердца в онтогенезе. Учебное пособие “Избранные главы фундаментальной и восстановительной медицины”.- Казань.- 2011.- С.95-114.