

Метамерные особенности развития мезофилла и содержания пигментов пластид листьев пшеницы
На правах рукописи
Даштоян Юлия Васильевна
МЕТАМЕРНЫЕ ОСОБЕННОСТИ РАЗВИТИЯ МЕЗОФИЛЛА
И СОДЕРЖАНИЯ ПИГМЕНТОВ ПЛАСТИД ЛИСТЬЕВ ПШЕНИЦЫ
03.00.05 – ботаника
Автореферат диссертации на соискание ученой степени
кандидата биологических наук
Саратов — 2009
Работа выполнена в государственном образовательном
учреждении высшего профессионального образования
«Саратовский государственный университет им. Н. Г. Чернышевского»
на кафедре микробиологии и физиологии растений
| Научный руководитель: | доктор биологических наук, профессор Степанов Сергей Александрович |
| Официальные оппоненты: | доктор биологических наук, профессор Березуцкий Михаил Александрович |
| кандидат биологических наук, доцент Жанабекова Елена Игоревна | |
| Ведущая организация: | ФГОУ ВПО «Саратовский государственный аграрный университет им. Н.И. Вавилова» |
Защита состоится «24» декабря в 14.00 часов на заседании диссертационного совета Д 212.243.13 при Государственном образовательном учреждении высшего профессионального образования «Саратовский государственный университет им. Н.Г. Чернышевского» по адресу: 410012, г. Саратов, ул. Астраханская, д. 83, e-mail: [email protected]
С диссертацией можно ознакомиться в Зональной научной библиотеке ГОУ ВПО «Саратовский государственный университет им. Н. Г. Чернышевского».
Автореферат разослан «20» ноября 2009 г.

Учёный секретарь
диссертационного совета С.А. Невский
Общая характеристика работы
Актуальность темы. Ведущую роль среди возделываемых человеком культур играет пшеница. Создание сортов интенсивного типа с высоким потенциалом урожайности требует углубленного изучения всех элементов фотосинтетической деятельности на разных уровнях организации ассимиляционного аппарата – от ценоза до клеток и хлоропластов (Ничипорович, 1982; Robertson et al., 1995; Sims, Gamon, 2002).
Гармоничная связь фотосинтеза с другими физиологическими функциями растения, прежде всего с функциями роста и развития органов растения, требует формулирования общей теории продуктивности растений (Мокроносов, 1981; Полевой, 2001; Thomas, 1997; Zhao et al., 2003). При создании подобной теории считается необходимым обратить внимание на метамерный принцип организации структуры побега растения (Шафранова, 1989, 1990; Степанов, 2008).
Основные направления изменения фотосинтетического аппарата пшеницы при повышении плоидности и переходе от диких форм к культурным описаны (Austin et al., 1982; Pуке, Leech, 1987). В некоторых исследованиях выявлена положительная корреляция между количеством зеленых пигментов и фотохимической активностью хлоропластов листьев (Алиев, 1988; Gitelson et al., 2003).
Наименее изученным в настоящее время остается клеточный и тканевый уровни организации фотосинтетического аппарата пшеницы (Мокроносов, 1978; Thompson, 2005). Разветвленная форма отдельных клеток мезофилла была описана сравнительно недавно (Tuan, 1962; Chonan, 1965; Sasahara, 1982). На основании числа ячеек мезофилльной клетки они были подразделены на несколько типов (Березина, Корчагин, 1987). Отмечено (Thompson, 2008) наличие взаимосвязей между типом клеток, пластической деформацией их клеточных стенок и непосредственно функцией клетки.
Детальное изучение организации фотосинтетического аппарата необходимо для дальнейшей успешной работы по созданию общей теории продуктивности растений, внедрения полученных результатов в селекционный процесс.
Цель и задачи исследования. Целью настоящей работы является установление метамерных особенностей развития мезофилла и содержания пигментов пластид листьев мягкой яровой пшеницы. В ходе реализации цели решались следующие задачи:
1. Выявить типологию клеток мезофилла листа мягкой пшеницы.
2. Установить метамерные особенности организации мезофилла пластинки листьев мягкой пшеницы.
3. Определить сортовые особенности организации мезофилла пластинки листьев мягкой пшеницы.
4. Представить характеристику роста и развития мезофилла пластинки листа мягкой пшеницы.
5. Установить метамерные и сортовые особенности в содержании пигментов пластид пластинки листьев мягкой пшеницы.
Научная новизна. В пластинке листа выявлено большое разнообразие морфологии клеток мезофилла, которые впервые были разделены на 11 типов. Отмечено, что мезофилл пластинки 1–3-го листьев представлен всеми типами клеток. В пластинке 4–7-го листьев разнообразие типов клеток мезофилла уменьшается, существенно увеличивается доля клеток с выраженной ячеистой формой. Выявлено, что линейные параметры клеток мезофилла пластинки листьев мягкой яровой пшеницы отражают свойства генотипа сорта. Выявлено, что линейные параметры клеток мезофилла пластинки листьев мягкой яровой пшеницы отражают свойства генотипа сорта. Длина клеток у всех сортов, в листьях верхних (5 – 7-го) метамеров, как правило, больше. Минимальные значения ширины клеток мезофилла наблюдаются в пластинке верхнего, 7-го листа. Размер разных типов клеток мезофилла не обязательно положительно коррелирует с формой клетки или с числом ячеек в ней. Отмечено, что определение среди дифференцирующейся паренхимы пластинки листа клеток мезофилла становится возможным спустя 3,5 недели с момента роста листа, по достижении им длины 20–30 мм. С момента дифференциации клеток мезофилла происходит возрастание разнообразия типов клеток мезофилла, некоторые из которых в дальнейшем преобладают. Установлено, что в содержании хлорофиллов а и b, каротиноидов, соотношения между ними в пластинках листьев наблюдаются особенности, отражающие их положение в системе метамеров побега, а также агроклиматические условия. В пределах одноименных листьев могут наблюдаться сортовые различия по содержанию хлорофилла а, b и каротиноидов, величине соотношения между ними.
Теоретическая и практическая значимость. Установлены качественные и количественные характеристики различных типов клеток мезофилла пластинки листа мягкой яровой пшеницы. Представлены особенности дифференциации мезофилла пластинки в процессе роста и развития листа. Выявлены метамерные и сортовые особенности в качественном и количественном содержании пигментов пластид. Полученные в результате исследования сведения вносят вклад в изучение анатомии, морфологии и физиологии одной из важнейших сельскохозяйственных культур – пшеницы и могут быть использованы для теоретического описания морфогенеза растений, в селекции для оценки продуктивности сортов. Результаты исследований использованы при выполнении НИР «Структурные и функциональные аспекты межметамерных взаимосвязей в онтогенезе побега пшеницы» в Саратовском государственном университете. Материалы исследования внедрены в учебный процесс, используются на лекциях и практических занятиях по анатомии и физиологии растений, а также при проведении лабораторных практикумов, выполнении курсовых и дипломных работ в Саратовском государственном университете.
Апробация работы. Основные результаты исследований представлены на: Международной конференции по морфологии растений, посвященной памяти И.Г. и Т.И. Серебряковых (Москва, 2004); Всероссийской научно-практической конференции, посвященной 117-й годовщине со дня рождения Н.И. Вавилова «Вавиловские чтения – 2004» (Саратов, 2004); Научной конференции аспирантов и студентов биологического факультета СГУ (Саратов, 2005); 4-й Всероссийской конференции молодых ученых «Наука. Образование. Молодежь» (Майкоп, 2007); Международной конференции «Современная физиология растений: от молекул до экосистем» (Сыктывкар, 2007).
Публикации. По материалам диссертации опубликовано 10 работ, одна из которых в издании, рекомендованном Перечнем ВАК РФ.
Декларация личного участия автора. Автор лично провел полевые и лабораторные эксперименты, осуществил сбор объектов, провел необходимые исследования. Обработка полученных данных, их интерпретация проведены автором самостоятельно. В совместных публикациях вклад автора составил 50 – 80%.
Структура и объем диссертации. Диссертационная работа изложена на 158 страницах машинописного текста и состоит из введения, 5 глав, заключения, выводов и 2 приложений, содержит 21 таблицу и 41 рисунок. Список цитированной литературы включает 276 отечественных и иностранных источников.
Основные положения, выносимые на защиту:
1. Мезофилл пластинки листа пшеницы представлен различными типами клеток, доля представительства которых зависит от метамерной принадлежности листа и сортовых особенностей растений.
2. Дифференциация мезофилла пластинки сопровождается последовательным изменением числа отдельных типов клеток в процессе роста и развития листа.
3. Содержание пигментов фотосинтеза, соотношение между ними зависит от метамерной принадлежности листа и генотипа сорта.
Содержание работы
Во введении дается краткая характеристика объекта и предмета исследования, формулируются цель и задачи, основные положения, выносимые на защиту, определяется актуальность, новизна, теоретическая и практическая значимость работы.
Глава 1. МЕТАМЕРНЫЕ ОСОБЕННОСТИ АНАТОМИИ, РАЗВИТИЯ ЛИСТЬЕВ ПШЕНИЦЫ И СОДЕРЖАНИЯ ПИГМЕНТОВ ПЛАСТИД
(обзор литературы)
На основе анализа отечественной и зарубежной литературы в главе рассматриваются понятие о метамере побега и его образовании, вопросы анатомо-морфологической организации пластинки листьев пшеницы, особенности организации мезофилла листьев пшеницы, роста и развития листьев. Проведен анализ литературных данных по вопросам метамерной специфичности фотосинтетического аппарата пшеницы, состава и содержания пигментов пластид в мезофилле листа пшеницы.
Глава 2. ОБЪЕКТЫ ИССЛЕДОВАНИЙ И МЕТОДЫ
Исследования проводились на кафедре микробиологии и физиологии растений биологического факультета СГУ и в полевых условиях селекционного севооборота НИИСХ Юго-Востока в период с 2003 по 2008 гг.. Для проведения исследований в лабораторных условиях использовались растения, выращиваемые в полевых мелкоделяночных опытах в трехкратной повторности. Обработка полей полностью соответствовала агротехническим требованиям, предъявляемым в зоне для возделывания яровой пшеницы.
В качестве объектов изучения были выбраны следующие сорта мягкой пшеницы: Саратовская 36, Нададорес 63, Саратовская 52, Лютесценс 62, Саратовская 56, Саратовская 58.
Для определения динамики роста листьев и их частей (пластинки и влагалища) с момента посева зерновок пробы брали (15 растений в каждой из трех повторностей, которые затем объединяли и отбирали группу из 20 растений) с периодичностью 48 ч. до момента прекращения их линейного роста. Определение абсолютной и относительной скорости роста примордия, пластинки и влагалища 5-го листа побега проводилось по Вильямсу (Williams, 1975).
Для анатомических исследований пластинок листьев (n=15) часть объектов фиксировалась в слабом растворе Навашина по М.Н. Прозиной (1960). Срезы готовились по общепринятой методике и окрашивались гематоксилином Гейденгайна и альциановым синим (Дженсен, 1965; Градчанинова, 1978). Толщина срезов составляла 10–15 мкм.
Для определения формы и параметров клеток мезофилла листа использовали среднюю часть пластинки (длина – 2 см, ширина – 0,5 см) листьев 1–7 метамеров побега. Мацерацию осуществляли в течение 15–60 мин. в зависимости от изучаемого типа клеток при нагревании на водяной бане в растворе соляной (1%) и хромовой (5%) кислот – 1:1. Определение типологии клеток мезофилла и разделение их на группы осуществлялось с помощью камеры Горяева (Березина, 1988; Бурундукова, 1993). Морфометрическое определение длины и ширины клеток мезофилла осуществлялось с помощью окуляр-микрометра МБС-9, n=50. Фотографии были получены при помощи цифровой камеры-окуляра для микроскопа SCOPETEK, модель DСМ35.
Количественное определение пигментов в пластинке листьев (n=3 - 4) пшеницы проводилось на спектрофотометре HEWLET PACKARD (США). Содержание хлорофиллов а и b, каротиноидов определялось в вытяжке пигментов без предварительного их разделения на основании экспериментально полученных удельных коэффициентов поглощения для 100%-ного ацетона – по Хольму-Веттштейну (Гавриленко, Жигалова, 2003).
Результаты исследований подвергались статистической обработке по Б.А. Доспехову (1985).
Глава 3. СТРУКТУРА МЕЗОФИЛЛА ПЛАСТИНКИ
ЛИСТЬЕВ ПШЕНИЦЫ
Для клеток мезофилла пластинки листа пшеницы характерна сильная разветвлённость клеточных стенок, являющаяся необходимым условием поддержания в ходе эволюции оптимального соотношения поверхности клетки к её объёму, а также наличие значительной доли межклетников. В пластинке листа выявлено несколько типов проводящих пучков: 1) крупные, смыкающиеся с поверхностью пластинки посредством хорошо выраженных склеренхимных тяжей по обе стороны от проводящего пучка; 2) крупные, имеющие склеренхимный тяж только к одной из сторон (адаксиальной или абаксиальной) пластинки листа; 3) мелкие пучки с недостаточно развитой ксилемой; 4) поперечные проводящие пучки. Некоторые крупные проводящие пучки имели двойную обкладку из клеток, вытянутых вдоль продольной оси пластинки листа (рис. 1).

Рис. 1. Поперечный срез пластинки 5-го листа мягкой пшеницы сорта Саратовская 36.
1 – клетки мезофилла; 2 – внешняя обкладка проводящего пучка; 3 – внутренняя обкладка проводящего пучка; 4 – флоэма пучка; 5 – ксилема пучка; 6 – волокна склеренхимы. Ув. 1040.
Клетки внешней обкладки содержали хлоропласты, тогда как клетки внутренней обкладки пучка были представлены типичными волокнами склеренхимы, соединенными друг с другом плазмодесмами. Особенности роста, ланцетовидная форма пластинки листа пшеницы, разнообразие типов проводящих пучков предполагает (Березина, 1989; Terashima, Saeki, 1985; Pуке, Leech, 1987) наличие различий по форме клеток и структурированности мезофилла в разных частях пластинки.
Типология клеток мезофилла листа пшеницы. В основу типологии клеток мезофилла пластинки листьев пшеницы были включены следующие признаки: 1. Наличие симметричности клетки в целом (определяется числом осей симметрии на двумерной плоскости); 2. Наличие или отсутствие лопастей клетки ("protuberance" по Chonan, 1965; "lobes" по Parker, Ford, 1982; "arm" по Sasahara, 1982); 3. Ширина цитоплазматического мостика (Березина, 1986) или глубина «перетяжек» между ячейками; 4. Пространственное положение ячейки относительно продольной оси клетки – перпендикулярно или наклонно с различным углом между продольной и поперечной осями симметрии на двумерной плоскости; 5. Соотношение длины к ширине клетки. Каждый из признаков, включенных нами в основу типологии клеток мезофилла, в совокупности позволяет характеризовать форму клеток ассимиляционной паренхимы пластинки листа.
Было выявлено большое разнообразие морфологии клеток мезофилла пластинки листа пшеницы, которые мы разделили на 11 основных типов: A, B, C, D, E, F, G, H, I, J и K. Некоторые из них представлены на рисунке 2:
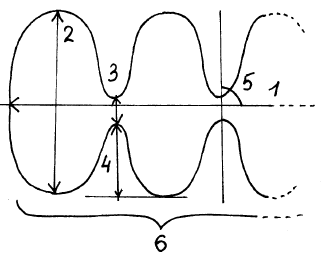 |  |  |
| A | J | K |
Рис. 2. Некоторые типы клеток мезофилла пластинки листа мягкой яровой пшеницы сорта Саратовская 36: 1 – длина клетки, 2 – ширина клетки, 3 – ширина цитоплазматического мостика, 4 – глубина перетяжки, 5 – угол наклона ячеек по отношению к продольной оси клетки, 6 – число ячеек в клетке
По принципу симметрии клетки были объединены в две группы: симметричные (к этой группе были отнесены клетки А, В, С, F, G-типов, имеющие одинаковое число ячеек с обеих сторон от продольной оси клетки) и не симметричные (клетки H, I, J, К-типов, отличающиеся разным числом ячеек по обе стороны от продольной оси клетки). Клетки D-типа представляли собой простые паренхимные клетки, не имеющие ячеистой формы, клетки Е-типа – преимущественно прозенхимные клетки с извилистыми стенками, наличие ячеек морфологически не определялось.
Для клеток A, F, H, J-типов ширина цитоплазматического мостика существенно меньше относительно других клеток, глубина перетяжки составляет 30–45% от ширины клетки, для клеток B, G, I-типов – в пределах 15–30% ширины клетки; клетки С, D и Е-типов либо не имели четкого деления на ячейки или с перетяжками, составляющими не более 15% от ширины клетки.
Клетки A, B, C, H-типов имели ячейки, расположенные перпендикулярно относительно продольной оси клетки, F, G, I, J-типов – наклонно, с различным углом между продольной и поперечной осями симметрии. Количество ячеек в некоторых типах клеток может варьировать от 2 до 14.
Типы клеток различались также по величине соотношения длины и ширины: А, В, F,G,H, I-типы – от 0,77 до 9,40; С, D, E, J и К-типы – от 0,96 до 4,00.
Основываясь на полученных результатах и литературных данных (Березина, 1989; Parker, Ford, 1982; Бурундукова и др., 1993, 2003), следует, что отмеченное явление универсально среди ведущих культурных злаков и подобная организация мезофилла может способствовать расширению адаптационного потенциала вида или сорта к агроклиматическим условиям.
Метамерные особенности организации мезофилла пластинки листьев пшеницы. Установлена специфика варьирования числа клеток мезофилла различных типов в листьях разных метамеров. В пластинке 1 и 2-го листьев встречались с разной частотой все типы клеток мезофилла, начиная с третьего листа разнообразие типов клеток мезофилла уменьшалось. В частности, в третьем листе не были обнаружены клетки E-типа, четвертом – E и К-типов, пятом – D, E и K-типов, 6-м, кроме перечисленных, – клетки J-типа, а в 7-м – также клетки C-типа. В пластинках 4–7-го листьев преобладали клетки A, В, F и H-типов, которые отличались значительным варьированием числа ячеек, соотношением длины к ширине клетки и глубокими перетяжками, определяющими ширину цитоплазматического мостика между ячейками (табл. 1).
Таблица 1
Содержание различных типов клеток мезофилла пластинки
листьев мягкой пшеницы сорта Саратовская 36
| Номер листа | Типы клеток мезофилла листа пшеницы, % | ||||||||||
| A | B | C | D | E | F | G | H | I | J | K | |
| 1 | 16 | 22 | 2 | 4 | 4 | 28 | 4 | 8 | 4 | 4 | 4 |
| 2 | 36 | 12 | 6 | 2 | 4 | 16 | 2 | 16 | 2 | 2 | 2 |
| 3 | 30 | 24 | 2 | 2 | 0 | 26 | 4 | 6 | 2 | 2 | 2 |
| 4 | 28 | 8 | 2 | 2 | 0 | 32 | 4 | 16 | 6 | 2 | 0 |
| 5 | 32 | 8 | 2 | 0 | 0 | 34 | 4 | 12 | 2 | 6 | 0 |
| 6 | 40 | 12 | 8 | 0 | 0 | 16 | 8 | 12 | 4 | 0 | 0 |
| 7 | 28 | 4 | 0 | 0 | 0 | 39 | 3 | 16 | 8 | 0 | 0 |
Линейные параметры клеток мезофилла изменялись в связи с принадлежностью листа тому или иному метамеру. Наибольшую длину имели клетки пластинки листьев верхних, 5–7-го метамеров, наименьшую ширину – клетки мезофилла пластинки 7-го листа (рис. 3, 4).
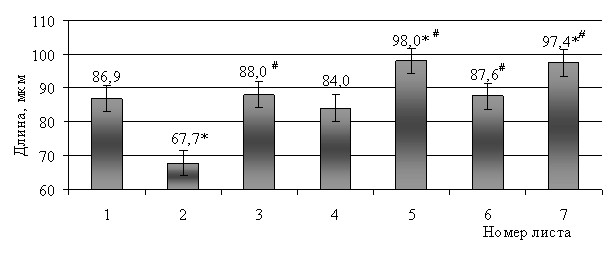
Рис. 3. Длина клеток мезофилла пластинки листьев мягкой яровой пшеницы
сорта Саратовская 36, мкм.
Примечание: * - достоверная разница относительно первого листа при Р 0,05;
# - достоверная разница относительно предыдущего листа при Р 0,05.
Выявлено, что размер клеток мезофилла не обязательно положительно коррелирует с формой клетки или с числом ячеек в ней. Клетки одного типа могут иметь различные размеры, клетки с меньшим числом ячеек – большие линейные размеры или наоборот.
Например, в пластинке 2-го листа среди клеток F-типа наблюдались клетки с 2–7 ячейками, длиной от 44,9 до 126,5 мкм и шириной от 21,6 до 53,3 мкм. В пластинке 5-го листа среди клеток F-типа отмечены клетки с 3–9 ячейками, но несколько большей максимальной длиной – от 36,6 до 163,2 мкм. Ширина различных клеток этого типа была примерно одинаковой в пластинке 2-го листа. Среди клеток F-типа в пластинке 7-го листа отмечены клетки с 3–10 ячейками, максимальная длина отдельных клеток была меньше по сравнению с клетками этого типа в пластинке 5-го листа.
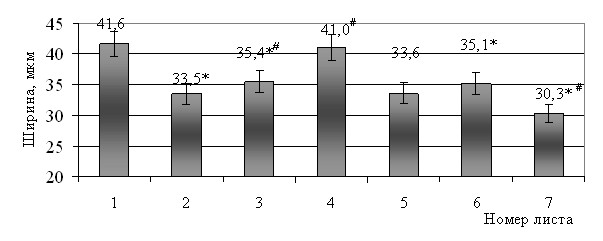
Рис. 4. Ширина клеток мезофилла пластинки листьев мягкой яровой пшеницы
сорта Саратовская 36, мкм.
Примечание: * - достоверная разница относительно первого листа при Р 0,05;
# - достоверная разница относительно предыдущего листа при Р 0,05.
Сортовые особенности организации мезофилла пластинки листьев мягкой пшеницы. Исследуя особенности организации мезофилла пластинки листьев других сортов мягкой пшеницы (Лютесценс 62, Саратовская 52, Саратовская 56, Саратовская 58, Нададорес 63) было установлено, что отмеченные ранее закономерности на примере мягкой пшеницы сорта Саратовская 36 справедливы и для указанных сортов этого вида. В частности, среди клеток мезофилла первых двух листьев присутствовали все типы клеток; в листьях выше расположенных метамеров разнообразие типов клеток мезофилла уменьшалось, в флаговом листе преобладали клетки A, В, F и H-типов и отсутствовали клетки C, D, E, J и K-типов.
С учетом доли представительства каждого из типов клеток в мезофилле пластинки листьев анализ их средней длины позволил выявить следующее. У растений сорта Лютесценс 62 минимальная длина клеток была отмечена в пластинке 2-го листа, максимальная – в пластинке 5-го листа. Ширина клеток составляла от 29,6 (7-й лист) до 38,6 мкм у 2-го листа. Для Саратовской 52 минимальная длина клеток мезофилла пластинки отмечена у 3-го листа (52,2 мкм), максимальная – у 4-го и 7-го – 97,4 и 96,6 мкм соответственно. Ширина клеток достигала 20 мкм у 3-го листа и 43,3 мкм у 4-го. У Саратовской 56 минимальная длина клеток была отмечена в пластинке 4-го листа, максимальная – в пластинке 5-го. Ширина клеток составляла от 30,1 мкм (6-й лист) до 42,4 мкм у 2-го листа. Для Саратовской 58 минимальная длина клеток была отмечена в пластинке 3-го листа, максимальная – в пластинке 6-го листа. Ширина клеток составляла от 23,3 (3-й лист) до 48,9 мкм у 4-го листа. У Нададорес 63 минимальная длина клеток была отмечена в пластинке 1-го листа, максимальная – в пластинке 6-го. Ширина клеток составляла от 15,6 мкм (1-й лист) до 39,1 мкм у 3-го листа (табл. 2).
Таблица 2
Сортовые особенности развития клеток мезофилла мягкой пшеницы
| Сорт | Параметры клетки, мкм | Номер листа | ||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | ||
| Лютесценс 62 | длина | 80,6± 3,22 | 70,3± 2,11 | 75,9± 2,28 | 81,7± 3,27 | 103,2± 5,16# | 85,2± 3,41 | - |
| ширина | 33,3± 1,00 | 38,6± 1,16 | 32,2± 0,97 | 34,6± 1,04 | 37,8± 1,13 | 29,6± 0,59 | - | |
| Саратовская 52 | длина | 71,5± 2,15 | 73,8± 2,21 | 52,2± 0,52#* | 97,4± 4,87# | 92,4± 4,62# | 85,9± 3,44# | 96,6± 4,85# |
| ширина | 36,7± 1,10 | 37,7± 1,13 | 20,0± 0,40#* | 43,3± 1,73* | 28,8± 0,58#* | 34,0± 1,02 | 35,1± 1,05 | |
| Нададорес 63 | длина | 55,0± 0,58* | 78,1± 2,34# | 101,4± 5,07#* | 91,8± 4,59# | 103,3± 5,17# | 109,7± 5,49#* | 100,3± 5,17# |
| ширина | 15,6± 0,16* | 36,2± 1,09# | 39,1± 1,17#* | 27,8± 0,56#* | 32,2± 0,97# | 36,7± 1,10#* | 35,7± 1,07# | |
| Саратовская 56 | длина | 79,5± 2,39 | 81,0± 3,24 | 78,0± 2,34 | 72,2± 2,17 | 102,0± 5,1# | 90,1± 4,51 | 81,9± 3,28 |
| ширина | 33,9± 1,01 | 42,4± 1,70# | 31,2± 0,94 | 41,8± 1,67#* | 33,3± 0,99 | 30,1± 0,90 | 33,4± 1,00 | |
| Саратовская 58 | длина | 71,4± 2,14 | 77,1± 2,31 | 45,5± 0,46#* | 87,6± 3,50# | 90,8± 4,54# | 104,7± 5,24#* | - |
| ширина | 29,6± 0,59 | 43,9± 1,76# | 23,3± 0,47#* | 48,9± 1,96#* | 31,5± 0,95* | 33,8± 1,01 | - | |
Примечание: * - достоверная разница относительно сорта Лютесценс 62 при Р 0,05;
# - достоверная разница относительно первого листа при Р 0,05
Таким образом, линейные параметры клеток мезофилла пластинки листьев мягкой яровой пшеницы отражают свойства генотипа сорта. Однако, как правило, длина клеток у всех сортов больше в листьях верхних метамеров – 5–7-го. Ширина клеток может быть меньше как в листьях верхних метамеров, так и в листьях нижних метамеров.
Глава 4. РОСТ И РАЗВИТИЕ МЕЗОФИЛЛА
ПЛАСТИНКИ ЛИСТА ПШЕНИЦЫ
Исследование проводили с использованием 5-го листа мягкой пшеницы сорта Саратовская 36. Было установлено, что рост пластинки пятого листа в длину соответствует сигмовидной кривой, отражающей непрерывный ход роста органа от его начала до прекращения (Обручева, Ковалев, 1979; Streck et al., 2003).От инициации примордия пятого метамера на 5–6-й день вегетации до достижения суммарной длины листа (пластинка + влагалище) 800–2000 мкм, характерно постепенное возрастание абсолютной скорости роста – до 300 мкм/день. Этот этап соответствует лаг-фазе сигмовидной кривой. В момент перехода с этой фазы на лог-фазу роста, что соответствует фенофазе 3-го листа, наблюдалось образование язычка, разделяющего примордий на собственно пластинку и влагалище.
Абсолютная и относительная скорость роста пластинки в длину с 24–25 дня от посева многократно увеличивается. Завершение первой фазы роста и начало второй фазы зафиксировано на 26–27-й день с начала вегетации при достижении длины листа 0,25 мм. В лог-фазу рост пластинки пятого листа идет с максимальной скоростью и к моменту завершения роста лист достигал в длину 198–210 мм на 40–42-ой день вегетации. В третью фазу роста пластинки 5-го листа отмечено незначительное увеличение длины – до 225 мм.
Одновременно с ростом пластинки листа в длину происходил её рост в поперечном направлении. Выявлено, что рост пластинки 5-го листа в поперечном направлении происходит с некоторым запозданием. В частности, если возрастание абсолютной и относительной скорости роста пластинки листа в длину отмечалось на 24–25 день, то возрастание роста пластинки в поперечном направлении наблюдалось с 30–31 дня с момента посева. Далее, с 31-го по 44-й день, наблюдался интенсивный диффузный рост пластинки листа в ширину при участии клеток межпучковой паренхимы и маргинальных меристем. В этот период ширина листовой пластинки увеличивается с 0,25 до 7,3 мм, т. е. примерно в 30 раз. Завершение роста пластинки пятого листа в ширину, что соответствует по времени третьей фазе, также наблюдалось позднее по сравнению с ростом в длину. В эту фазу ширина пластинки увеличивалась с 7,3 до 9,0 мм, т. е. возрастала в 1,2 раза.
Параллельно с ростом пластинки листа происходила дифференциация клеток, представленных в ней. Отмечено, что одними из первых дифференцируются клетки эпидермиса и проводящих пучков. Вначале роста пластинки 5-го листа в толщину в ней наблюдалось в зоне межпучковой паренхимы четыре слоя клеток; большее их число, от 6 до 9, отмечено в зоне инициации проводящих пучков (рис. 5).

Рис. 5. Поперечный срез побега мягкой пшеницы сорта Саратовская 36 на уровне основания конуса нарастания в момент инициации 6-го листа: 1 – пятый лист; 2 – образующийся шестой лист; 3 – основание конуса нарастания; 4 - эпидермис; 5 - клетки проводящего пучка;
6 – межпучковая паренхима листа. Ув.1040.
Определение среди дифференцирующихся паренхимных клеток пластинки листа специализированных клеток, в частности клеток мезофилла, становится возможно спустя примерно 3,5 недели с момента роста листа, когда он достигнет длины 20–30 мм. Для первого листа, по данным (Michalkova, 1992), его длина в момент дифференциации клеток мезофилла составляет 17 мм.
Существенное увеличение клеток мезофилла пластинки листа в длину происходило на 24–25 день вегетации, что соответствовало возрастанию абсолютной и относительной скорости роста. С 26 по 40 день, т. е. в течение 2-х недель длина клеток возрастала с 65,6 до 91,2 мкм. В дальнейшем отмечено варьирование длины клеток, что возможно (Thompson, 2005, 2008), определялось структурными изменениями самих клеток.
Рост клеток мезофилла в продольном направлении сопровождался также их ростом в поперечном направлении, который отмечался несколько позднее. В частности, если рост клеток в длину наблюдался на 24–25 день, то рост клеток в ширину через 6–7 дней. С 30 по 38 день средняя ширина клеток возрастала от 19,6 до 46,4 мкм. В дальнейшем отмечалось варьирование ширины клеток в пределах 30,7–41,5 мкм.
До 24-го дня вегетации клетки мезофилла 5-го листа пшеницы представляли собой простые паренхимные клетки. Они не имели характерной ячеистой формы и находились в состоянии интенсивного деления. С 24-го дня клетки приобретали ячеистую форму, причем в суспензии присутствовали как 2-х-ячеистые так и 5–6-ячеистые клетки, однако ячейки были выражены слабо. В дальнейшем было отмечено возрастание разнообразия типов клеток мезофилла. Этот процесс продолжается на протяжении всей лог-фазы роста пластинки. C наступлением фазы завершения роста разнообразие типов клеток уменьшалось, т. е. происходила, на наш взгляд, некая унификация структуры мезофилла (табл. 3).
Таблица 3
Изменение разнообразия типов клеток мезофилла в онтогенезе
пластинки пятого листа мягкой пшеницы сорта Саратовская 36
| Фаза роста | День вегетации | Типы мезофилльных клеток, % | ||||||||||
| A | B | C | D | E | F | G | H | I | J | K | ||
| Лог-фаза роста пластинки листа | 24-й | 0 | 0 | 26 | 18 | 36 | 0 | 20 | 0 | 0 | 0 | 0 |
| 26-й | 0 | 12 | 28 | 12 | 32 | 0 | 16 | 0 | 0 | 0 | 0 | |
| 28-й | 0 | 28 | 20 | 12 | 16 | 0 | 24 | 0 | 0 | 0 | 0 | |
| 30-й | 8 | 20 | 12 | 0 | 14 | 18 | 16 | 8 | 4 | 0 | 0 | |
| 32-й | 8 | 12 | 16 | 0 | 10 | 26 | 8 | 12 | 8 | 0 | 0 | |
| 34-й | 16 | 12 | 6 | 0 | 10 | 32 | 8 | 12 | 4 | 0 | 0 | |
| 36-й | 16 | 4 | 4 | 0 | 4 | 42 | 16 | 12 | 2 | 0 | 0 | |
| 38-й | 20 | 10 | 2 | 0 | 0 | 44 | 4 | 10 | 4 | 4 | 2 | |
| 40-й | 20 | 8 | 8 | 0 | 0 | 22 | 20 | 10 | 6 | 4 | 2 | |
| Фаза завершения роста листа | 42-й | 20 | 10 | 24 | 0 | 0 | 20 | 4 | 8 | 4 | 6 | 4 |
| 44-й | 32 | 6 | 4 | 0 | 0 | 30 | 6 | 14 | 4 | 4 | 0 | |
| 46-й | 26 | 4 | 6 | 0 | 0 | 32 | 6 | 12 | 4 | 10 | 0 | |
| 48-й | 32 | 6 | 6 | 0 | 0 | 30 | 4 | 12 | 6 | 4 | 0 | |
| 50-й | 32 | 8 | 4 | 0 | 0 | 32 | 4 | 12 | 4 | 4 | 0 | |
По достижении максимальных линейных параметров развития клеток мезофилла в дальнейшем их размеры могут изменяться. Этот процесс дополняется, как показано выше, структурной реорганизацией мезофилла на уровне отдельных типов клеток. Возможными причинами установленных фактов, очевидно, являются: 1) продолжающаяся дифференциация клетки по завершении её роста, сопровождающаяся процессами старения и деградации отдельных её компартментов (Pуке, Leech, 1987; Hoson, 1990; Kragler et al., 1998; Thompson, 2008); 2) лизис отдельных клеток (Шарова, 2004; McQueen-Mason, Cosgrove, 1995).
Глава 5. СОСТАВ И СОДЕРЖАНИЕ ПИГМЕНТОВ ФОТОСИНТЕЗА
В ПЛАСТИНКЕ ЛИСТЬЕВ ПШЕНИЦЫ
Количество пигментов фотосинтеза в листьях пшеницы, являясь наследуемым сортовым признаком, зависит от их возрастного состояния и фазы развития растения (Кумаков, 1985). На примере сорта яровой мягкой пшеницы Саратовской 58 выявлено, что в пластинке 1-го листа в условиях вегетации 2004 г. относительное содержание пигментов составляло: хлорофилла а – 57%, хлорофилла b – 26,1%, каротиноидов – 16,9%. Абсолютное содержание пигментов фотосинтеза имело следующие значения: хлорофилла а – 1,32, хлорофилла b – 0,61, каротиноидов – 0,39 мг/г сырого веса. Отношение хлорофилла а к хлорофиллу b у растений этого сорта было равно 2,16, хлорофиллов к каротиноидам – 4,95. В условиях вегетации 2005 г. абсолютное содержание хлорофилла а составляло 1,57мг/г сырого веса, хлорофилла b – 0,78 мг/г, каротиноидов – 0,46 мг/г. Отношение хлорофилла а к хлорофиллу b было равно 2,0, а хлорофиллов к каротиноидам – 5,06. Иное по сравнению с 2004 г. содержание пигментов фотосинтеза – хлорофиллов и каротиноидов, возможно, определялось агроклиматическими условиями. Однако сотношение количества пигментов другу к другу сохранилось примерно на том же уровне значений.
Состав и содержание пигментов фотосинтеза в пластинке листьев разных метамеров побега мягкой пшеницы сорта Саратовская 58. При исследовании метамерной изменчивости состава и содержания пигментов фотосинтеза в листовых пластинках мягкой пшеницы сорта Саратовская 58 было установлено, что существуют определенные особенности в содержании различных пигментов как и соотношения между ними в листьях пшеницы, принадлежащих разным метамерам побега. В частности, в 2004 г. доля хлорофилла а варьировала от 53,7 (второй лист) до 61,7% (третий лист). В 2005 г. относительное содержание хлорофилла а составляло от 54,5 (третий лист) до 58,4% (второй лист) побега пшеницы. Относительное содержание хлорофилла b в пластинке листьев также различалась по годам вегетации. Содержание хлорофилла b составляло: в условиях 2004 г. от 14,1 (третий лист) до 26,1% (первый лист), в 2005 г. – от 19,9 (седьмой лист) до 28,1 % (третий лист).
Отмечено, что относительное содержание хлорофиллов а и b в пластинке каждого листа имеет противоположную тенденцию, как правило, большему содержанию хлорофилла а соответствует меньшее содержание хлорофилла b и наоборот.
Относительное содержание каротиноидов в пластинках листьев пшеницы изменялось в зависимости от принадлежности листа тому или иному метамеру побега и условий вегетации. Содержание каротиноидов составляло: в 2004 г. от 16,9% в первом листе до 24,2% в третьем, в 2005 г. от 16,3% в первом листе до 23,3% в пятом. Минимальное относительное содержание каротиноидов по результатам двух лет исследования наблюдалось в первом листе (рис. 6).
Рис. 6. Относительное содержание каротиноидов в пластинке листьев
мягкой яровой пшеницы сорта Саратовская 58, %.
На основании проведенных исследований можно заключить, что существуют определенные особенности в содержании различных пигментов как и соотношения между ними в листьях пшеницы, принадлежащих разным метамерам побега. Отмеченные особенности могут свидетельствовать о наличии свойств автономности каждого из метамеров, выступающих в качестве самостоятельных донорно-акцепторных единиц в онтогенезе растений (Мокроносов,1993; Степанов, 2008).
Сортовые особенности в содержании пигментов фотосинтеза пластинки листьев пшеницы. Содержание хлорофилла а в пластинке листьев исследуемых сортов составляет от 0,53 до 2,68 мг/г. Минимальное содержание хлорофилла а отмечалось у большинства сортов в 1–5-ом листьях. Максимальное содержание хлорофилла а свойственно как правило верхним листьям. На содержание хлорофилла а существенное влияние оказывают условия вегетации.
Содержание хлорофилла b в пластинке листьев составляет от 0,16 до 1,28 мг/г. Минимальное содержание хлорофилла b у большинства сортов выявлено в 3 или 5-ом листьях. Максимальное содержание хлорофилла b у большинства сортов наблюдалось в расположенных ниже листьях.
Величина соотношения между хлорофиллами а к b составляет 1,19–5,53. Минимальное значение соотношения между хлорофиллами а к b выявлено у большинства сортов в пластинке нижерасположенных листьев, как правило, первом или втором. Максимальное значение соотношения хлорофиллов а к b у большинства сортов отмечается у вышерасположенных листьев, включая флаговый.
Содержание каротиноидов в пластинке листьев составляло от 0,19 до 0,95 мг/г. Минимальное содержание каротиноидов отмечено у большинства сортов в пластинке нижних листьев. Максимальное содержание каротиноидов установлено у большинства сортов в пластинке верхних листьев, преимущественно флагового листа.
Величина соотношения между хлорофиллами и каротиноидами составляет 2,44–7,5. Минимальное значение соотношения между содержанием хлорофиллов и каротиноидов выявлено у всех сортов в пластинке 3-го или 5-го листьев. Максимальное значение соотношения хлорофиллов к каротиноидам у большинства сортов отмечено в пластинке нижних, 1–2-го листьев. Одноименные листья исследованных сортов имели различия по соотношению содержания хлорофиллов к каротиноидам (табл. 4). Наиболее существенные различия были отмечены в листовых пластинках 2-го, 3-го и 5-го метамеров.
Таблица 4
Сортовые особенности соотношения содержания хлорофиллов к каротиноидам (2004 г.)
| Сорта | Номер листа | Среднее | ||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | ||
| Лютесценс 62 | 4,33 ± 0,20 | 3,67 ± 0,10# | 3,19 ± 0,09# | 3,83 ± 0,10 # | 3,28 ± 0,10# | 4,17 ± 0,20 | 3,30 ± 0,10 | 3,68 |
| Саратовская 36 | 4,38 ± 0,20 | 4,45 ± 0,20* | 4,54 ± 0,20* | 4,45 ± 0,20* | 3,53 ± 0,10# | 3,70 ± 0,10#* | 4,31 ± 0,20* | 4,19 |
| Саратовская 52 | 5,72 ± 0,30* | 7,50 ± 0,30#* | 4,12 ± 0,20#* | 3,94 ± 0,10# | 2,62 ± 0,09#* | 3,88 ± 0,10# | 3,25 ± 0,10# | 4,43 |
| Нададорес 63 | 4,26 ± 0,20 | 4,46 ± 0,20* | 3,10 ± 0,09# | 4,33 ± 0,20* | 4,50 ± 0,20* | 4,09 ± 0,20 | 3,42 ± 0,10# | 4,02 |
| Саратовская 56 | 4,25 ± 0,20 | 3,76 ± 0,10# | 2,69 ± 0,09#* | 4,40 ± 0,20* | 2,44 ± 0,09#* | 3,93 ± 0,10# | 3,17 ± 0,09# | 3,52 |
| Саратовская 58 | 4,93 ± 0,30* | 3,84 ± 0,10# | 3,13 ± 0,09# | 3,96 ± 0,10# | 3,77 ± 0,10#* | 4,21 ± 0,20# | 3,78 ± 0,10#* | 3,95 |
Примечание: * - достоверная разница относительно сорта Лютесценс 62 при Р 0,05;
# - достоверная разница относительно первого листа при Р 0,05
В частности, в пластинке 2-го листа соотношение составляет от 3,67 у Лютесценс 62 до 7,50 у Саратовской 52. В пластинке 3-го листа величина его изменяется в пределах от 2,69 у Саратовской 56 до 4,54 у Саратовской 36, а в пластинке 5-го – от 2,44 у Саратовской 56 до 3,77 у Саратовской 58.
Таким образом, в пределах одноименных листьев могут наблюдаться сортовые различия по содержанию хлорофилла а, b и каротиноидов, величине соотношения между ними, что определяется генотипом сорта, степенью его пластичности и устойчивости к экзогенным факторам среды.
ВЫВОДЫ
1. Разнообразие типов клеток мезофилла пшеницы не исчерпывается числом ячеек. На основании наличия симметричности клетки в целом, наличия или отсутствие лопастей клетки, ширины цитоплазматического мостика, пространственного положения ячейки относительно продольной оси клетки, соотношения длины к ширине клетки их можно классифицировать на 11 основных типов.
2. Существует сортовая специфика по степени варьирования числа клеток мезофилла различных типов в листьях разных метамеров. Вместе с тем, имеет место общая для всех изученных сортов мягкой пшеницы закономерность в смене степени представительства клеток мезофилла разных типов в листовых пластинках разных метамеров. Мезофилл пластинки 1–3-го листьев представлен всеми типами клеток. В пластинке 4–7-го листьев разнообразие типов клеток мезофилла уменьшается, увеличивается доля клеток с выраженной ячеистой формой. В зависимости от сорта и положения листа на побеге доля клеток мезофилла различных типов может составлять от 2 до 50%.
3. Линейные параметры клеток мезофилла пластинки листьев мягкой яровой пшеницы отражают свойства генотипа сорта. Однако, как правило, длина клеток у всех сортов больше в листьях верхних 5–7-го метамеров. Минимальные значения ширины клеток мезофилла наблюдаются в пластинке верхнего, 7-го листа. Размер разных типов клеток мезофилла не обязательно положительно коррелирует с формой клетки или с числом ячеек в ней.
4. Определение среди дифференцирующейся паренхимы пластинки листа клеток мезофилла становиться возможным спустя 3,5 недели с момента начала роста листа, когда он достигнет длины 20–30 мм. Существенное увеличение длины клеток мезофилла пластинки происходит на 24–25 день вегетации одновременно с возрастанием абсолютной и относительной скорости роста листа в длину. С момента дифференциации клеток мезофилла происходит возрастание разнообразия типов клеток мезофилла. По достижении листом фазы завершения роста разнообразие типов клеток уменьшается, что свидетельствует об унификации структуры мезофилла.
5. Содержание хлорофиллов а и b, каротиноидов в пластинках листьев отражает их положение в системе метамеров побега, а также агроклиматические условия в период вегетации.
6. В пластинке листьев содержание хлорофиллов а и b составляет от 0,81 до 3,96 мг/г. Минимальное или максимальное содержание хлорофиллов может наблюдаться в различных листьях, однако, у большинства сортов максимум содержания хлорофиллов отмечается в верхних 5–7-ом листьях. Величина соотношения между хлорофиллами а к b составляет 1,19–5,53. Минимальное значение соотношения между хлорофиллами а к b выявлено у большинства сортов в пластинке первого или второго листа, максимальное – у вышерасположенных листьев, включая верхние.
7. Содержание каротиноидов в пластинке листьев составляет от 0,19 до 0,95 мг/г. Минимальное содержание каротиноидов отмечено у большинства сортов в пластинках нижних листьев. Максимальное – в пластинке верхних листьев, преимущественно флагового листа. Величина соотношения между хлорофиллами и каротиноидами составляет 2,44–7,5. Минимальное значение соотношения между содержанием хлорофиллов и каротиноидов выявлено у всех сортов в пластинке 3 или 5-го листьев, максимальное – у большинства сортов отмечено в пластинке нижних, 1-го и 2-го листьев.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ
- публикации в изданиях, рекомендуемых перечнем ВАК РФ
1. Даштоян Ю.В. Содержание пигментов фотосинтеза первого листа некоторых видов и сортов пшеницы / Ю.В. Даштоян, Д.А. Хакалова, М.В. Иванова, С.А. Степанов // Вопросы биологии, экологии, химии и методики обучения: Сб. науч. ст. Вып. 7. – Саратов, 2004. – С. 93 – 96.
2. Степанов С.А. Качественные аспекты анатомо-морфологической организации зародыша зерновки яровой пшеницы / С.А. Степанов, Ю.В. Даштоян // Бюллетень Бот. сада СГУ. Вып.3. – Саратов: «Научная книга», 2004 г. – С. 149 – 158.
3. Даштоян Ю.В. Влияние изменения площади листьев на рост и развитие побега пшеницы / Ю.В. Даштоян, Е.К. Щеглова, С.А. Степанов //Труды VII Международной конференции по морфологии растений, посвященной памяти И.Г. и Т.И. Серебряковых /Под ред. А.Г. Еленевского. – М.: МПГУ, 2004. – С. 77 – 78.
4. Даштоян Ю.В. Рост и развитие листьев в онтогенезе яровой пшеницы / Ю.В. Даштоян, С.А. Степанов // Вавиловские чтения – 2004: сборник работ. – Саратов: изд-во СГАУ, 2004. – С. 132 – 135.
5. Степанов С.А. Трансформация межметамерных отношений в онтогенезе побега пшеницы / С.А. Степанов, В.В. Коробко, Ю.В. Даштоян // Известия СГУ. Серия Химия, биология, экология. Вып.2. – Саратов, 2005. – Т. 5. – С. 33 – 36.
6. Танайлова Е.А. Видовые и сортовые особенности развития листьев эмбрионального побега зародышей зерновок пшеницы / Е.А. Танайлова, Ю.В. Даштоян, В.В. Коробко // Исследования молодых ученых и студентов в биологии: сб. научных трудов. Вып.3. – Саратов: изд-во СГУ, 2005. – С. 99 – 103.
7. Даштоян Ю.В. Ярусная изменчивость структуры мезофилла пластинки листа яровой пшеницы / Ю.В. Даштоян // Наука. Образование. Молодежь: Материалы IV Всероссийской научной конференции молодых ученых (8 – 9 февраля 2007 года). Часть 1. – Майкоп: изд-во АГУ, 2007. – С. 41 – 43.
8. Степанов С.А. Ярусная изменчивость содержания хлорофилла листьев пшеницы / С.А. Степанов, Ю.В. Даштоян // Современная физиология растений: от молекул до экосистем: Материалы докладов Международной конференции (в трех частях). Часть 3. (Сыктывкар, 18 – 24 июня 2007 г.). – Сыктывкар, 2007. – C. 418 – 419.
9. Степанов С.А. Метамерные особенности развития проводящих тканей пластинки листьев пшеницы / С.А. Степанов, Ю.В. Даштоян, Д.А. Хакалова // Бюллетень Бот. сада СГУ. – Саратов, 2007. – Вып. 6. – С. 151 – 156.
10. Даштоян Ю.В. Метамерная изменчивость состава и содержания пигментов фотосинтеза листьев пшеницы / Ю.В. Даштоян, Н.В. Меринова, С.А. Степанов // Вестник Саратовского госагроуниверситета им. Н.И. Вавилова. – Саратов: изд-во СГАУ, 2008. – Вып. 2. – С. 24 – 25.