

Влияние микробных и немикробных факторов на формирование внеклеточных ловушек нейтрофилами периферической крови
На правах рукописи
Рыжкова
Анна Ивановна
влияние микробных и немикробных факторов на формирование внеклеточных ловушек нейтрофилами периферической крови
14.03.09 – клиническая иммунология, аллергология
Автореферат
диссертации на соискание ученой степени
кандидата медицинских наук
Челябинск – 2010
Работа выполнена в Научно-исследовательском институте иммунологии Государственного образовательного учреждения высшего профессионального образования «Челябинская государственная медицинская академия Федерального агентства по здравоохранению и социальному развитию»
Научный руководитель:
заслуженный деятель науки РФ,
член-корреспондент РАМН,
доктор медицинских наук, профессор Долгушин Илья Ильич
Официальные оппоненты:
доктор медицинских наук, Базарный Владимир Викторович
профессор
доктор медицинских наук, Бурмистрова Александра Леонидовна
профессор
Ведущая организация: Институт иммунологии и физиологии УрО РАН, г. Екатеринбург
Защита состоится «_29_»_июня_____2010г. в ______ часов на заседании диссертационного совета Д 208.117.03. при Государственном образовательном учреждении высшего профессионального образования «Челябинская государственная медицинская академия Федерального агентства по здравоохранению и социальному развитию» по адресу: 454092, г. Челябинск, ул. Воровского, д. 64.
С диссертацией можно ознакомиться в библиотеке Государственного образовательного учреждения высшего профессионального образования «Челябинская государственная медицинская академия Федерального агентства по здравоохранению и социальному развитию».
Автореферат разослан «___» _______________20__ г.
Ученый секретарь
диссертационного совета
доктор медицинских наук, профессор Л.Ф. Телешева
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы
Более века исследователи всего мира с интересом изучают строение, физиологию нейтрофилов, биохимический состав их гранул, структуры синтезируемых ими метаболитов и роль этих клеток в регуляции иммунного и других звеньев гомеостаза [Хаитов Р.М., Пинегин Б.В., 1995; Тотолян А.А., Фрейдлин И.С., 2000; Долгушин И.И., Бухарин О.В., 2001; Пинегин Б.В., Маянский А.Н., 2007; Virchow R., 1897; Lehrer R.I. еt аl., 1988; Pabst M.J., 1994; Smolen J.E., 1995].
В 1908 году русский ученый И.И. Мечников получил Нобелевскую премию за открытие центральной роли фагоцитирующих клеток в неспецифической защите организма и указал на их бактерицидные свойства и способность продуцировать «секретины» [Мечников И.И., 1947].
Нейтрофилы обладают способностью захватывать бактерии и другие частицы с помощью специфических рецепторов или без их участия, убивать захваченные микроорганизмы с использованием кислородзависимых и кислороднезависимых механизмов и переваривать захваченные объекты фагоцитоза [Фрейдлин И.С., 1998]. До недавнего времени нейтрофилы рассматривались как исключительно неспецифические эффекторные клетки врожденного иммунитета, не имеющие возможность четко направлять комплекс иммунных реакций. Наиболее вероятным исходом дифференцированных нейтрофилов считался апоптоз после фагоцитоза инфекционных объектов или некроз, обусловленный, например, бактериальными токсинами.
В последнее время знание об иммунологическом репертуаре нейтрофилов значительно расширилось. Были обоснованы представления о важной роли бактерицидных продуктов, выделяемых этими клетками наружу, в противомикробной защите [Долгушин И.И., Бухарин О.В., 2001]. Brinkmann V. и соавторы в 2004 году сообщили, что нейтрофилы выбрасывают во внеклеточное пространство свой хроматин, формируя нейтрофильные внеклеточные ловушки (Neutrophil Extracellular Traps, NETs, НВЛ), состоящие из нуклеиновых кислот, гранулированных пептидов и антимикробных молекул, с ранее неизвестной целью – для изоляции и уничтожения Грам-положительных, Грам-отрицательных бактерий и грибов [Долгушин И.И., Андреева Ю.С., 2009; Brinkmann V., Reichard U., et al., 2004]. Эта кислородзависимая клеточная гибель была названа термином «NETosis» [Steinberg B.E., Grinstein S., 2007]. Позднее подобное явление было обнаружено у тучных клеток и эозинофилов [Von Kockritz-Blickwede M. еt аl., 2008]. Формирование нейтрофилами внеклеточных ловушек – важный механизм врожденного иммунного ответа, когда, погибая, нейтрофил защищает организм от инфекционных патогенов [Fuchs T.A. еt аl., 2007; Wartha F., Henriques-Normark B., 2008].
Однако до сих пор четких представлений о формировании нейтрофилами внеклеточных ловушек нет.
Цель исследования
Оценить влияние представителей нормальной и условно патогенной микрофлоры слизистых оболочек гениталий и кишечника человека (Bifidobacterium bifidum, Lactobacillus spp., Escherichia coli, Staphylococcus aureus, Candida albicans) и иммуномодуляторов различной природы (пирогенал, солкотриховак, циклоферон, интерферон), а также гуморальных факторов (комплемент и специфические Ig) на формирование внеклеточных ловушек нейтрофилами периферической крови.
Задачи исследования
- Определить влияние Bifidobacterium bifidum, Lactobacillus spp., E. coli, S. aureus и C. albicans на формирование внеклеточных ловушек нейтрофилами в цельной гепаринизированной крови, в лейкоцитарной взвеси и нейтрофилами, выделенными на двойном градиенте фиколла-верографина.
- Оценить роль плазмы, сыворотки крови и их компонентов (комплемента и специфических Ig) в секреции внеклеточной ДНК нейтрофилами, выделенными на двойном градиенте фиколла-верографина, в системе in vitro.
- Изучить воздействие пирогенала, солкотриховака, циклоферона и интерферона на образование внеклеточных ловушек нейтрофилами, выделенными на двойном градиенте фиколла-верографина.
Научная новизна
Впервые было проведено исследование образования внеклеточных ловушек нейтрофилами, выделенными из периферической крови на двойном градиенте фиколла-верографина, нейтрофилами цельной гепаринизированной крови, лейкоцитарной взвеси. При этом использовались различные методы обнаружения внеклеточно расположенной нейтрофильной ДНК: фиксированные мазки цельной крови, окрашенные по Романовскому-Гимзе, оценивались при световой иммерсионной микроскопии; фиксированные препараты выделенных нейтрофилов окрашивались раствором акридинового оранжевого или Sytox green и подвергались люминесцентной микроскопии; образцы, содержащие выделенные нейтрофилы и сыворотку или плазму крови, исследовались в нативном состоянии при люминесцентной микроскопии и окраске акридиновым оранжевым. Все эти методы позволили произвести количественную оценку НВЛ по отношению к другим морфологическим формам нейтрофилов (с сегментированным и с недифференцированным ядром).
На основании количественной оценки установлен дозозависимый эффект влияния различных видов микроорганизмов и биологически активных веществ на образование НВЛ. Представители нормальной и условно-патогенной микрофлоры слизистых оболочек гениталий и кишечника человека стимулируют образование внеклеточных ловушек нейтрофилами, выделенными на двойном градиенте фиколла-верографина, но не оказывают влияния на нейтрофилы цельной гепаринизированной крови, лейкоцитарной взвеси и нейтрофилы в присутствии сыворотки или плазмы крови, независимо от концентрации микроорганизмов. При сравнительном анализе установлено, что наиболее выраженный стимулирующий эффект на формирование НВЛ оказывают Candida albicans и Lactobacillus spp.
Сыворотка и плазма крови, независимо от присутствия в них специфических иммуноглобулинов и комплемента, оказывают ингибирующее действие на внеклеточный выброс ДНК нейтрофилами, выделенными из периферической крови на двойном градиенте фиколла-верографина.
Иммуностимуляторы различной природы (пирогенал, солкотриховак, интерферон и циклоферон) оказывают активирующее влияние на секрецию хроматина нейтрофилами, выделенными из периферической крови на двойном градиенте фиколла-верографина. Однако различные дозы иммуностимуляторов по-разному влияют на формирование НВЛ. Интерферон в высоких концентрациях ингибирует образование нейтрофильных внеклеточных ловушек. Пирогенал, солкотриховак и циклоферон активнее стимулируют внеклеточный выброс нейтрофильной ДНК в концентрациях, эквивалентных средней терапевтической дозе для каждого конкретного препарата.
На основании проведенного исследования был разработан метод оценки эффективности вакцины «Солкотриховак» [приоритетная справка «Способ оценки эффективности вакцин, участвующих в формировании мукозального иммунитета» № 2009102575 (003293) от 26.01.2009].
Теоретическая и практическая значимость работы
Проведенные исследования позволяют расширить представление о роли нейтрофилов в антимикробной защите организма, в частности их способности к образованию внеклеточных ДНК-овых сетей.
Полученные результаты дают возможность оценить дозозависимый эффект и селективность секреции хроматина во внеклеточное пространство нейтрофильными гранулоцитами в ответ на стимуляцию факторами микробной и немикробной природы и обосновывают возможность использования метода определения НВЛ для оценки иммунотропной активности различных препаратов.
Основные положения, выносимые на защиту
- Под действием представителей нормальной и условно-патогенной микрофлоры активируется секреция хроматина во внеклеточное пространство нейтрофилами, выделенными из периферической крови на двойном градиенте фиколла-верографина.
- Иммуномодуляторы оказывают стимулирующий дозозависимый эффект на образование нейтрофилами внеклеточных ловушек, при этом максимальное влияние оказывает средняя терапевтическая доза препаратов.
- Сыворотка и плазма крови блокируют выброс нитей ДНК нейтрофилами во внеклеточное пространство. Участия специфических IgG в данном процессе не выявлено. Аутологичный комплемент в присутствии сыворотки и плазмы крови не оказывает влияния на формирование НВЛ, однако комплемент морской свинки дозозависимо стимулирует секрецию ДНК нейтрофилами, выделенными из периферической крови на двойном градиенте фиколла-верографина, этот эффект снижается после термической денатурации комплемента.
Внедрение результатов исследования
Результаты исследований внедрены в лабораторную диагностику НИИ иммунологии и в учебный процесс на кафедре микробиологии, вирусологии, иммунологии ГОУ ВПО ЧелГМА Росздрава.
Апробация работы
Основные положения диссертации доложены, обсуждены и опубликованы на X – X Всероссийском научном Форуме имени академика В.И. Иоффе «Дни иммунологии в Санкт-Петербурге: Молекулярные основы иммунорегуляции, иммунодиагностики и иммунокоррекции» (г. Санкт-Петербург, 2008, 2009), V Российской научной конференции «Персистенция микроорганизмов» (Оренбург 2009), V итоговой научно-практической конференции молодых ученых ЧелГМА (Челябинск, 2009).
Публикации
По теме диссертации опубликовано 14 научных работ, из них в ведущих рецензируемых научных изданиях и журналах, рекомендованных ВАК – 10.
Структура и объем диссертации
Диссертационная работа состоит из введения, обзора литературы, материалов и методов исследования, 3-х глав собственных исследований, заключения, выводов и списка литературы, включающего работы 91 отечественных и 128 иностранных авторов. Диссертация изложена на 143 страницах, включает 32 таблицы и 8 рисунков.
СОДЕРЖАНИЕ РАБОТЫ
Материалы и методы исследования
Нами было обследовано 102 условно здоровые женщины в возрасте 18 – 35 лет в первой фазе менструального цикла.
В качестве объекта изучения были выбраны нейтрофилы цельной гепаринизированной крови, лейкоцитарной взвеси, полученной после седиментации эритроцитов, и нейтрофилы, выделенные из периферической крови на двойном градиенте фиколла-верографина. Исследовалась их способность к образованию НВЛ под влиянием факторов микробной и немикробной природы.
Для выделения нейтрофилов 2 мл гепаринизированной крови (10 ЕД гепарина («Гедеон-Рихтер», Hyngery) на 10 мл крови) смешивали с 3 мл стерильного физиологического раствора, полученную смесь наслаивали на градиент плотности стерильных растворов фиколла-верографина (Pharmacia, Sweden; Spofa, CSSR). Плотность верхнего слоя – 1,075-1,077, нижнего – 1,093-1,095 [Wong L., 1975], объем каждого слоя – 2 мл. Через 40 минут центрифугирования при 1500 об/мин на границе градиентов образовывалось кольцо гранулоцитов с чистотой 98 – 100%, мононуклеары составляли 2% или отсутствовали. Кольцо нейтрофилов аккуратно собирали, переносили в стерильную пробирку. Клетки трижды отмывали от градиента стерильным физиологическим раствором центрифугированием при 1500 об/мин 10 минут, доводили до концентрации 5х106 клеток/мл и использовали в исследованиях.
Для оценки влияния микроорганизмов на формирование НВЛ использовали различные концентрации трехкратно отмытой от питательной среды взвеси суточной культуры контрольных штаммов S. aureus («Cowan 209»), Е. coli (штамм М-17), Lactobacillus spp. (препарат "Лактобактерин сухой", фирма "ИмБио", г.Нижний Новгород), Bifidobacterium spp. (препарат "Бифидумбактерин сухой", ЗАО "Экополис", г.Ковров), С. albicans (штамм 601) и смеси этих бактерий. По стандарту мутности БАК — 10 (ООО "Ормет", г. Екатеринбург) получали концентрацию 1 млрд. микробных клеток в 1мл (1х109/мл) [Иммунологические методы…, 1981], затем разводили в 10 и в 100 раз. Таким образом были получены концентрации бактерий 1х109/мл, 1х108/мл и 1х107/мл. 1 мл взвеси нейтрофилов, выделенных на двойном градиенте фиколла-верографина и доведенных до концентрации 5х106 клеток/мл, 1 мл цельной гепаринизированной крови или 1 мл лейкоцитарной взвеси инкубировали при температуре +370С в течение 30 минут в присутствии 0,1 мл взвеси микроорганизмов доведенных до различных концентраций. Для контроля использовали 1 мл взвеси нейтрофилов, выделенных на двойном градиенте фиколла-верографина и доведенных до концентрации 5х106 клеток/мл, 1 мл цельной гепаринизированной крови или 1 мл лейкоцитарной взвеси, инкубированных в тех же условиях, но без активаторов.
Для изучения влияния гуморальных факторов сыворотку или плазму крови, полученных из гепаринизированной и негепаринизированной аутологичной крови, помещенной в термостат с температурой 370С на 30 – 60 минут [Мирахмедов У.М., Резникова Л.С., 1975], и взвесь нейтрофилов, выделенных из периферической крови на двойном градиенте фиколла-верографина, инкубировали в присутствии C. albicans (штамм 601) при 370С в течение 30 минут. Для контроля инкубировали в тех же условиях взвесь нейтрофилов без активаторов, взвесь нейтрофилов в присутствии сыворотки или плазмы и взвесь нейтрофилов в присутствии C. albicans (штамм 601).
Для инактивации естественного комплемента исследуемую сыворотку крови прогревали на водяной бане при 56оС в течение 30 минут [Мирахмедов У.М., Резникова Л.С., 1975]. Этим же методом инактивировали комплемент в плазме крови. Для сравнения использовали сухой комплемент (ФГУП «НПО „Микроген“», г.Пермь). Дозу препарата подбирали, учитывая нормальные значения активности комплемента в организме человека, в перерасчете на 1 мл клеточной взвеси. Таким образом, комплемент морской свинки тестировали в концентрациях: 400 гемолитических единиц/мл, 40 ЕД/мл (соответствует нормальному уровню активности комплемента), 4 ЕД/мл, 0,4 ЕД/мл и 0,04 ЕД/мл.
В ряде опытов для оценки влияния специфических иммуноглобулинов сыворотки и плазмы крови, участвовали лица с подтвержденным повышенным содержанием антител класса IgG к С. albicans, которое устанавливалось с помощью иммуноферментного анализа (Кандида-IgG-ИФА-БЕСТ, г.Новосибирск-117). К осадку С. albicans (штамм 601) (1х109 клеток/мл) добавляли 0,5 мл свежей сыворотки крови, инкубировали в режиме, исключающем активацию комплемента, при +4°С в течение 30 минут, встряхивая каждые 5 минут [Маянский А.Н. и др., 1982]. Затем центрифугировали при 1500 об/мин в течение 10 минут и использовали в опытах как истощенную по специфическим иммуноглобулинам сыворотку, так и опсонизированную специфическими IgG С. albicans (штамм 601). Плазму крови истощали по специфическим иммуноглобулинам так же.
Для оценки влияния различных иммуностимуляторов на формирование НВЛ in vitro, нейтрофилы, выделенные из периферической крови на двойном градиенте фиколла-верографина, инкубировали 30 минут при 370С в присутствии различных концентраций тестируемых препаратов. Для контроля гранулоциты инкубировали в тех же условиях, но без активатора.
Интерферон человеческий лейкоцитарный (интерферон альфа) (ФГУП «НПО „Микроген“», г.Пермь) – препарат для местного применения на слизистые оболочки полости носа и рта, поэтому было невозможно точно произвести перерасчет средней терапевтической дозы на 1 мл клеточной взвеси. Исходя из этого, препарат тестировался в широком диапазоне концентраций: 500 МЕ на 1 мл взвеси нейтрофилов, 50 МЕ/мл, 5 МЕ/мл, 0,5 МЕ/мл, 0,05 МЕ/мл, 0,005 МЕ/мл и 0,0005 МЕ/мл.
Выбранные нами иммуномодуляторы – «Циклоферон» (ООО «НТФФ „ПОЛИСАН“», г.Санкт-Петербург), «Пирогенал» (НИИЭМ им. Н.Ф.Гамалеи / Медгамал, Россия) и «Солкотриховак» (SOLCO BASEL AG, Швейцария), – имеют точную, описанную в инструкции по применению, дозировку для парентерального применения, поэтому тестируемые концентрации подбирались в соответствии со средней терапевтической дозой для конкретного препарата, в перерасчете на 1 мл взвеси нейтрофилов, выделенных на двойном градиенте фиколла-верографина.
Для циклоферона была определена концентрация, соответствующая средней терапевтической дозе, – 0,5 мкг/мл, в 10 раз меньше – 0,05 мкг/мл, а также в 10 и в 100 раз больше – 5 мкг/мл и 50 мкг/мл, соответственно.
Вакцину «Солкотриховак» использовали в концентрациях 0,002 мкл/мл, 0,02 мкл/мл (эквивалент средней терапевтической дозы), 0,2 мкл/мл и 2 мкл/мл.
Действие пирогенала исследовали в концентрациях: 0,002 мкг/мл, 0,02 мкг/мл (соответствует разовой терапевтической дозе), 0,2 мкг/мл и 2 мкг/мл.
Клеточную взвесь после инкубации, в зависимости от активирующего агента, исследовали различными способами.
Нейтрофилы, выделенные из периферической крови на двойном градиенте фиколла-верографина, активированные микроорганизмами или иммуномодуляторами (интерферон-, пирогенал и солкотриховак), наносили на обезжиренное предметное стекло, высушивали и фиксировали 96% этиловым спиртом и окрашивали 0,04% раствором акридинового оранжевого. Учет проводили с помощью люминесцентного микроскопа, используя фильтры, обеспечивающие возбуждающий свет с длиной волны не более 490 нм и эмиссию с длинной волны 520 нм. При этом способе окраски мы оценивали процентное содержание нейтрофилов с сегментированными ядрами, с недифференцированными ядрами и нейтрофильных внеклеточных ловушек; активность фагоцитоза, фагоцитарное число; эффективность нейтрофильных внеклеточных ловушек и индекс нейтрофильной внеклеточной ловушки [Долгушин И.И., Андреева Ю.С., патент РФ на изобретение № 2384844].
Нейтрофилы, активированные препаратом «Циклоферон», также наносили на предметное стекло, высушивали, фиксировали 96% этиловым спиртом и окрашивали 0,002% раствором Sytox green. Учет проводили также, как при окраске акридиновым оранжевым.
Препараты лейкоцитарной взвеси и нейтрофилов, выделенных из периферической крови на двойном градиенте фиколла-верографина, инкубированных в присутствии сыворотки или плазмы крови, оценивали в нативном виде при окраске акридиновым оранжевым. Учет проводили в люминесцентном микроскопе с процентным количеством нейтрофилов с сегментированным и недифференцированным ядром и нейтрофильных внеклеточных ловушек.
Из цельной крови готовили мазки, высушивали, фиксировали 96% этиловым спиртом и окрашивали по методу Романовского-Гимзе. Учет проводили при иммерсионной световой микроскопии х100х10х2, оценивали процентное содержание нейтрофилов с сегментированным ядром, с недифференцированным ядром и нейтрофильных внеклеточных ловушек.
Полученные результаты исследований были подвергнуты статистической обработке на ПК под управлением операционной системы Windows ХР с использованием пакета статистических программ «Statistica for Windows 6.0» с вычислением средней арифметической и ее стандартной ошибки (М ± m), n – количество наблюдений в выборке. Различия между сравниваемыми группами считали достоверными при р 0,05, по непараметрическим критериям Mann-Whitney, Wald-Wolfowitz [Гублер Е.В., 1973; Гланц С., 1999; Боровиков В.П., 2003]. При проведении множественных сравнений использовали поправку Бонферрони [Гланц С., 1999]. Представленные цифровые данные были округлены до второго десятичного знака.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
Для количественной оценки уровня внеклеточных ловушек образованных нейтрофилами, выделенными из периферической крови на двойном градиенте фиколла-верографина, нами были применены несколько методов индикации НВЛ при окраске фиксированных препаратов: раствором акридинового оранжевого, раствором Sytox Green и по Романовскому-Гимзе, – и нативные препараты, окрашенные раствором акридинового оранжевого.
При этих методах проводили учет процентного содержания нейтрофилов с сегментированным ядром, с недифференцированным ядром и нейтрофильных внеклеточных ловушек.
После отработки всех методик, был произведен сравнительный анализ уровня всех морфологических форм нейтрофилов, используя выбранные нами способы окраски и микроскопии. Для этого нейтрофилы, выделенные из периферической крови на двойном градиенте фиколла-верографина, после инкубации при 370С в течение 30 минут в присутствии физиологического раствора, были разделены на 4 группы. Первая и вторая группы проб наносились на предметное стекло, высушивались, фиксировались 96% этиловым спиртом, окрашивались 0,04% раствором акридинового оранжевого или 0,002% раствором Sytox Green и затем исследовалась при люминесцентной микроскопии. Нейтрофилы третьей группы также наносились на стекло, высушивались, фиксировались 96% этиловым спиртом, а затем окрашивались по методу Романовского-Гимза, учет проводили в световом иммерсионном микроскопе. Четвертую группу клеток окрашивали в нативном состоянии 0,04% раствором акридинового оранжевого, готовили препараты «раздавленная капля» и оценивали при люминесцентной микроскопии.
Уровень нейтрофилов с сегментированным ядром, с недифференцированным ядром и нейтрофильных внеклеточных ловушек достоверно не отличался ни при одном методе индикации.
С целью оценки влияния представителей микробной флоры на формирование нейтрофильных внеклеточных ловушек нами были выбраны непатогенные представители флоры человека (Lactobacillus spp. и Bifidumbacterium spp.), а также условно-патогенные представители (E. coli (штамм М-17), S. aureus(«Cowan 209»)) и грибы рода Candida, обычно обитающих на слизистых оболочках гениталий и кишечника человека. Действие выбранных микроорганизмов оценивали при контакте с клетками цельной гепаринизированной крови, лейкоцитарной взвеси, а также оценивали влияние микроорганизмов на взвесь нейтрофилов, выделенных из периферической крови на двойном градиенте фиколла-верографина.
Первоначально, нами были проведены исследования влияния различных концентраций микроорганизмов на формирование внеклеточных ловушек нейтрофилами, выделенными из периферической крови на двойном градиенте фиколла-верографина. Из условно-патогенных бактерий была выбрана E. coli (штамм М-17), а из нормальной микрофлоры – Lactobacillus spp.
Таблица 1
Влияние различных концентраций E. coli и Lactobacillus spp. на формирование внеклеточных ловушек нейтрофилами, выделенными из периферической крови на двойном градиенте фиколла-верографина (n = 12, M ± m)
| Концен-трация м/о | № группы | Морфологические формы нейтрофилов (%) | ||
| Нейтрофилы с сегментированным ядром | Нейтрофилы с недифференциро- ванным ядром | Нейтрофильные внеклеточные ловушки | ||
| Контроль | 1 | 73,63 ± 0,93 | 20,01 ± 0,89 | 6,42 ± 0,68 |
| E. coli, 1х107/мл | 2 | 69,57 ± 2,79 | 18,78 ± 2,06 | 11,61 ± 1,17 p1-2=0,016 |
| E. coli, 1х108/мл | 3 | 67,64 ± 1,72 p1-3=0,022 | 17,59 ± 1,33 | 14,78 ± 1,20 p1-3=0,009 |
| E. coli, 1х109/мл | 4 | 68,04 ± 1,79 p1-4=0,022 | 17,23 ± 2,42 | 14,82 ± 1,02 p1-4=0,009 |
| Lactobacillus spp., 1х107/мл | 5 | 70,18 ± 2,87 | 16,02 ± 1,90 | 13,81 ± 1,56 p1-5=0,009 |
| Lactobacillus spp., 1х108/мл | 6 | 62,80 ± 2,24 p1-6=0,009 | 20,77 ± 1,74 | 16,43 ± 1,72 p1-6=0,009 |
| Lactobacillus spp., 1х109/мл | 7 | 65,19 ± 2,06 p1-7=0,022 | 20,81 ± 1,36 | 14,03 ± 0,89 p1-7=0,009 |
Примечание: р – достоверность различий показателей сравниваемых групп
Как видно из таблицы 1, E. coli (штамм М-17) и Lactobacillus spp. стимулируют секрецию нейтрофильной ДНК во внеклеточное пространство, однако значимых отличий между различными концентрациями этих микроорганизмов выявлено не было. Учитывая полученные данные, было решено проводить дальнейшие исследования с концентрацией микроорганизмов 1х108/мл, что не противоречит общепринятым принципам стимуляции нейтрофилов in vitro [Иммунологические методы…, 1981].
После определения действующей концентрации нами было изучено влияние всех выбранных представителей микрофлоры на формирование нейтрофильных внеклеточных ловушек.
В результате проведенных исследований определили, что представители условно-патогенной и сапрофитной флоры (рисунок 1) оказывают стимулирующее влияние на образование внеклеточных ловушек нейтрофилами, выделенными из периферической крови на двойном градиенте фиколла-верографина, при этом наибольшее количество НВЛ обнаружено в группе нейтрофилов, активированных Candida albicans (штамм 601).

Примечание: * - уровень НВЛ достоверно выше, чем в контроле
** – уровень НВЛ достоверно выше, чем при активации другими микроорганизмами
Рисунок 1. Сравнение влияния различных представителей микробной флоры на формирование нейтрофильных внеклеточных ловушек
Проведенные исследования влияния представителей микробной флоры на формирование НВЛ в цельной гепаринизированной крови (рисунок 2) показали, что в образцах крови, активированных выбранными микроорганизмами, количество нейтрофилов с сегментированным ядром, с недифференцированным ядром и нейтрофильных внеклеточных ловушек достоверно не отличалось от таковых показателей контрольных образцов.
 Примечание: достоверных отличий в сравниваемых группах не выявлено
Примечание: достоверных отличий в сравниваемых группах не выявлено
Рисунок 2. Влияние микроорганизмов на формирование внеклеточных ловушек нейтрофилами цельной гепаринизированной крови
Исходя из полученных данных, возникло предположение о неадекватности используемой концентрации микроорганизмов для данного исследования. Было решено оценить воздействие Candida albicans в концентрациях 1х109/мл, 1х108/мл и 1х107/мл на нейтрофильные гранулоциты периферической крови. Как видно из таблицы 2, соотношение морфологических форм нейтрофилов цельной гепаринизированной крови достоверно не отличалось ни в одной пробе, независимо от концентрации C. albicans (штамм 601).
Таблица 2
Влияние различных концентраций Candida albicans на образование НВЛ нейтрофилами гепаринизированной крови (n = 10, M ± m)
| Концентрация м/о | Морфологические формы нейтрофилов (%) | ||
| Нейтрофилы с сегментированным ядром | Нейтрофилы с недифференцир-ванным ядром | Нейтрофильные внеклеточные ловушки | |
| Контроль | 87,96 ± 0,42 | 9,21 ± 0,90 | 4,79 ± 1,24 |
| Candida albicans, 1х107/мл | 82,44 ± 1,30 | 11,02 ± 1,49 | 6,63 ± 0,58 |
| Candida albicans, 1х108/мл | 84,83 ± 1,08 | 7,62 ± 1,22 | 7,64 ± 1,15 |
| Candida albicans, 1х109/мл | 82,78 ± 1,67 | 11,55 ± 0,78 | 5,56 ± 1,22 |
Примечание: достоверных отличий в сравниваемых группах не выявлено
Учитывая результаты этого исследования, была выдвинута гипотеза об ингибирующем влиянии эритроцитов на внеклеточную секрецию хроматина нейтрофилами. Поэтому дальнейшие исследования проводили в лейкоцитарной взвеси, используя в качестве активатора C. albicans в концентрациях 1х109/мл, 1х108/мл и 1х107/мл. (таблица 3)
Таблица 3
Оценка влияния Candida albicans (штамм 601) на формирование внеклеточных ловушек нейтрофилами лейкоцитарной взвеси (n = 10, M ± m)
| Концентрация м/о | Морфологические формы нейтрофилов (%) | ||
| Нейтрофилы с сегментированным ядром | Нейтрофилы с недифференциро-ванным ядром | Нейтрофильные внеклеточные ловушки | |
| Контроль | 78,42 ± 1,19 | 17,29 ± 1,27 | 4,54 ± 0,50 |
| Candida albicans, 1х107/мл | 78,93 ± 2,22 | 14,12 ± 1,98 | 7,01 ± 1,13 |
| Candida albicans, 1х108/мл | 78,98 ± 1,30 | 14,83 ± 1,30 | 6,24 ± 0,68 |
| Candida albicans, 1х109/мл | 75,61 ± 1,07 | 17,92 ± 1,24 | 6,47 ± 1,06 |
Примечание: достоверных отличий в сравниваемых группах не выявлено
Таким образом, представители условно-патогенной и нормофлоры по-разному влияют на формирование внеклеточных ловушек нейтрофилами цельной гепаринизированной крови, лейкоцитарной взвеси и нейтрофилами, выделенными на двойном градиенте фиколла-верографина. Можно предположить, что имеются гуморальные факторы, блокирующие образование нейтрофильных внеклеточных ловушек.
Нами было изучено влияние сыворотки, плазмы и содержащихся в них специфических иммуноглобулинов IgG и комплемента.
Как видно из рисунка 3, нейтрофилы, выделенные из периферической крови на двойном градиенте фиколла-верографина, после добавления к ним гомологичной сыворотки (плазмы) утрачивают способность образовывать внеклеточные ловушки в ответ на стимуляцию микробным агентом. Специфические иммуноглобулины IgG к C. albicans, содержащиеся в аутологичной сыворотке или плазме крови, не влияют на способность грибов стимулировать образование экстрацеллюлярных сетей нейтрофилами, выделенными из периферической крови на двойном градиенте фиколла-верографина. В то же время, опсонизация C. albicans специфическими IgG стимулирует фагоцитарную функцию нейтрофилов по отношению к этим микроорганизмам (таблица 4).
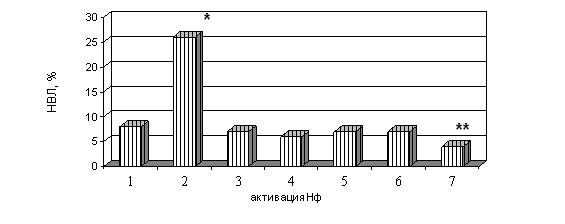 1 – Нф + физиологический раствор,
1 – Нф + физиологический раствор,
2 – Нф + C. albicans,
3 – Нф + плазма крови,
4 – Нф + плазма крови + C. albicans,
5 – Нф + опсонизированная C. albicans,
6 – Нф + истощенная по специфическим IgG плазма крови + C. albicans,
7 – Нф + прогретая при 560С плазма крови + C. albicans
Примечание: * - уровень НВЛ достоверно выше, чем в других сравниваемых группах
** – уровень НВЛ достоверно ниже, чем в других сравниваемых группах
Рисунок 3. Влияние специфических иммуноглобулинов IgG к C. albicans и аутологичного комплемента на формирование нейтрофильных внеклеточных ловушек
Таблица 4
Сравнительная характеристика фагоцитарной активности нейтрофилов, выделенных из периферической крови на двойном градиенте фиколла-верографина, по отношению к опсонизированной C. albicans и контрольному образцу C. albicans (штамм 601) (n = 12, M ± m)
| Показатели фагоцитоза | Вид микроорганизма | |
| Контрольный образец C. albicans | Опсонизированная C. albicans | |
| Активность фагоцитоза, % | 46,25 ± 0,96 | 75,00 ± 2,78 р = 0,000074 |
| Интенсивность фагоцитоза, у.е. | 0,97 ± 0,04 | 4,10 ± 0,20 р = 0,000076 |
| Фагоцитарное число, у.е. | 2,12 ± 0,07 | 5,47 ± 0,17 р = 0,000072 |
Примечание: р – достоверность различий показателей сравниваемых групп
Добавление к взвеси нейтрофилов прогретой при 560С в течение 30 минут (с целью инактивации комплемента) аутологичной сыворотки или плазмы крови несколько снижает способность клеток к образованию индуцированных C. albicans внеклеточных ловушек (рисунок 3). Добавление к взвеси нейтрофилов сыворотки крови морской свинки (в качестве источника комплемента) оказывает дозозависимое стимулирующее действие на секрецию ДНК. Такая стимуляция снижается после термической денатурации комплемента (таблица 5).
Таблица 5
Оценка влияния гетерогенного комплемента на формирование НВЛ (n = 10, M ± m)
| Активация нейтрофилов | № группы | Морфологические формы нейтрофилов (%) | |||
| Нейтрофилы с сегментирован-ным ядром | Нейтрофилы с недифференци-рованным ядром | Нейтрофильные внеклеточные ловушки | |||
| контроль | Нф + физ.раствор | 1 | 66,51 ± 1,10 | 28,48 ± 0,99 | 6,92 ± 0,55 |
| Нф + комплемент 400 МЕ/мл | 2 | 6,07 ± 0,85 р1-2 = 0,00016 | 19,71 ± 1,71 р1-2 = 0,0022 | 74,19 ± 1,90 р1-2 = 0,00016 | |
| Нф + прогретый при 560С комплемент | 3 | 42,38 ± 1,14 р1-3 = 0,00016 р2-3 = 0,00016 | 28,04 ± 1,51 р2-3 = 0,004 | 29,62 ± 1,63 р1-3 = 0,00016 р2-3 = 0,00016 | |
Примечания: р – достоверность различий показателей сравниваемых групп
Последней задачей нашего исследования была оценка влияния иммуностимуляторов различной природы на формирование внеклеточных ловушек нейтрофилами, выделенными из периферической крови на двойном градиенте фиколла-верографина. Нами были выбраны «Пирогенал» и «Солкотриховак» – иммуномодуляторы микробного происхождения, «Интерферон лейкоцитарный » – цитокин и «Циклоферон» – препарат растительного происхождения, индуктор синтеза интерферона.
«Пирогенал», «Солкотриховак» и «Циклоферон» тестировались в нескольких концентрациях с учетом средней терапевтической дозы (рисунок 4).
Как видно из рисунка 4, отмечалась выраженная зависимость биологического эффекта препаратов от его используемой дозы. Под влиянием «Пирогенала», «Солкотриховака» и «Циклоферона» в концентрации, рассчитанной для каждого конкретного препарата эквивалентно разовой терапевтической дозе, препараты максимально стимулировали формирование НВЛ нейтрофилами, выделенными из периферической крови на двойном градиенте фиколла-верографина.
 Примечание: * - уровень НВЛ достоверно выше, чем в других сравниваемых группах
Примечание: * - уровень НВЛ достоверно выше, чем в других сравниваемых группах
Рисунок 4. Влияние различных концентраций иммуностимуляторов на формирование НВЛ
«Интерферон лейкоцитарный » исследовался в широком диапазоне концентраций, так как в лечебных целях применяется на слизистые оболочки и невозможно точно рассчитать среднюю терапевтическую дозу.
 Примечание: * - уровень НВЛ достоверно выше, чем в контроле
Примечание: * - уровень НВЛ достоверно выше, чем в контроле
** – уровень НВЛ достоверно вышениже, чем в других сравниваемых группах
*** – уровень НВЛ достоверно ниже, чем в других сравниваемых группах
Рисунок 5. Влияние различных концентраций интерферона- на образование нейтрофильных внеклеточных ловушек
При изучении влияния иммуномодулятора «Интерферон лейкоцитарный » (рисунок 5), установлено, что препарат в концентрациях 5 МЕ/мл, 0,5МЕ/мл, 0,05 МЕ/мл и 0,005 МЕ/мл стимулировал образование нейтрофильных внеклеточных ловушек, при этом достоверно снижался процент нейтрофилов с сегментированным ядром. Причем максимальные изменения происходили под действие иммуностимулятора в дозе 0,5 МЕ/мл. Таким образом, полученные данные свидетельствуют о дозозависимости влияния интерферона альфа на формирование нейтрофильных внеклеточных ловушек: чрезмерно высокие концентрации ингибировали секрецию нейтрофильной ДНК, наиболее выраженное стимулирующее действие препарата на образование НВЛ определено для концентрации 0,5 МЕ/мл.
Результаты данного исследования позволяют заключить, что одним из возможных механизмов действия иммуномодуляторов является активация такой функции нейтрофильных гранулоцитов, как формирование внеклеточных ловушек.
Полученные данные свидетельствуют о том, что образование нейтрофильных внеклеточных ловушек индуцируется микроорганизмами. При этом данный феномен наблюдается только у нейтрофилов, выделенных из периферической крови на двойном градиенте фиколла-верографина. Нейтрофильные гранулоциты цельной гепаринизированной крови, лейкоцитарной взвеси, а также нейтрофилы, выделенные из периферической крови на двойном градиенте фиколла-верографина, в присутствии аутологичных сыворотки или плазмы крови, не отвечают на стимуляцию микроорганизмами экструзией ДНК в перицеллюлярное пространство. В этом, по-видимому, заключен огромный биологический смысл – образование нейтрофильных внеклеточных ловушек у здоровых лиц не должно происходить в кровяном русле. Это подтверждают литературные данные о том, что формирование НВЛ в кровотоке не только механически нарушает кровообращение тканей и органа [Clark S.R. еt аl., 2007], но может привести к развитию различных патологических состояний [Swarup V., Rajeswari M.R., 2007; Margraf S. еt аl., 2008; Logters T. еt аl., 2009], в том числе и аутоиммунных [Zhong X.-Y. еt аl., 2007; Neeli I. еt аl., 2008].
В целом, понимание того, что НВЛ, с одной стороны, могут функционировать как эффективный антимикробный механизм, а с другой – приводить к гемодинамическим расстройствам при неполноценности противодействующих механизмов регуляции, открывает новое направление в исследовании нейтрофилов в норме и при патологии.
Таким образом, несмотря на более чем вековые исследования нейтрофилов, основные клетки врожденного иммунитета скрывают от исследователей еще множество тайн. Однако уже сейчас можно с уверенностью утверждать, что нейтрофильные внеклеточные ловушки – это не только один из способов клеточной гибели, но и важный механизм противоинфекционной защиты организма, биологическая функция которого не менее важна, чем фагоцитоз и секреция биологически активных веществ (рисунок 6).

Рисунок 6. Схема основных биологических возможностей нейтрофилов
ВЫВОДЫ
- Нейтрофилы, выделенные из периферической крови человека на двойном градиенте фиколла-верографина и активированные in vitro взвесью микроорганизмов (Bifidobacterium bifidum, Lactobacillus spp., Escherichia coli, Staphylococcus aureus, Candida albicans), секретируют во внеклеточное пространство ДНК. При этом самым сильным индуктором образования ловушек из изученных микроорганизмов является Candida albicans.
- Нейтрофилы, выделенные из периферической крови на двойном градиенте фиколла-верографина, после добавления к ним гомологичной или аутологичной сыворотки (плазмы) утрачивают способность образовывать внеклеточные ловушки в ответ на стимуляцию взвесью бактерий. Нейтрофилы цельной крови и лейкоцитарной взвеси также не отвечают усилением внеклеточной секреции ДНК после инкубации со взвесью микроорганизмов.
- Добавление к взвеси нейтрофилов прогретой при 560С в течение 30 минут (с целью инактивации комплемента) аутологичной сыворотки или плазмы крови несколько снижает способность клеток к образованию индуцированных бактериями внеклеточных ловушек. Добавление к взвеси нейтрофилов сыворотки крови морской свинки (в качестве источника комплемента) оказывает дозозависимое стимулирующее действие на секрецию ДНК. Такая стимуляция снижается после термической денатурации комплемента.
- Специфические IgG к C. albicans, содержащиеся в аутологичной сыворотке или плазме крови, не влияют на способность грибов стимулировать образование экстрацеллюлярных сетей нейтрофилами, выделенными из периферической крови на двойном градиенте фиколла-верографина. В то же время, опсонизация C. albicans специфическими IgG стимулирует фагоцитарную функцию нейтрофилов по отношению к этим микроорганизмам.
- Количество эффективных внеклеточных ловушек достоверно ниже уровня активных фагоцитов в общей популяции нейтрофилов, активированных микроорганизмами (18,41 ± 1,25 % и 23,92 ± 0,79 %, соответственно). Однако количество микроорганизмов, захваченных нейтрофильными внеклеточными ловушками, значительно выше количества микроорганизмов, поглощенных при фагоцитозе, как в перерасчете на один фагоцит или ловушку (4,15 ± 0,21 у.е. и 17,73 ± 1,22 у.е., соответственно), так и на 100 нейтрофилов (97,75 ± 3,66 у.е. и 315,58 ± 19,37 у.е., соответственно).
- Иммуномодуляторы «Интерферон», «Циклоферон», «Пирогенал» и «Солкотриховак» стимулируют секрецию хроматина во внеклеточное пространство нейтрофилами, выделенными из периферической крови на двойном градиенте фиколла-верографина. Выявлен выраженный дозозависимый эффект стимулирующего действия и концентрации препаратов.
- Проведенное исследование позволяет предположить, что одним из механизмов действия препаратов «Интерферон», «Циклоферон», «Пирогенал» и «Солкотриховак» может являться активация врожденного иммунитета, в частности, образование нейтрофилами внеклеточных ловушек.
Список работ, опубликованных по теме диссертации
- Долгушин, И.И. Нейтрофильные ловушки / И.И. Долгушин, Ю.С. Андреева, А.И. Рыжкова, Е.А. Мезенцева, Е.В. Плеханова, А.Ю. Савочкина, М.А. Свиридов, Д.Н. Матвеева, А.Н. Скороходов // Российский иммунологический журнал. – 2008. – Т.2(11), №2-3. –С. 127.
- Савочкина, А.Ю. Иммунологические показатели в диагностике воспалительных заболеваний репродуктивного тракта женщин / А.Ю. Савочкина, Ю.С. Андреева, И.И. Долгушин, Л.Ф. Телешева, Е.А. Мезенцева, К.В. Никушкина, О.С. Абрамовских, Е.В. Плеханова, М.А. Свиридов, С.И. Марачев, А.И. Рыжкова // Вестник новых медицинских технологий. – 2009. – № 1. – С. 77 – 79.
- Андреева, Ю.С. Грибы рода Candida стимулируют образование нейтрофильных внеклеточных ловушек / Ю.С. Андреева, И.И. Долгушин, А.Ю. Савочкина, А.И. Рыжкова // Медицинская иммунология. – 2009. – Т. 11, № 4-5. – С. 301.
- Андреева, Ю.С. Нейтрофильные внеклеточные ловушки образуются in vitro под действием циклоферона / Ю.С. Андреева, А.Ю. Савочкина, И.И. Долгушин, А.И. Рыжкова // Вестник Уральской академической науки. – 2009. –№ 2/1. – С. 14 – 15.
- Долгушин, И.И. Влияние вакцины Солкотриховак на образование нейтрофильных внеклеточных ловушек / И.И. Долгушин, Ю.С. Андреева, А.И. Рыжкова, А.Ю. Савочкина, Е.А. Мезенцева, К.В. Никушкина, О.С. Абрамовских, С.И. Марачев, Д.Н. Матвеева // Журнал микробиологии, эпидемиологии и иммунологии. – 2009. – № 4. – С. 53 – 55.
- Долгушин, И.И. Нейтрофильные внеклеточные ловушки / И.И. Долгушин, Ю.С. Андреева, А.И. Рыжкова // Вестник новых медицинских технологий. – 2009. – Т. XVI, № 2. – С. 14 – 16.
- Мезенцева, Е.А. Влияние непатогенных и условно-патогенных представителей нормофлоры слизистых оболочек на секрецию нейтрофилами антимикробных продуктов и цитокинов / Е.А. Мезенцева, Ю.С. Андреева, И.И. Долгушин, А.Ю. Савочкина, К.В. Никушкина, О.С. Абрамовских, Е.В. Плеханова, М.А. Свиридов, С.И. Марачев, А.И. Рыжкова // Вестник новых медицинских технологий. – 2009. – Т. XVI, № 2. – С. 16 – 17.
- Рыжкова, А.И. Влияние различных концентраций препарата «Солкотриховак» на образование нейтрофильных внеклеточных ловушек / А.И. Рыжкова, И.И. Долгушин, Ю.С. Шишкова, А.Ю. Савочкина // Материалы VII итоговой науч.-практ. конф. молодых ученых ЧелГМА. – Челябинск, 2009. – С. 115 – 117.
- Рыжкова, А.И. Образование нейтрофильных внеклеточных ловушек в чистой фракции нейтрофилов и в цельной крови под влиянием грибов рода Candida / А.И. Рыжкова, Ю.С. Шишкова, А.Ю. Савочкина // Материалы VII итоговой науч.-практ. конф. молодых ученых ЧелГМА. - Челябинск, 2009. – С. 117 – 120.
- Долгушин, И.И. Нейтрофильные ловушки: методы определения, биологическая роль / И.И. Долгушин, Ю.С.Андреева, А.Ю. Савочкина, А.И. Рыжкова, В.А. Маркова, Н.А. Васильева // Медицинская наука и образование Урала. – 2009. – № 3. – С. 10 – 12.
- Шишкова, Ю.С. Влияние препарата «Пирогенал» на образование нейтрофильных внеклеточных ловушек / Ю.С. Шишкова, А.Ю. Савочкина, А.И. Рыжкова, Е.А. Мезенцева // Медицинская наука и образование Урала. – 2009. – № 3. – С. 23 – 25.
- Долгушин, И.И. Экспресс метод обнаружения нейтрофильных внеклеточных ловушек / И.И. Долгушин, Ю.С. Шишкова, А.И. Рыжкова, А.Ю. Савочкина // Труды научной сессии, посвященной 65-летию медицинской академии. – Челябинск, 2009. – С. 13 – 14.
- Шишкова, Ю.С. Препарат «Пирогенал» стимулирует образование нейтрофильных внеклеточных ловушек / Ю.С. Шишкова, А.И. Рыжкова, А.Ю. Савочкина // Труды научной сессии, посвященной 65-летию медицинской академии. – Челябинск, 2009. – С. 57 – 58.
- Долгушин, И.И. Технологии определения и роль нейтрофильных внеклеточных ловушек в антимикробной защите / И.И. Долгушин, Ю.С. Шишкова, А.Ю. Савочкина, А.И. Рыжкова, И.В. Курносенко, В.П. Евтушенко // Вестник РАМН. – 2010. – № 4. – С. 26 – 30.
На правах рукописи
Рыжкова
Анна Ивановна
влияние микробных и немикробных факторов на формирование внеклеточных ловушек нейтрофилами периферической крови
14.03.09 – клиническая иммунология, аллергология
Автореферат
диссертации на соискание ученой степени
кандидата медицинских наук
Челябинск – 2010
Отпечатано ………