

Хирономиды (diptera, chironomidae) бассейна нижнего амура. фауна, систематика, распространение
24
решения. Материалы межрегиональной научно-практической конференции: в 2 кн. (Хабаровск, 10–12 октября 2008 г.). Хабаровск: ДВО РАН. 2008. Кн. 2. С. 483–486.
14. Макарченко Е.А., Макарченко М.А., Яворская Н.М. Комары-звонцы подсемейства Orthocladiinae (Diptera, Chironomidae) бассейна р. Амур // III Дружининские чтения: Комплексные исследования природной среды в бассейне реки Амур. Мат. межрегиональной научной конференции: в 2 кн. (Хабаровск, 6–9 октября 2009 г.). Хабаровск: ДВО РАН, 2009. Кн. 2. С. 211–215.
15. Яворская Н.М. Личинки хирономид (Diptera, Chironomidae) ручьев Еловый и Змеиный и их роль в питании рыб (Нижнее Приамурье) // III Дружининские чтения: Комплексные исследования природной среды в бассейне реки Амур. Мат. межрегиональной научной конференции: в 2 кн. (Хабаровск, 6–9 октября 2009 г.). Хабаровск: ДВО РАН, 2009. Кн. 2. С. 291–294.
Яворская Надежда Мякиновна
ХИРОНОМИДЫ (DIPTERA, CHIRONOMIDAE)
БАССЕЙНА НИЖНЕГО АМУРА.
ФАУНА, СИСТЕМАТИКА, РАСПРОСТРАНЕНИЕ
АВТОРЕФЕРАТ
диссертации на соискание учёной степени
кандидата биологических наук
На правах рукописи
Яворская Надежда Мякиновна
ХИРОНОМИДЫ (DIPTERA, CHIRONOMIDAE)
БАССЕЙНА НИЖНЕГО АМУРА.
ФАУНА, СИСТЕМАТИКА, РАСПРОСТРАНЕНИЕ
03.00.09 – энтомология
АВТОРЕФЕРАТ
диссертации на соискание учёной степени
кандидата биологических наук
![]()
Владивосток – 2010
Работа выполнена в Лаборатории пресноводной гидробиологии Биолого-почвенного института ДВО РАН и в Лаборатории прикладной экологии Хабаровского филиала ФГУП «Тихоокеанский научно-исследовательский рыбохозяйственный центр».
Научный руководитель: доктор биологических наук,
старший научный сотрудник
Макарченко Евгений Анатольевич
Официальные оппоненты: доктор биологических наук,
старший научный сотрудник
Стороженко Сергей Юрьевич
доктор биологических наук,
профессор
Зинченко Татьяна Дмитриевна
Ведущая организация: Институт водных и экологических
проблем ДВО РАН, г. Хабаровск
Защита состоится « 21 » апреля 2010 г. в 13 часов на заседании диссертационного совета Д 005.003.03 при Биолого-почвенном институте ДВО РАН по адресу: 690022, г. Владивосток, проспект 100 лет Владивостоку, 159, факс: (4232) 310-193
С диссертацией можно ознакомиться в Центральной научной библиотеке ДВО РАН.
Автореферат разослан « » марта 2010 г.
| Учёный секретарь диссертационного совета, кандидат биологических наук | | Е.М. Саенко |
23
Статьи, опубликованные в отечественных периодических изданиях и региональных сборниках:
5. Макарченко Е.А., Макарченко М.А., Зорина О.В., Яворская Н.М. Предварительные данные по фауне хирономид (Diptera, Chironomidae) бассейна реки Амур // Пресноводные экосистемы бассейна р. Амур: Сб. науч. тр. / Под общ. ред. Е.А. Макарченко. – Владивосток: Дальнаука, 2008. С. 187–208.
6. Яворская Н.М. Личинки хирономид (Diptera, Chironomidae) реки Кади (Нижнее Приамурье) // Пресноводные экосистемы бассейна реки Амур: Сб. науч. тр. / Под общ. ред. Е.А. Макарченко. – Владивосток: Дальнаука, 2008. С. 209–217.
Работы, опубликованные в материалах всероссийских и международных конференций:
7. Яворская Н.М. Личинки хирономид (Diptera, Chironomidae) как виды–индикаторы состояния среды // Ресурсы и экологические проблемы Дальнего Востока. Мат. межрегиональной научно-практической конференции (Хабаровск, 2022 февраля 2006 г.) / Под ред. В.Т. Тагировой. – Хабаровск: ДВГГУ, 2006. С. 89–95.
8. Яворская Н.М. Структурная характеристика сообщества хирономид (Diptera, Chironomidae) р. Кади (Нижнее Приамурье) // Биология внутренних вод / Тез. докл. XIII Международной молодёжной школы-конференции (Борок, 23–26 октября 2007 г.). Борок, 2007. С. 78.
9. Яворская Н.М. Фауна амфибиотических насекомых реки Пей–Хой (Нижнее Приамурье) // Природные ресурсы и экологические проблемы Дальнего Востока: межрегиональная научно-практическая конференция (20–21 февраля 2007 г., г. Хабаровск). Хабаровск: ДВГГУ. 2007. С. 188–192.
10. Яворская Н.М. Бентофауна реки Мачтовая (Нижнее Приамурье) // Современное состояние водных биоресурсов: научная конференция посвященная 70-летию С.М. Коновалова (25–27 марта, 2008 г., Владивосток). Владивосток: ТИНРО–центр. 2008. С. 698–699.
11. Яворская Н.М. Зообентос реки Анастасьевка (Нижнее Приамурье) // Экологические проблемы крупных рек4: тезисы докладов международной конференции (8–12 сентября 2008 г., г. Тольятти, Россия). Тольятти: ИЭВБ РАН. 2008. С. 204.
12. Яворская Н.М. Личинки хирономид (Diptera, Chironomidae) реки Хиванда (Нижнее Приамурье) // Актуальные проблемы экологии, морской биологии и биотехнологии. Материалы VIII рег. конф. по актуальным проблемам экологии, биологии и биотехнологии студентов, аспирантов вузов и научных организаций Дальнего Востока России (Владивосток, 11–13 декабря 2008 г.). Владивосток: ДВГУ. 2008. C. 164–166.
13. Яворская Н.М. Донные беспозвоночные лососевой реки Саласу (Нижнее Приамурье) // Регионы нового освоения: экологические проблемы, пути
22
5. Изучение распределения хирономид по 5 категориям водных объектов показало, что наибольшее число видов обитает в самых малых (109 видов) и очень малых (99 видов) водотоках. В средних и малых водотоках зарегистрировано по 70 видов, в водоемах – 34 вида. Во всех категориях водных объектов доминируют представители подсем. Orthocladiinae, по типам ареалов – голаркты.
6. Фауна хирономид басс. Нижнего Амура (330 видов) по числу видов наиболее близка таковым о-ва Сахалин (329 видов) и басс. Среднего Амура (312 видов), но беднее фауны Приморского края (426 видов). Наибольшее сходство фауны басс. Нижнего Амура наблюдается c фаунами басс. Среднего Амура и Приморского края, наименьшее – с фауной о-ва Сахалин. Орографы включения показали «оригинальность» фауны бассейнов Нижнего и Среднего Амура, Приморского края и «банальность» – о-ва Сахалин.
7. Для хирономид басс. Нижнего Амура выделено 9 фенологических групп. Наибольшее число изученных хирономид (214 видов) имеет одну генерацию в год. Наивысшие пики активности лёта имаго – летний (основной) и весенне-летний.
8. В рационах рыб басс. Нижнего Амура выявлено 57 видов и форм личинок хирономид из 5 подсемейств, которые встречаются в пище 33 видов рыб. Установлено, что хирономиды являются наиболее излюбленным кормовым объектом молоди кеты.
По теме диссертации опубликованы следующие работы:
Статьи, опубликованные в ведущих рецензируемых научных журналах:
1. Макарченко Е.А., Яворская Н.М. Новый вид хирономид Monodiamesa kamora sp.n. (Diptera, Chironomidae, Prodiamesinae) из бассейна р. Амур // Евразиатский энтомологический журнал. Новосибирск–Москва, 2008. Т. 7. Вып. 2. С. 175–176.
2. Яворская Н.М. Личинки хирономид (Diptera, Chironomidae) рек бассейна р. Амур и материкового побережья Татарского пролива // Известия ТИНРО. Владивосток, 2008. Т. 152. С. 201–214.
3. Макарченко Е.А., Макарченко М.А., Яворская Н.М. Новые находки хирономид (Diptera, Chironomidae, Orthocladiinae) на Дальнем Востоке и сопредельных территориях. VI. Hydrobaenus Fries // Евразиатский энтомологический журнал. Новосибирск–Москва, 2009. Т. 8. Прил. 1. С. 33–50.
4. Яворская Н.М., Макарченко М.А. Первые сведения по фауне хирономид (Diptera, Chironomidae) Большехехцирского государственного природного заповедника (Хабаровский край) // Евразиатский энтомологический журнал. Новосибирск–Москва, 2009. Т. 8. Прил. 1. С. 93–96.
3
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность исследований. Хирономиды (Chironomidae), или комары–звонцы – всесветно распространённое семейство длинноусых двукрылых насекомых с полным метаморфозом, процветающее в современных условиях благодаря своей экологической пластичности. Взрослые насекомые эфемерны, имеют редуцированный ротовой аппарат и не питаются. Развитие куколки при благоприятных условиях завершается в течение суток. В стадии личинки хирономиды проводят наиболее длительный период жизни, который составляет от нескольких недель до двух лет. Они освоили лотические и лентические системы, пресноводные и гипергалинные водоёмы, а также периферические части океана и некоторые наземные местообитания с высокой влажностью.
В благоприятных условиях обитания личинки хирономид доминируют над другими донными беспозвоночными рек и озёр, играют важную роль в сообществах, перерабатывая органическое вещество в минеральное, а также участвуют в самоочищении водной среды, являются ценным кормом для бентосоядных рыб. Эти амфибиотические насекомые успешно используются учёными в качестве индикаторов степени загрязнения рек (Балушкина, 1976; Зинченко, 2009) и трофности озёр (Sther, 1975, 1979). В ядрах клеток слюнных желез личинки имеют политенные хромосомы и поэтому широко применяются в лабораторных исследованиях цитогенетиками и молекулярными биологами. При массовом вылете и роении комары–звонцы вызывают у людей, живущих в окрестностях больших рек и озёр, различные аллергии, в том числе бронхиальную астму, сезонные риниты и коньюктивиты (Kawai, Konishi, 1986, 1988).
Оценка продуктивности водоёмов и определение их пригодности для интродукции тех или иных видов или пород рыб, составление прогнозов биологической динамики искусственных водоёмов невозможны без знания систематического состава и особенностей экологии и биологии хирономид (Линевич, 1981).
В настоящее время для мировой фауны хирономид известно не менее 5000 видов из 440 родов и 11 подсемейств, для Палеарктики зарегистрировано более 1500 видов (Ashe, Cranston, 1990; Sther et al., 2000), для Неарктики – 1051 вид из 205 родов (Oliver et al., 1990). В России по предварительным данным обитает более 900 видов (Макарченко, Макарченко, 2009). Фауна хирономид российского Дальнего Востока представлена 736 видами (Макарченко и др., 2005). Всего для водотоков и водоёмов басс. р. Амур (без оз. Ханка) к настоящему времени зарегистрировано 404 вида (Макарченко и др., 2008).
Несмотря на относительно большое количество таксономических публикаций в последние несколько лет, фауна хирономид басс. Нижнего Амура изучена еще недостаточно, слабо разработана систематика отдельных подсемейств и многих родов, для большинства видов отсутствуют данные по пре-
4
имагинальным стадиям развития, а также их биологии, плохо изучено географическое распространение. Всё изложенное выше повлияло на выбор тематики настоящего исследования и определило его цель и задачи.
Цель и задачи исследования. Цель наших исследований – изучить фауну, систематику и распространение хирономид басс. Нижнего Амура.
Для достижения этой цели были поставлены следующие задачи:
- Изучить морфологию личинок, куколок и имаго самцов хирономид.
- Выявить таксономический состав и определить таксономическую структуру изученных хирономид, составить аннотированный список видов басс. Нижнего Амура.
- Изучить систематику наиболее богатого видами, но слабо изученного подсем. Orthocladiinae, составить определительные таблицы родов и видов этого подсемейства для басс. р. Амур.
- Изучить географическое распространение, выявить типы ареалов, обнаруженных в басс. Нижнего Амура видов.
- Изучить распределение хирономид по водным объектам басс. Нижнего Амура.
- Провести фаунистический анализ хирономид басс. Нижнего Амура в сравнении с таковыми некоторых сопредельных территорий.
- Выявить некоторые особенности биологии комаров–звонцов.
- Показать значение хирономид в питании рыб.
Научная новизна. Впервые обобщены оригинальные и литературные данные по фауне, систематике и распространению хирономид басс. Нижнего Амура, для которого выявлено 330 видов, относящихся к 107 родам шести подсемейств. Составлен аннотированный список видов. Описаны 2 новых для науки вида, 2 вида впервые указаны для Палеарктики, 7 видов оказались новыми для фауны России, 7 – фауны Дальнего Востока России, 4 рода и 21 вид – новыми для басс. Нижнего Амура. Подготовлена новая версия определительной таблицы родов и видов подсем. Orthocladiinae для басс. р. Амур по имаго самцам. Выявлены основные ареалы для 302 видов хирономид басс. Нижнего Амура. Проведён анализ сходства фаун басс. Нижнего Амура и сопредельных территорий. Впервые изучена фауна хирономид Большехехцирского государственного природного заповедника, для которого зарегистрирован 81 вид, из 40 родов четырех подсемейств. Обобщены данные по фенологии хирономид басс. Нижнего Амура и их встречаемости в питании 33 видов рыб.
Теоретическая и практическая ценность. Обобщен обширный материал по фауне хирономид басс. Нижнего Амура. Выявлены закономерности зоогеографического и таксономического разнообразия хирономид. Полученные фаунистические данные могут быть включены в государственный кадастр животного мира Хабаровского края. Выявленный видовой состав фауны хирономид на территории Большехехцирского государственного природного заповедника использован для составления «Летописи природы». Составлен-
21
1959; Мухачёва, 1950; Кохменко, 1965; Леванидов, 1959; Тиунова и др., 2008; и др.).
Из басс. Нижнего Амура в пище 33 видов рыб обнаружено 57 видов хирономид, относящихся к 5 подсемействам: Orthocladiinae (17 родов), Chironominae (10 родов), Diamesinae (5 родов), Prodiamesinae (3 рода) и Tanypodinae (2 рода). В пищевом рационе рыб доминируют представители подсем. Orthocladiinae (51%) и Chironominae (33%), второстепенными являются виды из подсемейств Diamesinae (9%), Prodiamesinae (4%) и Tanypodinae (3%).
Больше всего в водотоках басс. Нижнего Амура личинками хирономид питается молодь кеты, у которых в пищевом комке их бывает до 100% по частоте встречаемости и 99% по массе. По сравнению с личинками, куколки хирономид играют меньшую роль в питании, достигая в желудках молоди этих лососей до 18,6% по массе (Константинов, 1950). Другие бентосоядные рыбы помимо хирономид употребляют в пищу личинок веснянок, поденок и ручейников, однако, хирономиды всё равно остаются неотъемлемой составной частью их пищевого рациона. Так, по частоте встречаемости в питании ленка и хариуса личинки хирономид составляют до 100%, мальмы – до 72,2%, бычка-подкаменщика – до 8%, колюшки до 36% (Константинов, 1950; Леванидов, Леванидова, 1962; Кохменко, 1962, 1964).
Выводы
1. Для басс. Нижнего Амура зарегистрировано 330 видов хирономид из 107 родов, относящихся к 6 подсемействам – Podonominae (2 рода, 2 вида), Tanypodinae (7 родов, 15 видов), Diamesinae (8 родов, 17 видов), Prodiamesinae (3 рода, 6 видов), Orthocladiinae (50 родов, 173 вида), Chironominae (37 родов, 117 видов).
2. Описано 2 новых для науки вида, 2 вида впервые указаны для Палеарктики, 7 видов оказались новыми для фауны России, 7 – фауны Дальнего Востока России, 4 рода и 21 вид – новыми для басс. Нижнего Амура. Составлен аннотированный список видов.
3. Изучена систематика доминирующего в донных сообществах басс. Нижнего Амура подсем. Orthocladiinae, для которого выявлено 173 вида. По имаго самцам составлена определительная таблица для 51 рода и 207 видов, обнаруженных в бассейне всего Амура.
4. Выявлено 7 типов ареалов для 302 видов. Преобладают палеарктические виды (173 или 57,3% ) над голарктическими (129 видов или 42,7%). Из палеарктических видов доминируют восточно-палеарктические материково-островные (55 видов или 31,8% от всех палеарктов), восточно-палеарктические материковые (44 вида или 25,4%) и трансевразиатские полидизъюнктивные виды (36 видов или 20,8%).
20
рая – на июнь–август и сентябрь, или первая генерация наблюдается в мае, вторая – в октябре; первая генерация также может быть в июне, вторая – в сентябре; иногда обе генерации приходятся на летние месяцы. Уникален выход имаго из куколок с декабря до начала марта у бивольтинного вида Diamesa leona Roback.
Таблица 6
Распределение видов хирономид бассейна Нижнего Амура по типам генераций
| Подсемейство хирономид | Бивольтинные | Моновольтинные | Поливольтинные | |
| Chironominae | 40 | 58 | 5 | |
| Diamesinae | 5 | 11 | – | |
| Orthocladiinae | 27 | 132 | 4 | |
| Podonominae | – | 1 | 1 | |
| Prodiamesinae | 1 | 5 | – | |
| Tanypodinae | 3 | 7 | 2 | |
| Всего видов: | 76 | 214 | 12 | |
К третьей группе относятся поливольтинные виды, которые в условиях басс. Нижнего Амура могут иметь более двух поколений в год. Вылет имаго у них происходит ранней весной – с начала марта по май, в течение летних месяцев – июнь–июль, осенью – сентябрь–октябрь и редко в зимний период – в декабре. В эту группу мы включили 12 видов хирономид – Ablabesmyia monilis, Lasiodiamesa sphagnicola, Procladius (H.) ferrugineus, Limnophyes edwardsi, L. natalensis, Orthocladius (Orthocladius) setosus, Parachironomus arcuatus, Parametriocnemus stylatus, Endochironomus tendens, Paratendipes albimanus, Chironomus (Chironomus) yoshimatsui, Cladotanytarsus wixionensis.
ГЛАВА 9. ХИРОНОМИДы в питании рыб
Обобщены литературные данные по потреблению хирономид рыбами Дальнего Востока России. Оказалось, что в рационе 68 видов дальневосточных рыб встречаются 328 видов и форм личинок и куколок хирономид относящихся к 5 подсемействам: Orthocladiinae (36 родов), Chironominae (29 родов), Diamesinae (9 родов), Prodiamesinae (3 рода), Tanypodinae (9 родов); плюс неопределенные формы. Доминируют в питании рыб виды из подсем. Chironominae (43%) и Orthocladiinae (29%), второстепенными являются Diamesinae (10%), Tanypodinae (7%), Prodiamesinae (3%) (Константинов, 1951,
5
ные определительные таблицы родов и видов хирономид подсем. Orthocladiinae могут быть использованы сотрудниками природоохранных и рыбохозяйственных учреждений Хабаровского края при проведении экологических экспертиз и изучениИ кормовой базы бентосоядных рыб. Фаунистические данные важны при оценке и сохранении биологического разнообразия, а также при составлении региональных Красных книг, где в качестве охраняемых видов животных должны быть включены редкие виды хирономид. Личинки хирономид являются важным пищевым ресурсом для бентосоядных рыб и их молоди, что особенно актуально для басс. Нижнего Амура, имеющего большинство притоков, относящихся к высшей рыбохозяйственной категории. Знания особенностей биологии и экологии личинок хирономид помогут оценить продуктивность водоёмов и в дальнейшем определить их пригодность для интродукции тех или иных пород и видов рыб.
Личное участие в получении научных результатов: В работе использованы материалы, собранные лично автором на 43 реках, 14 ручьях, 10 протоках, 17 озёрах (235 сборов имаго, 87 качественных сборов личинок и куколок), а также данные лаборатории пресноводной гидробиологии БПИ ДВО РАН. Для выявления видовой принадлежности преимагинальных стадий развития автором сделано 163 выведения комаров из личинок четвёртого возраста и куколок. Изготовлено более 400 микропрепаратов для идентификации личинок, куколок и имаго. Собранные автором пробы на кариоанализ обработаны к.б.н. А.Г. Истоминой (Институт цитологии и генетики СО РАН, г. Новосибирск). Планирование работ и выполнение экспериментов проведены самостоятельно. Интерпретация полученных результатов проведена совместно с научным руководителем д.б.н. Е.А. Макарченко. Все литературные данные, использованные в работе, имеют ссылки на источники.
Апробация работы. Результаты работы были представлены на межрегиональной научно-практической конференции «Ресурсы и экологические проблемы Дальнего Востока» (Хабаровск, 2006); Международной молодёжной школе-конференции «Биология внутренних вод» (Борок, 2007); межрегиональной научно-практической конференции «Природные ресурсы и экологические проблемы Дальнего Востока» (Хабаровск, 2007); Чтениях памяти В.Я. Леванидова (Владивосток, 2008); научной конференции, посвященной 70-летию С.М. Коновалова «Современное состояние водных биоресурсов» (Владивосток, 2008); международной конференции «Экологические проблемы крупных рек–4» (Тольятти, 2008); региональной конференции по актуальным проблемам экологии, биологии и биотехнологии студентов, аспирантов ВУЗов и научных организаций Дальнего Востока России (Владивосток, 2008); межрегиональной научно-практической конференции «Регионы нового освоения: экологические проблемы, пути решения» (Хабаровск, 2008); Дружининских чтениях (Хабаровск, 2009); Отделе зоологии Биолого-почвенного института ДВО РАН (Владивосток, 2010).
6
Публикации. По теме диссертации опубликовано 15 работ.
Структура и объём диссертации. Диссертация состоит из введения, 9 глав, выводов, списка литературы, состоящего из 254 наименований (в том числе 69 на иностранных языках). Работа изложена на 266 страницах и иллюстрирована 12 таблицами и 30 рисунками.
Благодарности. Выражаю искреннюю благодарность моему научному руководителю, зав. лабораторией пресноводной гидробиологии Биолого-почвенного института ДВО РАН (БПИ ДВО РАН), д.б.н. Е.А. Макарченко за ценные консультации и советы, помощь на всех этапах планирования, выполнения и написания работы, а также за помощь в определении хирономид подсемейств Diamesinae, Podonominae и Prodiamesinae. Выражаю сердечную признательность за помощь в определении материала и ценные консультации по систематике подсемейств Orthocladiinae и Chironominae М.А. Макарченко и
к.б.н. О.В. Зориной (Лаборатория пресноводной гидробиологии БПИ ДВО РАН, г. Владивосток). За проведение кариологического анализа и определение видов рода Chironomus автор благодарен к.б.н. А.Г. Истоминой (Институт цитологии и генетики СО РАН, г. Новосибирск). В течение всей работы автору оказывалась постоянная помощь и поддержка сотрудников института Хабаровского филиала ТИНРОцентра (г. Хабаровск). Автор искренне благодарен и признателен директору института Г.В. Новомодному и зав. лабораторией прикладной экологии Н.М. Панасенко за содействие, поддержку и организацию выполнения работы, зав. музеем–аквариумом «Рыбы Амура» Е.В. Новомодному за ценные консультации и предоставленную литературу по энтомологии. Автор считает приятным долгом выразить благодарность энтомологу-любителю В.Я. Федорову за помощь, оказанную в ходе выполнения работ в экспедициях, директору Большехехцирского государственного заповедника (г. Хабаровск) С.В. Спиридонову и экс-заместителю директора Г.П. Телицыну за предоставленную возможность изучить фауну хирономид заповедника, а также научным сотрудникам заповедника А.М. Долгих и к.б.н. К.Н. Ткаченко за оказанную помощь в ходе выполнения работ.
СОДЕРЖАНИЕ РАБОТЫ
ВВЕДЕНИЕ
Во введении приведены общие сведения о семействе Chironomidae, обоснована актуальность темы, сформулированы цель и задачи исследования, приведены научная новизна, теоретическое и практическое значение работы.
ГЛАВА 1. ОБЗОР ЛИТЕРАТУРЫ
В главе рассмотрены история изучения фауны и систематики хирономид, результаты современных исследований кариологии, биологии и метаморфоза комаров-звонцов, а также проблемы, связанные с построением естественной системы семейства по личинкам, куколкам и имаго. Проанализированы все
19
ГЛАВА 8. НЕКОТОРЫЕ ОСОБЕННОСТИ БИОЛОГИИ ХИРОНОМИД БАССЕЙНА НИЖНЕГО АМУРА
Сезон лёта имаго хирономид в басс. Нижнего Амура практически длится 7,5 месяцев, начинается со второй декады апреля и продолжается до начала
ноября. Первые хирономиды появляются в конце апреля и представлены видами родов Chaetocladius, Hydrobaenus, Limnophyes, Metriocnemus, Orthocladius, Pseudosmittia, Smittia; последние вылеты комаров-звонцов (рода Orthocladius) зарегистрированы нами в начале ноября. По срокам лёта имаго хирономид в басс. Нижнего Амура выделено 9 групп.
Большинство хирономид входит в группы летних (196 видов – 64,9%) и весенне-летних (51 вид – 16,9%) видов. Далее следуют весенняя (25 видов – 8,3%), летне-осенняя (12 видов – 4%), осенняя (7 видов – 2,3%) и весенне-летне-осенняя (7 видов – 2,3%) группы, зимне-весенне-летне-осенняя (2 вида – 0,7%). Оставшиеся весенне-осенняя и зимне-весенняя группы представлены наименьшим количеством видов и составляют каждая по 1 виду – по 0,3% (рис. 7).
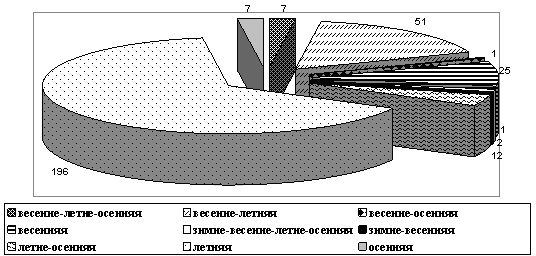
Рис. 7. Распределение числа видов хирономид по фенологическим группам
По числу генераций в год изученные хирономиды басс. Нижнего Амура поделены на три группы видов, среди которых доминируют виды, имеющие одну генерацию в год (табл. 6).
К первой группе относятся моновольтинные виды, которые имеют только одно поколение в год и вылет имаго происходит у них обычно с весны до осени. В басс. Нижнего Амура обнаружено 214 моновольтинных видов хирономид.
Ко второй группе относятся бивольтинные виды, которых в настоящее время в басс. Нижнего Амура насчитывается всего 76. Они имеют два поколения в год – первая генерация приходится на март–апрель, начало мая, вто-
18
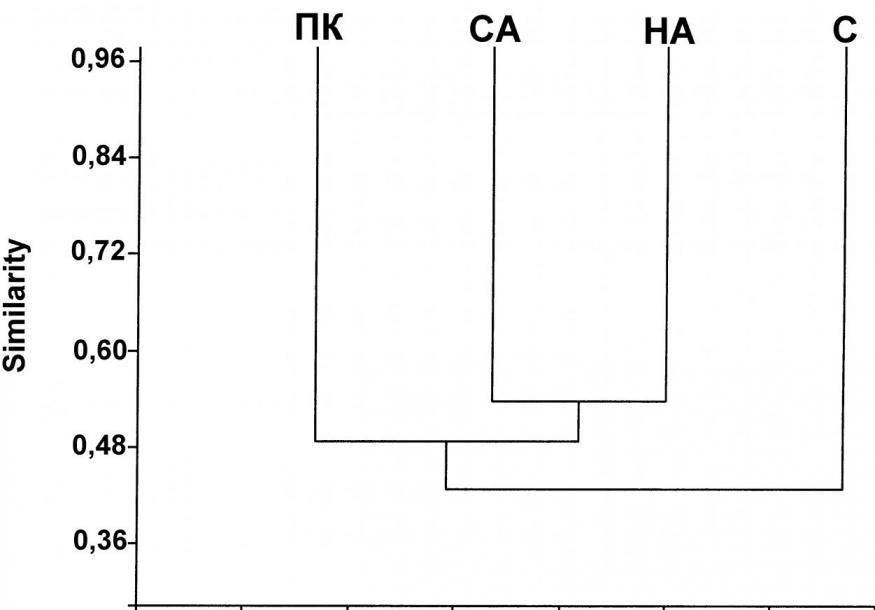
Рис. 5. Дендрограмма сходства фаун сопредельных территорий (UPGMA, ICS). Обозначения: ПК – Приморский край, СА – басс. Среднего Амура, НА – басс. Нижнего Амура, С – о-в Сахалин.
Для графического отображения матриц отношений сходства были построены ориентированные графы включения фаун сопредельных территорий. Анализ орографов включения фаун сопредельных территорий по общему числу видов показал что в Приморский край входит наибольшее число стрелок, следовательно, фауна этой территории является наиболее «оригинальной», а фауна о-ва Сахалин – «банальной». Фауна басс. Нижнего Амура занимает промежуточное положение (рис. 6).

Рис. 6. Орограф включения фаун сопредельных территорий по общему числу видов (при =0,5).
7
основные работы по изучению хирономид р. Амур и ее бассейна. Выделены главные этапы в исследовании комаров-звонцов российскими учеными в этом регионе. Наиболее значимый по полученным результатам период исследования фауны и систематики хирономид и других донных беспозвоночных р. Амур был во время проведения работ Амурской ихтиологической экспедиции в 1945–1949 гг., когда было выявлено для бассейна реки 127 видов и личиночных форм хирономид. В 1969 г. за счет обследования В.Я. Леванидовым полугорных участков крупных притоков список был расширен до 197 видов и личиночных форм (Леванидов, 1969). С 1974 г. было начато планомерное изучение фауны и систематики хирономид Дальнего Востока, в том числе и басс. р. Амур Е.А. Макарченко и М.А. Макарченко, а с 1998 г. и О.В. Зориной. В результате исследований этих хирономидологов к началу наших работ для басс. р. Амур было зарегистрировано 275 видов (Макарченко и др., 2005).
ГЛАВА 2. МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЙ
2.1. Материал
В основу диссертационной работы положен материал, собранный автором в период с 2005 по 2009 гг. в Хабаровском, Амурском, им. Лазо, Нанайском, Комсомольском, Ульчском и Николаевском районах Хабаровского края, а также в Смидовичском р-не Еврейской Автономной области (рис. 1). Всего было обследовано 25 озер, 40 рек, 10 проток р. Амур, 29 ручьев, 3 болота и 2 пруда. Кроме этого использованы данные лаборатории пресноводной гидробиологии Биолого-почвенного института ДВО РАН. Всего за время исследований автором было обработано более 250 проб, просмотрено свыше 2 тыс. экз. насекомых, изготовлено более 400 микропрепаратов.
Для выявления видовой принадлежности преимагинальных стадий развития проводили выведение комаров из личинок четвёртого возраста и куколок. Всего было сделано 163 выведения, в результате чего получено 85 экз. имаго, к сожалению 33 из них оказались самками.
Так как большинство видов рода Chironomus невозможно определить без кариоанализа, для его проведения отбирали личинок IV возраста, которых фиксировали в жидкости Карнуа (3:1). Всего для этой цели было собрано 270 личинок. Идентификация видов по политенным хромосомам выполнена к.б.н. А.Г. Истоминой в Институте цитологии и генетики СО РАН, г. Новосибирск.
2.2. Методы исследований
Методы сбора и обработки хирономид использовали по А.И. Шиловой (1976) и Е.А. Макарченко (1985, 2006). Полевые сборы материала проводили с марта по ноябрь, единично – в декабре. Комаров ловили энтомологическим сачком во время роения, «обкашиванием» прибрежной растительности, в тёмное время суток на свет, а также ловушкой Малеза. Качественные гидробиологические пробы личинок и куколок отбирали с помощью водного сачка. Материал фиксировали жидкостью Удеманса и 70%-м этанолом. При опреде-
8
лении видовой принадлежности хирономид делали постоянные микропрепараты на стеклах с использованием жидкости Фора–Берлезе.
Для определения хирономид, уточнения номенклатуры таксонов и установления ареалов были использованы статьи, определители, каталоги, монографии и ревизии отечественных и зарубежных хирономидологов (Шилова, 1976; Макарченко, 1985; Макарченко, Макарченко, 1999, 2006, 2008; Макарченко и др., 2005; Зорина, 2005, 2006; Сергеева, Макарченко, 2006; Langton, Pinder, 2007; Makarchenko, Makarchenko, 2008, 2009; и др.). Терминология в очерке по морфологии и в определительных таблицах дана по О.А. Сэзеру (Sther, 1980).
Для изучения распределения хирономид по водотокам использовали классификацию водных объектов А.М. Комлева и Е.А. Черных (1984), согласно которой по длине реки делятся на самые малые (0–10 км), очень малые (11–25 км), малые (26–100 км), средние (101–500 км), большие (501–1000 км), очень большие (свыше 1001 км). При выяснении биотопической приуроченности личинок хирономид использовали классификацию А.А. Линевич (1981).
Типы ареалов даны по К.Б. Городкову (1984). Сравнение фаун сопредельных территорий проводилось с использованием кластерного анализа (метод UPGMA), в программе Past ver. 1.57 с применением коэффициента Чекановского-Съёренсена, а также по методике В.Л. Андреева (1978), Ю.А. Песенко (1982), Б.И. Семкина и Л.С. Куликовой (1981).
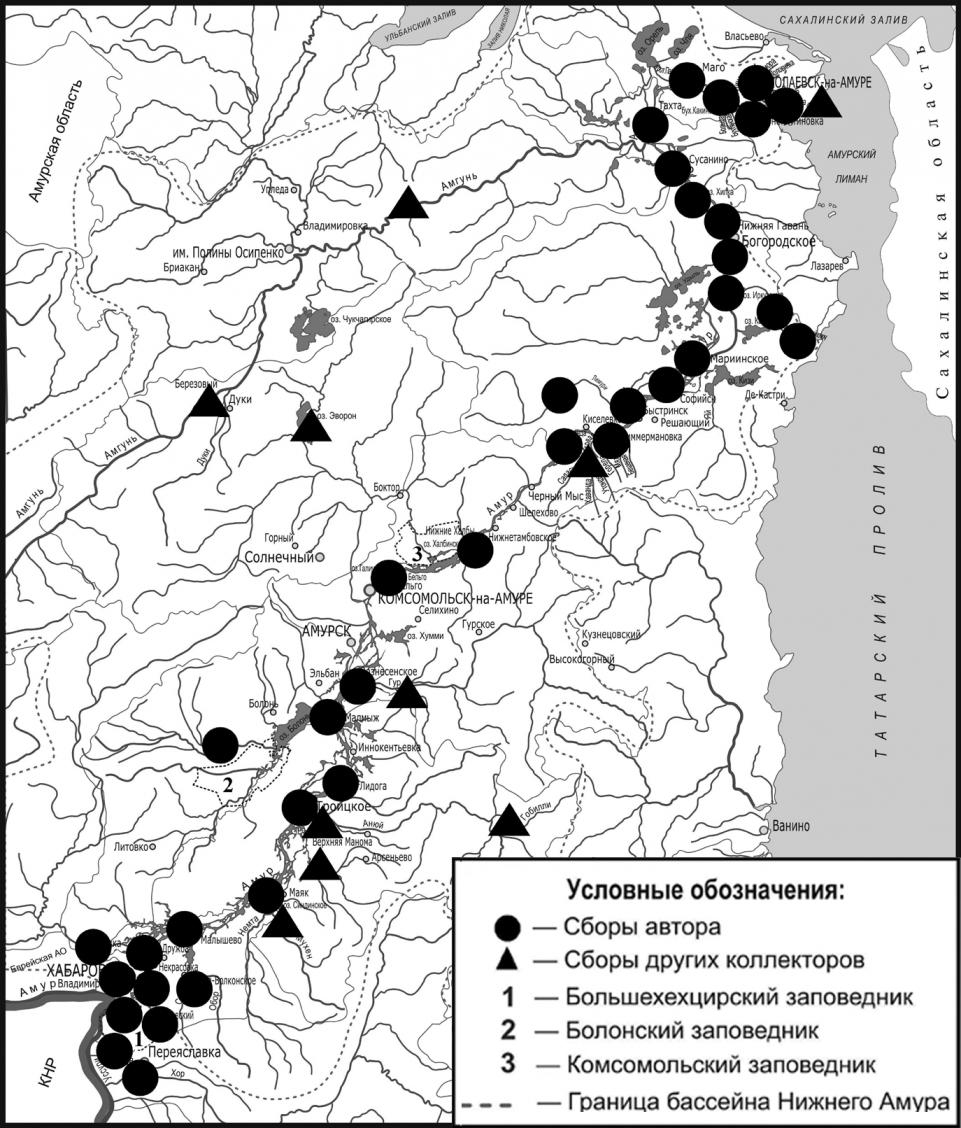
Рис. 1. Карта-схема района исследований с указанием мест сбора материала.
17
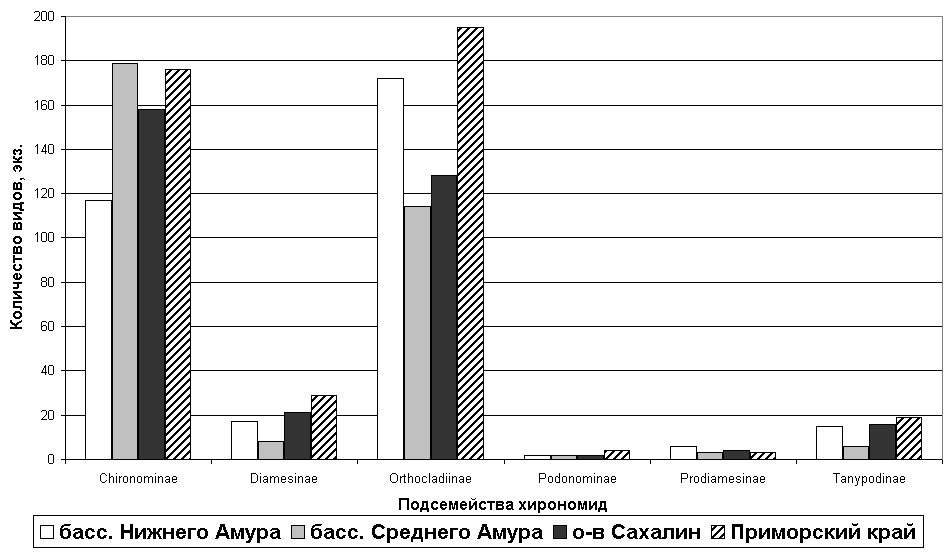
Рис. 4. Число видов в подсемействах Chironomidae басс. Нижнего Амура, басс. Среднего Амура, о-ва Сахалин и Приморского края.
Сравнение фаун хирономид басс. Нижнего Амура, басс. Среднего Амура, о-ва Сахалин и Приморского края показало, что коэффициенты их сходства по Чекановскому-Съёренсену высоки и составляют от 41% до 53% (табл. 5).
Таблица 5
Количество общих видов и показатель сходства видового состава хирономид
| Регион | Басс. Нижнего Амура, N = 330 |
| Басс. Среднего Амура с = 172, N = 312 | S = 53% |
| О–в Сахалин с = 138, N = 329 | S = 41% |
| Приморский край с = 186, N = 426 | S = 49% |
| Условные обозначения: с – количество общих видов, N – количество видов в регионе, S – показатель сходства. | |
Полученные данные показывают, что наибольшее сходство фауны басс. Нижнего Амура наблюдается c фаунами басс. Среднего Амура и Приморского края, наименьшее – с фауной о-ва Сахалин (рис. 5). Последнее объясняется в первую очередь слабой изученностью хирономид северо-западной части Сахалина, примыкающей к Амурскому лиману.
16
Таблица 4
Таксономическое разнообразие Chironomidae басс. Нижнего Амура и некоторых сопредельных территорий
| Таксоны | Территория | |||
| О-в Сахалин | Басс. Нижнего Амура | Басс. Среднего Амура | Приморский край | |
| Chironominae | 39/158 | 37/117 | 43/179 | 41/176 |
| Chironomini | 29/107 | 28/89 | 33/135 | 30/115 |
| Pseudochironomini | 1/1 | – | – | 1/1 |
| Tanytarsini | 9/50 | 9/28 | 10/44 | 10/60 |
| Diamesinae | 7/21 | 8/17 | 5/8 | 10/29 |
| Boreoheptagyiini | – | – | – | 1/2 |
| Diamesini | 6/19 | 7/16 | 4/7 | 7/25 |
| Protanypodini | 1/2 | 1/1 | 1/1 | 2/2 |
| Orthocladiinae | 37/128 | 50/173 | 39/114 | 52/195 |
| Podonominae | 2/2 | 2/2 | 2/2 | 4/4 |
| Prodiamesinae | 3/4 | 3/6 | 2/3 | 2/3 |
| Tanypodinae | 8/16 | 7/15 | 5/6 | 11/19 |
| Coelotanypodini | – | – | – | 1/2 |
| Macropelopini | 1/2 | – | – | 3/3 |
| Pentaneurini | 6/10 | 5/7 | 2/3 | 6/9 |
| Procladini | 1/4 | 1/6 | 2/2 | 1/5 |
| Tanypodini | – | 1/2 | 1/1 | – |
| Всего подсемейств: | 6 | 6 | 6 | 6 |
| Всего родов: | 96 | 107 | 96 | 120 |
| Всего видов: | 329 | 330 | 312 | 426 |
Примечание. В числителе – количество родов, в знаменателе – видов.
Среднюю нишу заняли два подсемейства: Diamesinae и Tanypodinae. Диамезины доминируют в Приморском крае (10 родов, 29 видов – 2%), хорошо они представлены и на о-ве Сахалин (7 родов, 21 вид – 2%), далее следует басс. Нижнего Амура (8 родов, 17 видов – 1 %) и меньше всего их обнаружено в басс. Среднего Амура (5 родов, 8 видов – 1%). Таниподины преобладают в Приморском крае (11 родов, 19 видов – 1%), на о-ве Сахалин количество их уже снижается (8 родов, 16 видов – 1%), потом следует басс. Нижнего Амура (7 родов, 15 видов – 1%) и меньше всего видов из данного подсемейства найдено в басс. Среднего Амура (5 родов, 6 видов – 0,4%). Последнюю по числу видов ступеньку занимают подсемейства Podonominae и Prodiamesinae. Их представители во всех указанных сопредельных территориях обнаружены от двух до шести видов – от 0,1 до 0,4%.
9
ГЛАВА 3. ФИЗИКО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА
БАССЕЙНА НИЖНЕГО АМУРА
Территория, на которой проводились исследования, расположена на юге Хабаровского края (рис. 1). Согласно Лоции рек Амурского бассейна (1968), басс. Нижнего Амура расположен между параллелями 4750 и 5402 с.ш. и меридианами 13255 и 14125 в.д. Общая площадь бассейна составляет 208080 кв. км. Его естественные границы на севере проходят по хребтам Мевачан и Кивун, на западе – по хребтам Буреинский и Ям-Алинь; с юго-востока и востока он ограничен горным хребтом Сихотэ-Алинь. На основании литературных данных приведена гидрологическая характеристика р. Амур и обследованных водотоков и водоёмов басс. Нижнего Амура.
ГЛАВА 4. КРАТКИЙ Очерк морфологии Chironomidae
Рассмотрено строение основных структур имаго, куколки и личинки Chironomidae, используемых в диагностике подсемейств, родов и видов, приведены их сравнительные характеристики для представителей различных родов и видов. Основой для написания данной главы послужили данные, опубликованные в Определителе насекомых Дальнего Востока России (Макарченко, 2006) и глоссария морфологической терминологии хирономид (Sther, 1980).
Имаго самец. При идентификации родов и видов самцов наибольшее значение имеет строение глаз, клипеуса, степень развития максиллярного щупика и фронтальных бугорков головы, антенн, строение и хетотаксия груди, особенно передне- и среднеспинки, наличие или отсутствие шпор и пульвилл ног, жилкование крыльев, наличие и степень опушения чешуйки крыла и строение гениталий, которое часто имеет решающую роль при определении видов. В строении гипопигия необходимо особо обратить внимание на форму верхних и нижних придатков гонококсита, анального отростка, поперечной стернаподемы, наличие или отсутствие вирги.
Важное значение в диагностике видов имеют различные индексы некоторых выше перечисленных морфологических структур: AR (отношение длины последнего членика жгутика антенны к общей длине всех предыдущих члеников); LR (отношение длины ta1 к t на P1, P2 и P3), SV (отношение длин f+t к длине ta1 на P1, P2 и P3), BV (отношение суммы длин f+t+ta1 на P1, P2 и P3 к сумме длин ta2+ta3+ta4+ta5 на P1, P2 и P3), BR (отношение длины щетинок ta1 к минимальной ширине ta1), SVoR (отношение длины верхнего придатка, измеренной от его вершины до латеральной щетинки, к длине остальной части верхнего придатка, измеренной от латеральной щетинки до его конца), HR (отношение длины гонококсита к длине гоностиля).
Куколка. При определении хирономид по куколкам наибольшее значение имеет строение фронтальной апотомы (степень развития бугорков головы и фронтальных выростов, наличие фронтальных щетинок), строение торака-льных рогов и их форма, очертания и величина бугорков торакального
10
гребня, количество щетинок на груди, вооружение тергитов и стернитов брюшка, степень развития анального гребня или шпоры, строение анального сегмента (наличие шагрени, дорсальных щетинок и различных выростов или пучков щетинок, количество и степень развития плавательных щетинок).
Личинка. Хорошо определяются по строению структур головы (антенны, фронтальный, щёчный, гулярный и затылочный склериты, клипеус, лабрум, ментум, мандибулы, максиллы, лабральные пластинки, эпифаринкс, гипофаринкс, премандибулы, вентроментальные пластинки) и последних члеников брюшка (подталкиватели, папиллы, латеральные отростки). В диагностике видов оправдано применение антеннального индекса (AR) – отношение длины базального членика антенны к длине члеников жгутика.
ГЛАВА 5. ТаксономиЯ И СИСТЕМАТИКА хирономид
БАССЕЙНА Нижнего АМУРА
5.1. Таксономическая структура
Система хирономид принята по классификации O.А. Сэзера (Sther, 1977, 2000). В результате наших исследований для басс. Нижнего Амура выявлено 330 видов, относящихся к 107 родам, 7 трибам, 6 подсемействам – Podonominae (2 рода, 2 вида), Tanypodinae (7 родов, 15 видов), Diamesinae (8 родов, 17 видов), Prodiamesinae (3 рода, 6 видов), Orthocladiinae (50 родов, 173 вида), Chironominae (37 родов, 117 видов) (табл. 1).
Таблица 1
Таксономическая структура Chironomidae басс. Нижнего Амура
| Подсемейства хирономид | Число таксонов | ||
| Триб | Родов | Видов | |
| Podonominae | – | 2 | 2 |
| Tanypodinae | 3 | 7 | 15 |
| Diamesinae | 2 | 8 | 17 |
| Prodiamesinae | – | 3 | 6 |
| Orthocladiinae | – | 50 | 173 (20) |
| Chironominae | 2 | 37 | 117 (1) |
| Всего: | 7 | 107 | 330 (21) |
Примечание. В скобках указано число видов, впервые обнаруженных в басс. Нижнего Амура автором
Наибольшее число видов выявлено в родах Polypedilum (20 видов), Chironomus (9 видов), Cryptochironomus (7 видов) (подсем. Chironominae), Cricotopus (18 видов), Limnophyes (12 видов), Orthocladius (11 видов) (подсем. Orthocladiinae) и Procladius (6 видов) (подсем Tanypodinae). В других родах число видов значительно ниже и не превышает 5.
15
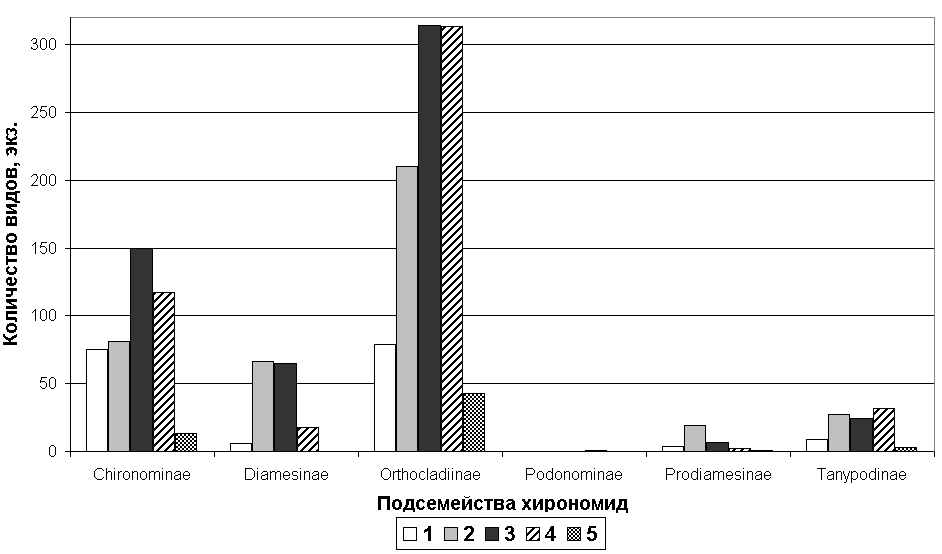
Рис. 3. Соотношение числа видов хирономид по подсемействам в водных объектах. Примечание: 1 – средние водотоки (1 категория), 2 – малые водотоки (2 категория), 3 – очень малые водотоки (3 категория), 4 – самые малые водотоки (4 категория), 5 – водоёмы (5 категория).
ГЛАВА 7. ФАУНА ХИРОНОМИД БАССЕЙНА НИЖНЕГО АМУРА
Фауна хирономид басс. Нижнего Амура представлена 330 видами и по сравнению с Приморским краем, где зарегистрировано 426 видов, характеризуется меньшим видовым разнообразием. Наиболее близкой по числу видов для басс. Нижнего Амура является фауна хирономид о-ва Сахалин (329 видов) и басс. Среднего Амура (312 видов) (табл. 4).
После анализа всех подсемейств оказалось, что наиболее крупными по количеству родов и видов в сопредельных территориях являются Chironominae и Orthocladiinae (рис. 4). Хирономины доминирует в басс. Среднего Амура (43 рода, 179 видов – 13% от общего числа в регионе) и в Приморском крае (41 род, 176 видов – 13%), потом следует о-в Сахалин (39 родов, 158 видов – 11%) и меньше всего обнаружено видов из данного подсемейства в басс. Нижнего Амура (37 родов, 117 видов – 8%). Подсем. Orthocladiinae хорошо представлено в Приморском крае (52 рода, 195 видов – 14%) и в басс. Нижнего Амура (50 родов, 173 вида – 12%), чуть меньшим видовым разнообразием отличается о-в Сахалин (37 родов, 128 видов – 9%), а также басс. Среднего Амура (39 родов и 114 видов – 8%).
14
В басс. Нижнего Амура нами впервые обнаружен 21 вид – Euryhapsis fuscipropes Sther et Wang, 1992, Eukiefferiella togaeutertia Sasa et Okazawa, 1992, Krenosmittia borealpina (Goetghebuer, 1944), K. toyamaquerea (Sasa, 1996), Nanocladius (Nanocladius) spiniplenus Sther, 1997, Paraphaenocladius penerasus (Edwards, 1929), Pseudorthocladius curtistylus Goerghebuer, 1921, Pseudosmittia rostriformis Makarchenko et Makarchenko, 2006, P. angusta (Edwards, 1929), Boreosmittia elevata Makarchenko et Makarchenko, 2009, P. longiseta (Thienemann, 1921), Trichosmittia hikosana Yamamoto, 1998, Bryophaenocladius flavoscutellatus (Malloch, 1915), B. pleuralis (Malloch, 1915), B. auritus Makarchenko et Makarchenko, 2006, Hydrobaenus biwaquartus (Sasa et Kawai, 1987), H. laticaudus Sther, 1976, H. sikhotealinensis Makarchenko et Makarchenko, 2006, Orthocladius (Orthocladius) defensus Makarchenko et Makarchenko, 2006, Chironomus (Chironomus) yoshimatsui Martin et Sublette, 1972, Parakiefferiella vshivkovae Makarchenko et Makarchenko, 2001. Наиболее интересными находками из этих видов в басс. Нижнего Амура являются H. laticaudus, который был известен только из типового местообитания на Аляске, B. pleuralis, указанный ранее лишь из Северной Америки, K. toyamaquerea и T. hikosana, впервые были обнаружены за пределами Японии.
Также, во время изучения фауны хирономид Большехехцирского государственного заповедника впервые для его территории зарегистрирован 81 вид из 40 родов 4 подсемейств – Tanypodinae (3 вида), Diamesinae (1 вид), Orthocladiinae (54 вида), Chironominae (23 вида) (Яворская, Макарченко, 2009).
6.2. Распределение по водным объектам
По классификации А.М. Комлева и Е.А. Черных (1984) водотоки басс. р. Амур делятся на 4 категории, и отдельно мы выделили пятую категорию для водоёмов. Всего в водных объектах басс. Нижнего Амура обнаружено 207 видов и форм хирономид, относящихся к 5 подсемействам. По числу таксонов среди выявленных подсемейств лидируют представители подсемейства Orthocladiinae (рис. 3). По типам ареалов во всех водных объектах доминируют голаркты (табл. 3).
Таблица 3
Частота встречаемости хирономид по типам ареалов в водных объектах басс. Нижнего Амура, %
| Типы ареалов | Средние водотоки | Малые водотоки | Очень малые водотоки | Самые малые водотоки | Водоёмы |
| ВМО | 18 | 17 | 17 | 16 | 10 |
| ВПМ | 9 | 6 | 13 | 16 | 6 |
| ГОЛ | 55 | 63 | 54 | 51 | 68 |
| ПАЕ | 1 | 9 | 9 | 10 | 3 |
| ПТП | 13 | 4 | 6 | 6 | 10 |
| ПТТ | 4 | 1 | 1 | 1 | 3 |
Обозначения как в табл. 2..
11
За последние несколько лет различными авторами для басс. Нижнего Амура было описано 23 новых для науки вида из подсемейств Prodiamesinae, Orthocladiinae и Chironominae (Макарченко и др., 2008, 2009), в том числе 2 вида (Monodiamesa kamora Makarchenko et Yavorskaya, 2008 и Hydrobaenus sigaensis Makarchenko, Makarchenko et Yavorskaya, 2009) были описаны мной в соавторстве с руководителем и 10 видов (Boreosmittia khehtsyrika Makarchenko et Makarchenko, 2009, Bryophaenocladius pseudosetosus Makarchenko et Makarchenko, 2009, B. setosus Makarchenko et Makarchenko, 2009, B. nadezhdae Makarchenko et Makarchenko, 2009, Cricotopus (Isocladius) amurensis Makarchenko et Makarchenko, 2007, Propsilocerus amurensis Makarchenko et Makarchenko, 2009, Dicrotendipes orientalis Zorina, 2006, Hydrobaenus maladistinctus Makarchenko et Makarchenko, 2009, H. parvacaudatus Makarchenko et Makarchenko, 2009, Chaetocladius magnalobus Makarchenko et Makarchenko, 2009) – по моим материалам.
5.2. Аннотированный список видов
В аннотированном списке приведена характеристика для 330 видов, которая включает данные по первоописанию, географическому распространению, распределению по водотокам и водоемам, изученности стадий метаморфоза, срокам лёта имаго, биотопической приуроченности личинок, а также их нахождению в питании рыб. Анализ этого списка показал, что для 177 видов известны имаго самцы, куколки и личинки, для 27 – имаго самцы и куколки, для 98 – только имаго самцы, для остальных видов – только личинки. Для 302 видов указаны сроки лёта имаго, 180 – биотопическая приуроченность личинок и их нахождение в желудках рыб.
5.3. Систематика подсемейства Orthocladiinae
Подсемейство Orthocladiinae установлено Эдвардсом в 1929 г. (Edwards, 1929). Его представители встречаются во всех зоогеографических областях, включая Антарктику. В ранних работах по систематике подсемейство часто делили на три – Orthocladiinae, Clunioninae и Corynoneurinae, а первое – на две трибы: Orthocladiini и Metriocnemini. В настоящее время это разделение не принято (Макарченко, Макарченко, 2006).
Для имаго Orthocladiinae характерно отсутствие в жилковании крыла поперечной жилки MCu. Крылья обычно хорошо развиты и могут иметь макротрихии. Жилки R1, R2+3, R4+5 в основном отчетливые и отделены друг от друга. Антенна обычно с 13 флагелломерами. Первый членик передней лапки короче передней голени. Пульвиллы часто отсутствуют, но могут быть хорошо или слабо развиты. Тергит IX может быть с анальным отростком разной формы, длины, прозрачности или без него. Вирга в основном имеется. Гоностиль обычно с терминальным шипом и подвижно сочленён с гонококситом и загнут внутрь. Гонококсит обычно с нижней волзеллой, реже с верхней волзеллой и очень редко со срединной волзеллой.
Куколки Orthocladiinae характеризуются закрытыми стигмами на голых или покрытых шипиками торакальных рогах, если таковые имеются, лопасти
12
анального сегмента могут быть с бахромой плавательных щетинок или без них, с макрощетинками, анально-латеральные углы сегмента VIII без шипов и гребней, сегмент VIII без каудальных лопастей.
Личинки Orthocladiinae имеют следующие характерные признаки: невтяжные антенны, некольчатый 3-й членик антенны, отсутствующие или слабо развитые вентроментальные пластинки, глазные пятна одной стороны распо-
ложенные один за другим или слиты в одно пятно, лабрум без поперечного ряда чешуек, мандибула с внутренней щетинкой, расщеплённой обычно на несколько ветвей.
В Голарктике насчитывается более 700 видов ортокладиин из 90 родов (Sther et al., 2000). Подсемейство наиболее богато видами и его представители доминируют во многих донных пресноводных сообществах, но оно очень плохо изучено в России.
По данным Е.А. Макарченко и др. (2005) фауна хирономид российского Дальнего Востока подсемейства Orthocladiinae представлена 268 видами из 59 родов. Из них в басс. р. Амур пока нами не обнаружены представители 12 родов: Heleniella, Mesocricotopus, Sasacricotopus, Doncricotopus, Antillocladius, Parachaetocladius, Paratrissocladius, Oliveridia, Ninelia, Zalutschia, Tsudayusurika, Compterosmittia.
По оригинальным и литературным данным для басс. Нижнего Амура в настоящее время нами зарегистрировано 173 вида ортокладиин из 50 родов. Впервые для этого района обнаружены 4 рода: Boreosmittia, Paralimnophyes, Parasmittia, Trichosmittia, и 20 видов. Так как было нецелесообразно делать определительную таблицу для родов и видов только одной части басс. р. Амур, мы составили ключи по имаго самцам для идентификации родов и 207 видов, обнаруженных в бассейне всей реки.
ГЛАВА 6. ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ И РАСПРЕДЕЛЕНИЕ ПО ВОДНЫМ ОБЪЕКТАМ ХИРОНОМИД БАССЕЙНА НИЖНЕГО АМУРА
6.1. Географическое распространение
Изучено географическое распространение и выявлено 7 типов ареалов для 302 видов хирономид басс. Нижнего Амура (табл. 2).
Наиболее разнообразной является группа палеарктов, включающая 173 вида – это 57,3% от всей фауны и группа голарктов, состоящая из 129 видов, что составляет 42,7%. Среди видов с палеарктическим типом распространения преобладают восточно-палеарктические материково-островные (55 видов (или 31,8% от всех палеарктов), восточно-палеарктические материковые (44 вида или 25,4%) и трансевразиатские подизъюнктивные виды (36 видов или 20,8%). Меньше обнаружено представителей с палеарктическим амфиевразиатским ареалом (29 видов или 16,8%). Доля видов с другими типами распространения незначительна – 8 видов или 4,6% палеарктических темперантных и один вид аркто-альпийский (рис. 2).
13
Таблица 2
Распределение видов хирономид басс. Нижнего Амура по типам ареалов
| Тип ареала | Всего видов | % | Подсемейства | |||||
| 1 | 2 | 3 | 4 | 5 | 6 | |||
| ВМО | 55 | 18,2 | 24 | 4 | 25 | – | – | 2 |
| ВПМ | 44 | 14,5 | 7 | 1 | 34 | – | 2 | – |
| ГОЛ | 129 | 42,7 | 38 | 9 | 74 | 2 | 3 | 3 |
| ПАА | 1 | 0,3 | – | – | 1 | – | – | – |
| ПАЕ | 29 | 9,6 | – | 1 | 23 | – | 1 | 4 |
| ПТП | 36 | 12 | 29 | 1 | 4 | – | – | 2 |
| ПТТ | 8 | 2,7 | 5 | – | 2 | – | – | 1 |
| Всего | 302 | 100 | 103 | 16 | 163 | 2 | 6 | 12 |
Примечание: типы ареалов: ГОЛ – голарктический, ПАЕ – палеарктический амфиевразиатский, ПТП – палеарктический трансевразиатский полидизъюнктивный; ПТТ – палеарктический трансевразиатский темперантный, ПАА – палеарктический аркто-альпийский, ВМО – восточно-палеарктический материково-островной, ВПМ – восточно-палеарктический материковый. Подсемейства: 1 – Chironominae, 2 – Diamesinae, 3 –Orthocladiinae; 4 – Podonominae, 5 – Prodiamesinae, 6 – Tanypodinae.
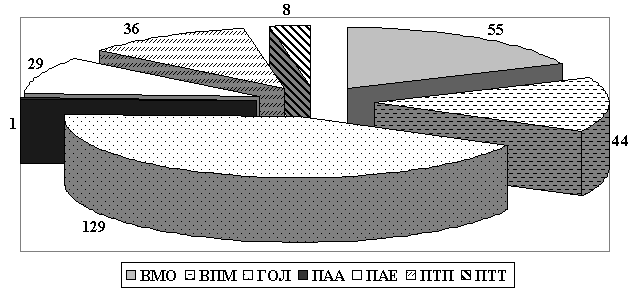
Рис. 2. Распределение видов хирономид басс. Нижнего Амура по типам ареалов. Обозначения как в табл. 2.
Впервые для Палеарктики из басс. Нижнего Амура нами указано 2 вида – Hydrobaenus laticaudus Sther, 1976 и Bryophaenocladius pleuralis (Malloch, 1915). Новыми для фауны России оказалось 7 видов – Trichosmittia hikosana Yamamoto, 1998, Krenosmittia toyamaquerea (Sasa, 1996), B. pleuralis (Malloch, 1915), Paralimnophyes longiseta (Thienemann, 1921), Pseudosmittia angusta (Edwards, 1929), H. laticaudus Sther, 1976, H. biwaquartus (Sasa et Kawai, 1987), причём род Trichosmittia до нашей находки в басс. Нижнего Амура не был известен за пределами типового местообитания на о-ве Кюсю в Японии. Впервые для фауны Дальнего Востока России зарегистрировано 7 видов – T. hikosana Yamamoto, 1998, K. toyamaquerea (Sasa, 1996), B. pleuralis (Malloch, 1915), P. longiseta (Thienemann, 1921), P. angusta (Edwards, 1929), H. laticaudus Sther, 1976, H. biwaquartus (Sasa et Kawai, 1987).