

Роль грызунов в циркуляции хантавирусов в лесных экосистемах южного приморья
На правах рукописи
СимоновА Татьяна Леонидовна
Роль грызунов в циркуляции хантавирусов
в лесных экосистемах Южного Приморья
03.00.16 - экология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Владивосток - 2008
Работа выполнена в лаборатории биогеографии и экологии
Тихоокеанского института географии ДВО РАН
Научный руководитель: доктор географических наук, ст.н.с.
Симонов Сергей Будимирович
Официальные оппоненты: доктор биологических наук, профессор
Нестеренко Владимир Алексеевич
кандидат биологических наук
Бурковский Олег Александрович
Ведущее учреждение: Институт водных и экологических проблем
ДВО РАН, г. Хабаровск
Защита состоится 4 декабря 2008 г. в 10 часов на заседании
диссертационного совета Д 005.003.03 при Биолого-почвенном институте ДВО РАН
по адресу: 690022, Владивосток, проспект 100 лет Владивостоку, 159.
Факс: (42342) 310-193
С диссертацией можно ознакомиться в Центральной научной библиотеке ДВО РАН
Автореферат разослан "_____" ______________ 2008 г.
Ученый секретарь
диссертационного совета,
кандидат биологических наук Е.М. Саенко
Общая характеристика работы
Актуальность темы. Хантавирусы вызывают заболевание людей геморрагической лихорадкой с почечным синдромом (ГЛПС) - одной из самых распространённых зоонозных природноочаговых инфекций, которая по показателям заболеваемости лидирует в России. Большинство заражений людей происходит в Европейской части нашей страны (97 %), наиболее изученной в отношении этой болезни, и только 3 % - в Азиатской, главным образом, на Дальнем Востоке (Ткаченко, 2000). Клиническое течение ГЛПС в Европейской части более благоприятное, чем в дальневосточных очагах. В последних чаще регистрируются тяжёлые либо среднетяжёлые формы ГЛПС (Яшина и др., 2003).
Приморский край относится к активному природному очагу хантавирусной инфекции со среднегодовыми показателями заболеваемости до 5,6 на 100 тысяч населения (Слонова и др., 2002). Установлено, что особенности эпидемиологического проявления хантавирусной инфекции зависят от доминирования на очаговой территории определённых видов грызунов - носителей инфекции. В Приморье выявлена циркуляция 5 генотипов хантавируса, для каждого из которых определён природный хозяин. С эпидемической точки зрения в природных биотопах наибольшую значимость имеют полевая (Apodemus agrarius Pallas,1771) и восточноазиатская мыши (Apodemus peninsulae Thomas,1906 ), носители патогенных генотипов Hantaan и Amur, роль которых в инфицировании людей доказана (Слонова и др., 2002).
Главные достижения в природноочаговых исследованиях на Дальнем Востоке связаны прежде всего с изучением полевой мыши, обитающей в лугово-полевых ландшафтах равнин. В горно-лесных территориях, занимающих почти 2/3 площади Приморского края, первостепенное значение как резервуара хантавирусов принадлежит восточноазиатской мыши, роль которой в циркуляции инфекции детально не изучена, что объясняется сравнительно недавней идентификацией вида как основного носителя одного из патогенных возбудителей (Yashina et al., 2000).
Цель и задачи исследования. Основная цель работы - выявление роли динамики населения мышевидных грызунов в циркуляции хантавирусов в горно-лесных экосистемах Южного Приморья.
В связи с этим необходимо было решить следующие задачи:
1. Показать роль фоновых видов грызунов лесных экосистем в возникновении эпизоотий хантавирусной инфекции.
2. Проанализировать активность эпизоотического процесса в зависимости от фаз численности популяционного цикла грызунов - основных носителей хантавирусной инфекции.
3. Выявить эпизоотологические особенности горно-лесного очага хантавирусной инфекции на юге Приморского края.
4. Проследить инфицированность хантавирусом грызунов хребта Ливадийского в связи с особенностями их высотно-поясной дифференциации.
5. Выявить типы пространственной структуры популяций мышевидных грызунов лесных экосистем Сихотэ-Алиня.
6. Показать возможность существования мест устойчивого сохранения возбудителя хантавирусной инфекции и проследить их взаимосвязь с пространственной неоднородностью популяций грызунов.
Защищаемые положения.
1. Активность эпизоотического процесса хантавирусной инфекции в лесных экосистемах Южного Приморья обусловлена пространственно-временной динамикой грызунов-вирусоносителей.
2. Инфицированность грызунов хантавирусом в горно-лесных ландшафтах является отражением высотно-поясного распределения видов-вирусоносителей.
3. Тип пространственной структуры популяций лесных видов грызунов неустойчив и обусловлен уровнем их численности.
4. Места устойчивого сохранения возбудителя хантавирусной инфекции приурочены к участкам постоянной локализации грызунов.
Научная новизна работы. Показана роль структуры сообществ мышевидных грызунов в потенциальной опасности заболевания людей ГЛПС. Впервые представлены данные по высотно-поясному распределению инфицированных хантавирусом мышевидных грызунов на Южном Сихотэ-Алине, обозначены высоты, на которых отмечена максимальная численность инфицированных зверьков. На горно-лесной очаговой территории выявлены типы местообитаний, в которых инфицированные хантавирусом мышевидные грызуны отлавливались с наибольшей численностью. Прослежена структура сообществ грызунов лесных экосистем на протяжении полного популяционного цикла их численности и показано, что потенциальная опасность заражения людей ГЛПС возрастает в годы доминирования в населении восточноазиатской мыши, а также в годы её высокой численности. Показана синхронность циклов популяционной динамики восточноазиатской мыши, красно-серой полёвки и возбудителя хантавирусной инфекции. Рассмотрены характерные особенности пространственной структуры популяций лесных видов грызунов в различные фазы их популяционной численности. Показано, что зимний период определяет дальнейшее развитие эпизоотического процесса на очаговой горно-лесной территории. Показано наличие ядер очаговости хантавирусной инфекции на энзоотичной территории юга Приморского края
Практическая значимость работы. Обозначенные критерии оценки активности эпизоотического процесса на разных фазах популяционного цикла грызунов лесного комплекса могут быть использованы при оценке потенциальной опасности территории по заболеваемости людей геморрагической лихорадкой с почечным синдромом. Материалы по структуре сообществ мышевидных грызунов, их численности, структуре очагов хантавирусной инфекции, высотно-поясному распределении инфицированных зверьков, пространственной структуре популяций видов - вирусоносителей могут служить основой при оценке эпидемической опасности территории и планировании профилактических мероприятий.
Апробация работы. Материалы диссертационной работы были представлены на VI научной конференции молодых географов Сибири и Дальнего Востока (Иркутск, 1975); II и III региональных научных конференциях "Динамика численности мелких млекопитающих на Дальнем Востоке СССР и их роль в экосистемах" (Владивосток, 1986; Хабаровск, 1988); XIX Тихоокеанском конгрессе (Сидней, 1999); 29 международном географическом конгрессе (Сеул, 2000); международной конференции, посвящённой классификации и динамике лесов Дальнего Востока (Владивосток, 2001); 3-ей региональной научно-практической конференции к Всемирным дням Воды и Метеорологии (Владивосток, 2002); научно-практической конференции с международным участием, посвящённой 70-летию изучения проблемы геморрагической лихорадки с почечным синдромом на Дальнем Востоке России (Владивосток, 2003); XII научном совещании географов Сибири и Дальнего Востока (Владивосток, 2004); 6-ой и 7-ой международных конференциях по геморрагической лихорадке с почечным синдромом, хантавирусному пульмональному синдрому и хантавирусам (Сеул, 2004; Буэнос-Айрес, 2007).
Публикации. По теме диссертации опубликовано 25 работ.
Структура и объём работы. Диссертация состоит из введения, 5 глав, выводов, списка литературы и приложения. Оформлена в виде рукописи, содержащей 134 страницы печатного текста, включает 5 таблиц, 28 рисунков. Список литературы состоит из 211 источников, в том числе 32 на иностранных языках.
Благодарности. Автор выражает искреннюю благодарность Р.А. Слоновой, П.С. Симонову, Г.Г. Компанец и Т.В. Кушнарёвой за помощь, оказанную в сборе и обработке материала, особенную признательность - моему руководителю С.Б. Симонову.
Глава 1. ФИЗИКО-ГЕОГРАФИЧЕСКИЕ ОСОБЕННОСТИ
ПРИМОРСКОГО КРАЯ
В главе приведены краткие сведения о физико-географических особенностях района исследований. Показано разнообразие орографических, гидрологических, климатических и геоботанических показателей на рассматриваемой территории (Ливеровский, Колесников, 1949; Куренцова, 1968; Ларинцева, 1990; Ивашинников, 1999 и др.). Отмечено, что в результате антропогенной деятельности в настоящее время меняются условия существования многих животных, что в свою очередь приводит к изменению условий формирования и функционирования очагов хантавирусной инфекции.
Глава 2. МЕТОДИКА И МАТЕРИАЛ
Полевые работы по изучению пространственно-временной динамики населения мышевидных грызунов и её связи с характером циркуляции хантавирусов проводились в горно-лесных ландшафтах Южного Приморья на северном макросклоне хребта Ливадийского с 1998 по 2006 гг. (рис. 1).
В соответствии с основной целью работы для оценки высотно-поясного распределения грызунов и активности эпизоотического процесса ежегодно с 1999 по 2003 гг. по сезонам, в строго ограниченные сроки (весной, летом и осенью), в диапазоне высот от 100 до 1100 м над ур. моря на различных высотных уровнях с шагом в 100 м по абсолютной высоте на стационарных линиях осуществлялись учёты мышевидных грызунов по стандартной методике (Новиков, 1953; Кучерук, 1963). На каждом высотном уровне на одну ночь выставлялось не менее 100 плашек Геро, а в годы глубоких депрессий численности - до 200. Для каждой линии ловушек делалось стандартное геоботаническое описание (Сукачёв, 1972). Дополнительные наблюдения на хребте Ливадийском были проведены в год высокой популяционной численности грызунов в июле 2005 г.
Для понимания более тонких внутрипопуляционных механизмов, обусловливающих характер эпизоотического процесса, выявления характера динамики пространственной структуры популяций фоновых видов в 2003 - 2006 гг. осуществлялись наблюдения на модельном стационарном участке, расположенном в диапазоне высот от 200 до 400 м над ур. моря и вытянутом вдоль долин ручьёв Прямой Ключ и Тёмный приблизительно на 5 км при ширине до 1 км. Учёты на площадке проводились ежегодно по сезонам (весна, лето, осень), в каждый тур таких учётов выставлялось на одну ночь 60 - 62 линии по 25 ловушек в каждой.
Всего за годы исследований отработано 38750 ловушко-ночей и отловлено 4608 зверьков 5 видов: восточноазиатская мышь, полевая мышь, красно-серая полёвка (Myodes rufocanus Sundervall, 1846), красная полёвка (M. rutilus Pallas,1778) и большая, или дальневосточная полёвка (Microtus fortis Buchner,1889) (названия видов приведены по Каталогу млекопитающих СССР (1981) с исправлениями И.Я. Павлинова, 2006).
У каждой отловленной особи определяли пол, участие в размножении в момент исследования и возраст (Варшавский, Крылова, 1948; Кошкина, 1959, Тупикова и др., 1957; 1979). Для изучения характера эпизоотического процесса хантавирусной инфекции проводились специальные работы по отбору биологических проб и крови у отловленных грызунов. Всего на наличие хантавирусов обследовано 2787 особей, от которых взято 18509 проб внутренних
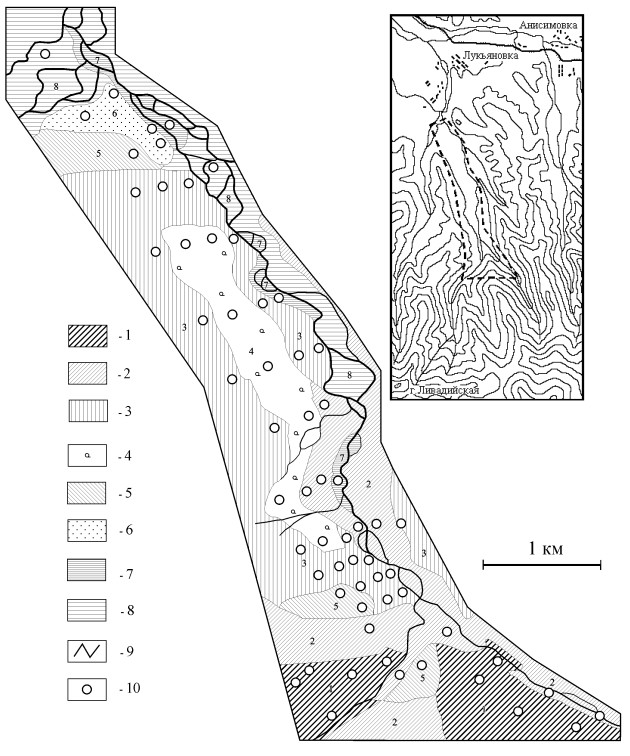
Рис. 1. Схема района работ и стационарной учётной площадки
Типы местообитаний: 1 - темнохвойные леса; 2 - хвойно-широколиственные леса; 3 - кедрово-широколиственные леса, местами со старыми выборочными рубками; 4 - кедрово-широколиственные редины по старым рубкам в долинах; 5 - мелколиственные леса с примесью широколиственных; 6 - травяно-кустарниковые сообщества; 7 - чозениево-тополёвые долинные леса; 8 - полидоминантные широколиственные долинные леса.
9 - реки; 10 - места постановки ловушко-линий на площадке.
Пунктиром отмечено месторасположение стационарной площадки.
органов и крови. К особям, участвующим в эпизоотическом процессе, отнесены те, в пробах крови и внутренних органов которых в результате иммунологических методов исследования был обнаружен антиген хантавируса либо специфические антитела. Поиск антигена вируса проводился методом иммуноферментного анализа (ИФА), используя "Хантагност" производства Института полиомиелита и вирусных энцефалитов им. М.Н. Чумакова РАМН. Антитела к хантавирусам определяли непрямым методом флюоресцирующих антител (НМФА) с использованием культуральных антигенов хантавирусов. Все вирусологические исследования выполнены сотрудниками лаборатории ГЛПС НИИЭМ СО РАМН. Инфицированность грызунов приведена в особях-вирусоносителях на 100 л-н (Бернштейн и др., 1989).
Оценка динамики пространственно-временного распределения сообществ грызунов и специфики функционирования очага хантавирусной инфекции проводилась с помощью картографических методов (Тупикова, 1969; Тупикова, Комарова, 1979). В качестве картографической основы использовались топографические карты территории масштаба 1:25000 и 1:50000, лесотаксационные планы Анисимовского лесничества и полевая геоботаническая съёмка автора.
Степень доминирования видов грызунов определялась по доле их участия в населении: более 50,0 % населения - абсолютный доминант; 30,1 - 50,0 % - доминант; 10,1 - 30,0 % - содоминант; 2,1 - 10,0 % - второстепенный член; не более 2,0 % - третьестепенный.
Глава 3. ДИНАМИКА НАСЕЛЕНИЯ ГРЫЗУНОВ И ЕЁ РОЛЬ
В ЦИРКУЛЯЦИИ ХАНТАВИРУСОВ
Анализ пространственно-временного распределения сообществ грызунов в течение двух полных популяционных циклов численности показал, что в зависимости от сезона и фазы популяционной численности участие видов в населении значительно колеблется. В целом средняя многолетняя структура мышевидных грызунов характеризуется абсолютным доминированием восточноазиатской мыши - 69,6%, содоминированием красно-серой полёвки - 23,7%. Доля полевой мыши составляет 3,7%, красной полёвки - 2,3% населения. Большая полёвка отлавливалась в единичных экземплярах.
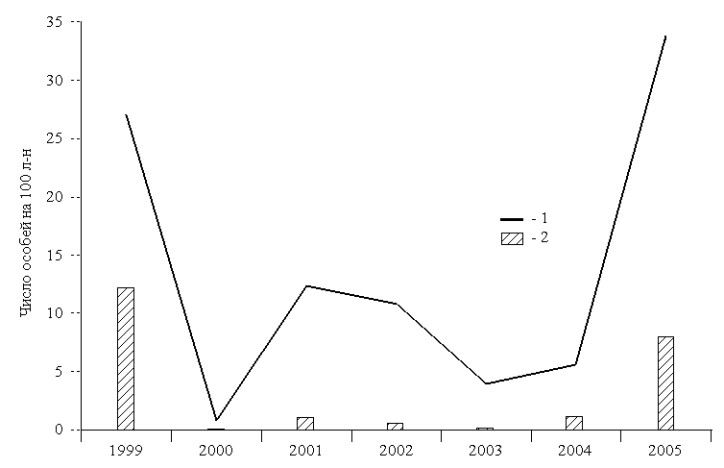
Инфицированность грызунов хантавирусом непосредственно связана с популяционными колебаниями их численности (рис. 2). Так, в годы "депрессий" (2000 и 2003 гг.) наблю-
Рис. 2. Сопряжённость численности грызунов и инфицированности их хантавирусом.
1 - все особи; 2 - инфицированные особи.
далось затухание эпизоотического процесса, в годы падения либо подъёма численности (2002 и 2004 гг.) отмечалась его активизация, а в годы "пиков" (1999 и 2005 гг.) выявлена максимальная инфицированность грызунов хантавирусом, которая прежде всего обусловлена инфицированностью восточноазиатской мыши - основного носителя патогенного генотипа Amur. При благоприятных условиях грызуны способны успешно размножаться в зимний период, активно расселяться по территории и резко наращивать численность к весне. Постоянные контакты между грызунами в этот период способствуют активной циркуляции хантавируса в их населении.
Глава 4. ВЛИЯНИЕ ВЫСОТНОЙ ПОЯСНОСТИ НА РАСПРЕДЕЛЕНИЕ И
ИНФИЦИРОВАННОСТЬ ГРЫЗУНОВ ХАНТАВИРУСОМ
В главе рассмотрены особенности инфицированности грызунов, обусловленные их высотно-поясной дифференциацией на северном макросклоне хр. Ливадийского (рис. 3).
Восточноазиатская мышь доминировала во всех исследованных типах местообитаний основных поясов растительности (59,3-77,0%). Её участие в населении постепенно снижается с ростом абсолютной высоты, при этом наблюдается увеличение доли лесных полёвок. Доля красно-серой полёвки остаётся относительно стабильной (на уровне содоминанта) почти на всех высотных уровнях. Красная полёвка принимает наибольшее участие в сообществах грызунов верхних высотных поясов, где преобладают тёмнохвойные породы деревьев.
Полевая мышь, предпочитая лугово-полевые местообитания, при наличии пригодных биотопов может встречаться в широком диапазоне высот, вплоть до 1100 м над уровнем моря. Общая численность грызунов при подъёме в горы постепенно падает от 14,5 до 6,5 ос./100 л-н. Их инфицированность при этом колеблется. Минимальная (0,3 ос./100 л-н) зафиксирована в поясе кедрово-широколиственных лесов на высоте 500-700 м. Максимальная (2,5 ос./100 л-н) - в этом же поясе на высоте 300-500 м над уровнем моря. Выше 700 м общая инфицированность зверьков колеблется от 0,9 до 1,2 ос./100 л-н.
Следует подчеркнуть, что максимальная инфицированность грызунов хантавирусом приходится на местообитания, расположенные до 500 м над уровнем моря, которые являются наиболее посещаемыми туристами, отдыхающими и сборщиками дикоросов.
Глава 5. ПРОСТРАНСТВЕННАЯ СТРУКТУРА ПОПУЛЯЦИЙ
ГРЫЗУНОВ - ВИРУСОНОСИТЕЛЕЙ
5.1. Население грызунов стационарного участка
Комплекс работ по оценке характера эпизоотического процесса и динамики пространственной структуры видов мышевидных грызунов проводился на стационарной площадке, заложенной, как указывалось выше, в диапазоне высот от 200 до 400 м над ур. моря, где была зафиксирована максимальная инфицированность грызунов.
На исследуемой территории выделено 8 основных типов местообитаний, каждый из которых характеризуется определённым населением грызунов и характером эпизоотического процесса. Структура территориальных группировок в отдельных типах местообитаний подвержена периодическим изменениям: перестройки сообществ грызунов происходят как во времени, так и в пространстве в зависимости от экологической ситуации, сложившейся на территории их обитания. Активно передвигаясь, зверьки находят наиболее благоприятные места, которые непостоянны, а меняются в зависимости от условий существования видов в конкретное время. В большинстве типах местообитаний достаточно чётко прослеживаются
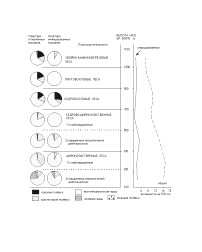
Рис. 3. Высотная поясность и инфицированность населения грызунов.
сезонные аспекты населения, характеризующиеся сменой доминирующих видов вне зависимости от фазы популяционной численности последних. Инфицированность грызунов во всех типах местообитаний колеблется в зависимости от фазы популяционного цикла и сезонов года, при этом потенциальная опасность заражения людей хантавирусной инфекцией также меняется.
5.2. Картографирование как метод изучения пространственного распределения
грызунов
Изучение пространственной структуры популяций грызунов позволяет понять механизмы развития эпизоотического процесса хантавирусной инфекции на энзоотичной территории. При этом большое значение следует придавать анализу ключевых участков с помощью приёмов крупномасштабного картографирования, так как сопоставление картосхем, полученных этим путём, позволяет проследить пространственную динамику распределения мышевидных грызунов и дать количественную оценку движущих сил эпизоотического процесса (Кучерук и др., 1965; Ковалевский, Коренберг и др., 1974). Основными способами картографирования подобных явлений можно считать картограмму и значковый (Тупикова, Комарова, 1979), которые отражают разные стороны изучаемого процесса.
Первый, картограмма, показывает осреднённые значения численности вида в исследованных природных растительных выделах. При этом создаётся впечатление, что особи изучаемого вида распределены в них равномерно. Второй, значковый способ, отражает конкретные места локализации животных в момент учётов, тем самым восполняя пробелы картограммы. В работе были применены оба способа, однако для целей наших исследований значковый способ оказался более информативным.
В результате анализа динамики численности создана серия картосхем, отражающая особенности пространственного распределения видов-вирусоносителей на ключевом участке горно-лесного ландшафта (рис. 4). Сделаны выводы о возможности применения различных способов картографирования при выявлении роли пространственно-временной структуры грызунов в поддержании очагов хантавирусной инфекции.
В последующих разделах главы рассмотрена особенности пространственной структуры отдельных видов на разных фазах их популяционной численности.
5.3. Пространственная структура популяции восточноазиатской
мыши и её инфицированность
Восточноазиатская мышь - носитель патогенного хантавируса Amur, является наиболее многочисленным и широко распространённым видом грызунов лесных ландшафтов.
5.3.1. Фаза популяционной депрессии численности.
2003 год характеризовался низкой численностью восточноазиатской мыши. Дана сезонная динамика распределения данного вида и вирусоносителей по отдельным типам местообитаний. Если весной встречи зверьков были очень редки, летом- фиксировались во всех 8 типах местообитаний, при концентрации в хвойно-широколиственных лесах, то осенью, перераспределившись по территории, максимальное их количество отмечалось в темнохвойных лесах. Инфицированные особи восточноазиатской мыши в 2003 г. отлавливались во все сезоны в местах их концентрации.
5.3.2. Фаза популяционного подъёма численности.
В 2004 г. на фоне незначительного роста численности восточноазиатской мыши по сравнению с 2003 годом существенно увеличилось число отловленных инфицированных особей в летний и осенний периоды, что совпало с расселением вида по территории. К осени наметилась тенденция к концентрации зверьков в местах предстоящей зимовки.
5.3.3. Фаза популяционного максимума численности.
Зима 2004-2005 гг. оказалась благоприятной для мышевидных грызунов. Невысокий снежный покров, отсутствие резких перепадов температур и наличие высококалорийного корма ( урожай кедровых орехов) способствовали увеличению численности и активному перемещению восточноазиатской мыши уже в зимний период. К весне 2005 г. характер её распределение по территории стал диффузным и оставался таким на протяжении всего года, слабо меняясь по сезонам. Зверьки встречались во всех типах местообитаний, концентрируясь в темнохвойных и широколиственных лесах (более 20 ос./100 л-н). Высокая численность вида обусловила активизацию эпизоотического процесса уже к весне 2005 г. Максимум инфици-
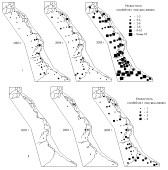
Рис. 4. Динамика численности (I) и инфицированности (II)
восточноазиатской мыши в летний период.
рованных особей пришёлся на лето, при этом их распределение было также диффузным.
Осенью вирусоносители не отмечались только в широколиственных лесах и разнотравно-кустарниковых сообществах.
5.3.4. Фаза популяционного минимума численности.
Наблюдения весной 2006 года показали, что полный цикл численности восточноазиатской мыши завершился. Данный вид, как и в 2003 году имел типичное для "депрессии" пятнистое распределение по территории, при этом инфицированные особи отлавливались единично.
5.4. Пространственная структура популяции красно-серой полёвки
и её инфицированность хантавирусом
Красно-серая полёвка - носитель хантавируса Puumala, является одним из доминирующих видов в сообществах мышевидных грызунов горно-лесных ландшафтов Приморья.
5.4.1. Фаза популяционной "депрессии" численности.
2003 год характеризовался низкой численностью красно-серой полёвки. Пространственная структура данного вида была однотипной на протяжении всего года, единичные инфицированные особи встречались в лесах с присутствием хвойных пород деревьев.
5.4.2. Фаза популяционного подъёма численности.
В 2004 г. наметился незначительный подъём численности красно-серой полёвки, достигший сезонного максимума в осенний период. В это же время отлавливалось больше всего инфицированных особей (1,0 - 1,9 ос./100 л-н), концентрировавшихся в хвойно-широколиственных, чозениево-тополёвых лесах и кедрово-широколиственных рединах.
5.4.3. Фаза популяционного максимума численности.
Для красно-серой полёвки, как и для восточноазиатской мыши, зимовка 2004-2005 гг. была очень благоприятной и оказала решающее влияние на изменение структуры популяции к весне 2005 г. Если в весенние периоды 2003 и 2004 гг. наблюдался пятнистый характер распределения вида по территории, то в соответствующий период 2005 г. он становится диффузным. Полёвки встречались во всех типах местообитаний с численностью, нередко превышающей 8,0 ос/100 л-н. В это же время был отмечен сезонный пик их инфицированности - до 6,0 ос./100 л-н. Летне-осенний рост обилия данного вида не привел к активизации эпизоотического процесса.
5.4.4. Фаза популяционной "депрессии" численности.
Весной 2006 г. популяционный цикл численности красно-серой полёвки завершился: её распределение по территории было таким же, как в год "депрессии" 2003 г. Количество инфицированных особей было низким (менее 1,0 ос./ 100 л-н).
5.5. Пространственная структура популяции полевой мыши и её
инфицированность хантавирусом
Полевая мышь - носитель патогенного хантавируса генотипа Hantaan периодически заходит на горно-лесную территорию при вспышках численности. Однако наличие пригодных для неё местообитаний и возможность закрепления её на данной территории вызывает необходимость рассмотрения распределения данного вида на площадке.
В фазе "депрессии" численности (2003 г.) полевая мышь на исследуемой территории не встречалась
В фазе популяционного подъёма численности (2004 г) она отлавливалась с небольшой численностью в некоторых типах местообитаний. С уловистостью от 4,0 до 7,9 ос./100 л-н данный вид фиксировался летом в нарушенных долинных широколиственных лесах и разнотравно-кустарниковых сообществах. Только в последних в этот же период года встречались инфицированные особи.
В 2005 году, когда отмечался "пик" численности грызунов, полевая мышь отлавливалась во все сезоны года в наиболее благоприятных для неё наиболее нарушенных человеком местообитаниях (1,0 - 7,9 ос./100 л-н). Инфицированные хантавирусом зверьки также отлавливались во все сезоны с численностью не превышающей 1,9 ос./100 л-н.
5.6. О типизации пространственной структуры грызунов лесных местообитаний
Особенности локализации мест эпизоотических проявлений многих природно-очаговых болезней зависит от структуры популяций грызунов-вирусоносителей в момент исследования. Поэтому работы, показывающие динамику размещения отдельных видов - носителей хантавируса приобретают особую значимость при изучении очагов хантавирусной инфекции.
М.Е.Косой (1987) указывал, что пространственная структура популяции восточноазиатской мыши в Приморье относится к пульсирующему типу. Однако, анализ распределения данного вида, проведённый с учётом сезонной динамики, не позволяет нам согласиться с этим, т.к. отсутствует ряд признаков, характеризующих этот тип. Считаем, что популяционная структура восточноазиатской мыши не является устойчивой и на разных фазах популяционного цикла может изменяться по сезонно-циклическому типу (при низкой численности) либо эквальному (годы максимума) (в понятии В.Е. Флинта, 1977).
Распределение красно-серой полёвки на исследуемой очаговой территории также не является устойчивым и меняется в зависимости от уровня численности животных. В годы невысокой уловистости зверьки распределены пятнами, которые в зависимости от сезона могут несколько увеличивать свои размеры. В год максимума их размещение носит диффузный характер. Распределение красно-серой полёвки в лесных экосистемах Южного Приморья не вкладывается ни в один из описанных ранее типов, занимая промежуточное положение между эквальным и циклическим.
Размещение полевой мыши на горно-лесной территории носит случайный характер: вид не образует устойчивых поселений, хотя отдельные особи способны проникать в лесные биотопы достаточно далеко, вплоть до высоты 1100 м над ур. моря. Горно-лесные ландшафты являются для полевой мыши своеобразной зоной выселения, куда зверьки откочёвывают при их перенаселении в оптимальных лугово-полевых биотопах.
5.7. Локализация грызунов и устойчивость очаговых проявлений
Опыт медико-зоологических исследований показал, что в силу ряда обстоятельств, возбудитель болезни сохраняется в течение длительного времени не на всей территории очага, а лишь в определённых его точках, стратегически важных узлах (Наумов, 1955; 1959), которые в настоящее время принято называть ядрами очага (Кучерук, 1972).
Территориально локализация ядер обусловлена наличием биогеоценозов (фаций), условия которых благоприятны для длительного существования возбудителя, что обычно связано с особенностями образа жизни носителей и переносчиков. В нашем случае - это доминирующие в населении грызунов восточноазиатская мышь и красно-серая полёвка.
Структура природного очага хантавирусной инфекции наиболее детально изучена нами на примере носителя патогенного генотипа Amur - восточноазиатской мыши. Опираясь на наблюдения, отражающие особенности распределения вида на стационарной площадке в течение полного популяционного цикла, нам удалось выделить места постоянного присутствия зверьков, которые можно рассматривать как элементарные поселения, или мерусы (Флинт, 1977).
Обобщив места отлова инфицированных особей по сезонам, мы рассмотрели особенности их размещения на энзоотичной территории в годы с различной численностью вида (депрессия, подъём, пик).
При очень низкой численности в 2003 г. отмечено случайное распределение зверьков данного вида: на стационарных учётных линиях инфицированные особи отлавливались не чаще 1 раза в год.
В год подъёма численности (2004) инфицированные мыши отлавливались чаще и на более обширной площади. В отдельных выявленных нами эпизоотийных пятнах вирусоносители фиксировались 2, в одном - 3 раза в течение года.
Пик численности восточноазиатской мыши в 2005 г. совпал с пиком её инфицированности хантавирусом: только на 4 учётных линиях из 62-х инфицированные особи не были встречены ни разу за год. В большинстве случаев они отлавливались 2-3 раза.
Сопоставив места отлова инфицированных мышей за все годы исследования нами было показано, что большая часть ядер инфицированности являются блуждающими, то есть их перемещение не имеет определённого вектора, а сами ядра могут смещаться в разных направлениях, периодически исчезая и появляясь вновь. Последнее, по-видимому, можно объяснить особенностями использования территории грызунами в конкретный год исследования.
Между выделенными элементарными поселениями восточноазиатской мыши и ядрами очаговости существует связь: в пяти из семи случаев их участки перекрываются полностью либо частично (рис. 5).
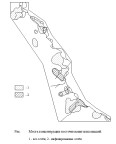
Рис. 5. Места локализации восточноазиатской мыши.
1 - все особи; 2 - инфицированные особи
В таких участках на каждой постоянной ловушко-линии мыши отлавливались не менее, чем в 7 из 9 туров наблюдений (сезонов), а инфицированные хантавирусом особи - не менее 4 раз за весь изученный нами популяционный цикл.
Участки наиболее частых встреч восточноазиатской мыши, совпавшие с эпизоотийными пятнами, имели приуроченность к долинным смешанным лесам: кедрово-широколиственным и хвойно-широколиственным.
Более низкий процент участия в структуре населения грызунов красно-серой полёвки не позволил нам провести детальный анализ частоты её встреч по отдельным годам.
Частота встреч особей данного вида на одной и той же ловушко-линии за весь период наблюдений почти в 1,5 раза ниже, чем у восточноазиатской мыши: на 5 линиях зверьки встречались 5 раз и только на одной - 6.
Выделенные ядра очаговости, по-видимому, таковыми можно считать условно, так как инфицированные полёвки встречались в них лишь по 2 раза за всё время наблюдений. Тем не менее, все эпизоотийные пятна (за исключением двух) сопряжены с местами наиболее частых отловов зверьков.
Ядра локализации красно-серых полёвок (как инфицированных, так и нет) были приурочены к долинным чозениево-тополёвым, хвойно-широколиственным, темнохвойным лесам и кедрово-широколиственным рединам.
Выводы
1. Лесной очаг хантавирусной инфекции характеризуется устойчивой среднемноголетней структурой населения грызунов с доминированием восточноазиатской мыши и содоминированием красно-серой полёвки.
2. Структура сообществ грызунов лесных экосистем определяет эпидемическую опасность территории по хантавирусной инфекции. Потенциальная опасность заражения людей ГЛПС определяется уровнем численности и степенью доминирования восточноазиатской мыши - носителя патогенного генотипа Amur.
3. Распределение грызунов в горно-лесных ландшафтах является отражением высотной поясности растительности, а их инфицированность определяется численностью видов-вирусоносителей. Наибольшая инфицированность наблюдается в нижних поясах растительности на высотах до 500 м над ур. моря - районах, активно посещаемых людьми.
4. Циклы популяционной динамики грызунов и динамики возбудителя хантавирусной инфекции синхронны.
5. Особенности сезонной динамики каждого вида грызунов обусловливают проявление сезонных максимумов их инфицированности. У восточноазиатской мыши он приходится на летний период, а у красно-серой полёвки - на весну или осень. Максимальная общая инфицированность грызунов отмечалась в коренных лесах, а минимальная - во вторичных сообществах.
6. Зимний период является определяющим в развитии эпизоотического процесса на горно-лесной очаговой территории.
7. Пространственная структура популяций лесных видов грызунов обусловлена уровнем их численности и на разных фазах популяционного цикла может изменяться по сезонно-циклическому типу (при низкой численности) либо эквальному (в годы максимума).
8. Ядра очаговости хантавирусной инфекции в значительной степени сопряжены с местами локализации грызунов, и большинство из них являются блуждающими.
Список работ, опубликованных по теме диссертации
Коллективные монографии:
1. Слонова Р.А., Симонов С.Б., Компанец Г.Г., Симонова Т.Л. и др. Хантавирусная инфекция в Приморском крае: Медико-географический атлас. Владивосток, 2007. 47 с.
2. Симонов П.С., Симонов С.Б., Симонова Т.Л. Высотно-поясная дифференциация сообществ грызунов в горных экосистемах Приморья. Владивосток: Дальнаука, 2008. 118 с.
Статьи, опубликованные в ведущих рецензируемых научных журналах:
3. Симонова Т.Л. Население мелких грызунов и ландшафтная структура территории // Вестник МГУ, сер. география. 1977, № 2. С. 105-110.
4. Яшина Л.Н., Слонова Р.А., Олейник О.В., Кузина И.И., Кушнарёва Т.В., Компанец Г.Г., Симонова Т.Л., Симонов С.Б., Нетесов В.С., Морзунов С.П. Новый генетический вариант вируса Пуумала в Приморье и его природный резервуар, красно-серая полёвка Clethrionomys rufocanus // Вопросы вирусологии. 2004. № 6. С. 34-38.
5. Слонова Р.А., Кушнарёва Т.В., Компанец Г.Г., Максема И.Г., Яшина Л.Н., Симонова Т.Л., Симонов С.Б. Геморрагическая лихорадка с почечным синдромом - особенности эпидемического процесса в очагах циркуляции разных серотипов/генотипов хантавирусов // Дальневосточный журнал инфекционной патологии. 2005, № 6. С. 47-48.
6. Симонова Т.Л., Кушнарёва Т.В., Симонов С.Б., Компанец Г.Г., Слонова Р. А. Зоографическая характеристика популяции мыши Apodemus peninsulae и её роль в поддержании хантавирусной инфекции на юге Приморского края // Журнал микробиологии, эпидемиологии и иммунобиологии. 2006, № 3, Приложение, С. 81-84.
7. Симонов С.Б., Симонова Т.Л., Слонова Р. А., Кушнарёва Т.В., Компанец Г.Г. Структура сообществ мышевидных грызунов лесного комплекса и их инфицированность хантавирусом на разных фазах популяционного цикла в Южном Приморье // Дальневосточный журнал инфекционной патологии, 2006, № 8. С. 14-20.
8. Слонова Р. А., Кушнарёва Т.В., Компанец Г.Г, Максема И.Г., Симонова Т.Л., Симонов С.Б. Хантавирусная инфекция в Приморском крае - эпидемиологическая ситуация в очагах циркуляции разных серотипов вируса // Журнал микробиологии, эпидемиологии и иммунобиологии, 2006, № 3, Приложение, С. 74-77.
Статьи, опубликованные в отечественных периодических изданиях и региональных сборниках:
9. Симонова Т.Л., Симонов С.Б. Динамика численности и структура населения мышевидных грызунов в лесных биогеосистемах Среднего Сихотэ-Алиня // Структурная организация компонентов биогеосистем (Сравнительный и количественный анализ). Владивосток: ДВНЦ АН СССР, 1988. С. 120-128.
10. Симонов С.Б., Симонова Т.Л., Симонов П.С. Основные черты пространственного распределения мышевидных грызунов хр. Ливадийского в летний период // Ландшафтно-растительная поясность Ливадийского хребта (Южное Приморье). Владивосток: Дальнаука, 2001. С. 82-92.
11. Симонова Т.Л., Симонов С.Б., Симонов П.С. Половозрастная структура и динамика размножения мышевидных грызунов лесных биогеоценозов хр. Ливадийского // Там же. С. 93-104.
12. Симонов С.Б., Слонова Р.А., Кушнарёва Т.В., Симонова Т.Л., Компанец Г.Г. Динамика популяций грызунов и её отражение в эпидемическом процессе хантавирусной инфекции // Хантавирусы и хантавирусные инфекции. Владивосток, 2003. С. 294-306.
13. Симонов П.С., Симонов С.Б., Симонова Т.Л. Особенности сезонной и многолетней динамики населения мышевидных грызунов Южного Приморья // Сборник науч. трудов молодых учёных. Географические и геоэкологические исследования на Дальнем Востоке (Владивосток, ДВГУ, 29-30 ноября 2005 г.) Владивосток: Дальнаука, 2007. Вып. 3. С. 40-48.
Работы, опубликованные в материалах всероссийских, региональных и международных научных конференций, совещаний и съездов:
14. Simonov S.B., Simonova T.L., Simonov P.S. Spatial allocation of rodents in mount Pidan region (South Primorye, Russion Far East) // 29th International Geographical congress, 14 - 18 August 2000, Seoul, Korea. Seoul, 2000. P. 75 - 76.
15. Симонов С.Б., Симонова Т.Л. Трансформация лесов Приморья и структура сообществ мышевидных грызунов // Классификация и динамика лесов Дальнего Востока // Мат-лы международ. конф. Владивосток: Дальнаука, 2001. С. 178-181.
16. Симонов С.Б., Симонова Т.Л. Картографический анализ пространственного распределения грызунов Приморья // V Дальневосточ. конф. по заповедному делу. Тез. конф. Владивосток: Дальнаука, 2001. С. 258 - 259.
17. Симонов С.Б., Симонова Т.Л. К опыту картографирования населения мышевидных грызунов Приморского края // Териофауна России и сопредельных территорий (VII съезд Териологического общества). Материалы Международного совещания 6-7 февраля 2003 г., Москва. - Москва, 2003. С. 319.
18. Симонова Т.Л., Кушнарёва Т.В., Симонов С.Б., Компанец Г.Г., Слонова Р.А. Структура населения мышевидных грызунов и их инфицированность хантавирусом на разных стадиях популяционного цикла в Южном Приморье // Хантавирусы, геморрагическая лихорадка с почечным синдромом. Науч.-практич. конференция, 24-25 сентября 2003 г. Владивосток, 2003. С. 82-85.
19. Симонов С.Б., Кушнарёва Т.В., Симонова Т.Л., Компанец Г.Г., Слонова Р.А. Динамика инфицированности мышевидных грызунов хантавирусом и её соответствие структуре их населения в Приморском крае // Хантавирусы, геморрагическая лихорадка с почечным синдромом. Науч.-практич. конференция, 24-25 сентября 2003 г. Владивосток, 2003. С. 79-82.
20. Слонова Р.А., Кушнарёва Т.В., Компанец Г.Г., Якубович Н.В., Яшина Л.Н., Симонов С.Б.Особенности эпидемического проявления геморрагической лихорадки с почечным синдромом в очагах циркуляции хантавируса генотипа Amur // Актуальные проблемы здоровья населения Сибири: гигиенические и эпидемиологические аспекты. Мат-лы V межрегиональной научно-практической конф. с международн. участием. Омск, 25-26 ноября 2004 г. Омск, 2004. Т. 2. С. 187-192.
21. Simonov S.B., Kushnareva T.V., Simonova T.L., Slonova R.A, Kompanets G.G.Structure of small rodent population and their Hantavirus infection in Southern Primorie, Russia // The 6 th International Confererence on Hemorragic Fever with Renal Syndrom (HFRS), Hantavirus Pulmonary Syndrome (HPS) and Hantaviruses. Conference Program and Abstracts. 23-25 June 2004. Seoul, 2004. P. 113-114.
22. Kushnareva T.V., Kompanets G.G., Slonova R.A, Yashina L.N., Yacubovich N.V., Simonova T.L., Simonov S.B. Some factors associated with Hantavirus infection in ecologically different Apodemus rodents // Ibid. - P. 115.
23. Yashina L., Oleinik O., Slonova R., Kuzina I., Kushnareva T., Kompanets G., Simonova T., Simonov S., Morzunov S. Genetic analysis of hantavirus in red-backed vole Clethrionomys rufocanus in Primorsky region of Russia // Ibid. - P. 37.
24. Simonov S, Simonova T., Slonova R., Kompanets G., Kushnareva T., Simonov P. Ecological and epidemiological zoning of the Primorye region (Russia) on Hantavirus infection // VII International Conference on HFRS, HPS and Hantaviruses. Abstract book. June 13th-15th, 2007.Buenos- Aires, Argentina, 2007. - Buenos-Aires, Argentina, 2007. - P. 138.
25. R. Slonova, G. Kompanets, T.Kushnareva, I. Maksyoma, T. Simonova, S. Simonov Epidemiology of hantavirus infection in the south of Far East of Russia (Primorye region) // Ibid.- P. 93.