

И залежныефитосистемы тувы: структурно-функциональнаяорганизация и оптимизация природопользования
На правах рукописи
ДУБРОВСКИЙ НиколайГригорьевич
СТЕПНЫЕ И ЗАЛЕЖНЫЕФИТОСИСТЕМЫ ТУВЫ:
структурно-функциональнаяорганизация и оптимизация
природопользования
03.00.05 – Ботаника
03.00.16 – Экология
Автореферат
диссертации на соисканиеученой степени
доктора биологическихнаук
Улан-Удэ
2009
Работа выполнена накафедре общей биологии Тывинскогогосударственного университета и накафедре ботаники Бурятскогогосударственного университета
Научныйконсультант:
доктор биологическихнаук, профессор
Бимба-ЦыренБатомункуевич Намзалов
Официальныеоппоненты:
доктор биологическихнаук, профессор ТатьянаПетровна Анцупова
доктор биологическихнаук, профессор ИванИкидиусович Илли
доктор биологическихнаук Вера ИвановнаВласенко
Ведущееучреждение:
Алтайскийгосударственный университет,
г. Барнаул
Предварительная датазащиты « 29 » мая 2009г. в ______ часов на заседаниидиссертационного совета Д 212.022.03 вБурятском государственном университете поадресу: 670000, г. Улан-Удэ, ул. Смолина, 24а,Бурятский государственныйуниверситет
Факс: (3012) 21-05-88; E-mail:[email protected]
E-mail: [email protected]
С диссертацией можноознакомиться в библиотеке Бурятскогогосударственного университета
Автореферат разослан « ____ »___________ 2009 г.
Ученый секретарь
диссертационного совета,
к.б.н. Н.А.Шорноева
ВВЕДЕНИЕ
Актуальностьтемы. Изучение растительногомира – этойсложнейшей материальной системы нашейпланеты, во всем его многообразии и на всехизвестных уровнях организации относится кчислу важнейших задач биологической науки.
За последнее время визучении растительного покрова Сибирисделаны большие успехи. Однако на обширныхпросторах Сибири отдельные части ее всееще остаются мало исследованными, к числутаких регионов и относится Тува.
Исследованиесовременного состояния степей какнаиболее широко распространенного типарастительности, слагающей основуландшафтов как в межгорных котловинах, таки высоко в горах особенно актуально в связис тенденцией нарастания антропогеннойтрансформации степных экосистем. Причем,это связано не только с пастбищнойдеградацией растительности, но и большимимасштабами распашки степей, которые внастоящее время на больших площадяхнаходятся в залежном режиме.
Касаясь региональныхособенностей демутационных процессов,следует отметить факт развития особоймелкобурьянистой стадии, предшествующейразвитию крупного бурьяна, в условияхабаканских и тувинских степей(Голубинцева, 1930; Дымина, 1983; Куминова, 1985). Сдругой стороны, на залежах по фонукаштановых почв в Бурятии наблюдаютсяслучаи выпадения бурьянистой стадии (Быкови др., 2003). В связи с этим возникланеобходимость специального изученияособенностей зарастания заброшеннойпашни, видового состава растительности,стадий зацелинения и их длительности,возможности восстановления плодородияпочвы при зарастании залежей вЦентрально-Тувинской котловине РеспубликиТыва. В настоящее время площадь пашни,подвергшейся стихийной консервации вреспублике по официальным даннымсоставляет 59 тыс. га. Этой малоизученной иактуальной проблеме посвящена настоящаядиссертация. Кроме этого, познаниеструктуры и динамики растительногопокрова степей, продуктивности имногообразного влияния антропогенныхфакторов на степные пастбища припреимущественном овцеводческомнаправлении хозяйства Тувы представляеттакже и большой практический интерес.
Цель и задачиисследования. Целью работы являетсявыявление флористического ифитоценотического разнообразия степных изалежных фитоценозов Тувы, а также ихструктурно-функциональных особенностей всвязи процессами демутации растительностистепных залежей.
Для достиженияпоставленной цели предусматривалисьследующие задачи:
1. Выявить состав флорыстепей и степных залежей осуществитьсистематический, биоморфологический иэколого-географический анализ;
2. Сформулироватьосновные положения принятой классификациии на ее основе дать полную геоботаническуюхарактеристику выделеннымтаксономическим категориям степной изалежной растительности;
3. Установить основныестадии в сукцессии залежнойрастительности с характеристикойструктуры сообществ и динамикипродуктивности надземной фитомассы;
4. Исследовать микробныесообщества в опадах залежных фитоценозов иопределить сезонную динамику численностимикроорганизмов – бактерий деструкторов
5. Определить скоростьмикробиологической деструкции целлюлозы ибелка в исследуемых растительныхсообществах, а также изучить характердеструкции в зависимости от абиотическихфакторов окружающей среды (температуры ивлажности)
6. Оценитьсовременное состояние степных и залежныхэкосистем и выработать подходы кэколого-адаптивной концепции вприродопользовании в Тыве.
Материалы и методыисследования. Входе выполнения данной работы былипроведены полевые исследования с 1995-2008 гг.Собран гербарный материал, включающийболее 2300 гербарных листов, сделаны более 720полных геоботанических описаний. Дляизучения видового состава растенийиспользовали: «Определитель растенийТувинской АССР», «Флора Сибири» и другиеисточники.
Сбор материалапроизводился c использованиемобщепринятых методов геоботанических ифлористических исследований. Проведеныисследования степных и залежныхфитоценозов кожуунов: Барун-Хемчикский,Улуг-Хемский, Дзун-Хемчикский, Кызылский,Тандынский, Эрзинский, Овюрский,Монгун-Тайгинский и Пий-Хемский. Названиявидов сосудистых растений приведены пофлористической сводке С.К. Черепанова (1995).Географические группы выделены по В.М.Ханминчуну (1980). Жизненные формы выявленыпо классификации И.Г. Серебрякова (1964),экологические группы – по А.В. Куминовой(1960).
При классификациирастительности использовандоминантно-детерминантный подход,сочетающий критерии флористической иэколого-фитоценотической классификаций,обоснованной во флороценогенетическойконцепции П.Н.Овчинникова (1947), Р.В. Камелина(1979), Б.Б.Намзалова (1994). При систематизациифитоценозов применена компьютернаяпрограмма TURBO (VEG) для первичной обработкигеоботанических описаний.
Исследования помикробиологической деструкциирастительных остатков проводились в 2005-2007гг. в Центрально-Тувинской котловине напробных площадках, соответствующихосновным стадиям зацелинения залежныхфитоценозов. Продуктивность наземнойфитомассы исследуемых растительныхсообществ определяли методом укосов, впяти повторностях (Родин,1968). Исследованиескорости разложения растительного опадапроводили методом механической изоляции(Перель, Карпачевский,1968). Общуюбиологическую активность растительногоопада (протеазная и целлюлазная) изучалиметодом аппликации (Теппер и др.,1987). Учетчисленности микроорганизмов проводилиметодом предельных разведений. Аэробные ианаэробные целлюлозоразрушающие бактерии(ЦРБ) выращивали в жидкой среде Гетчинсона(Романенко, Кузнецов,1974) с добавлениемфильтровальной бумаги. Учет численностиаэробных и факультативно-анаэробныхпротеолитиков, амилолитиков иглюколитиков проводили в агаризованнойсреде Пфеннига (Phenning, 1965) с добавлением 1,5%пептона, крахмала и глюкозысоответственно.
Научная новизна. Впервые изученвидовой состав высших сосудистых растенийстепей и растительности залежнойрастительности региона исследования и врезультате установлен список флоры степейТувы. Последняя составлена 785 видами высшихсосудистых растений (272 рода и 54 семейства).Проведен разносторонний детальный анализфлоры.
Проведена оригинальнаяклассификация видового составарастительности залежей Тувы (110 видовсосудистых растений, 27 семейств и 80 родов) сустановлением трех категорийфлористического комплекса залежей (ФКЗ), аименно –типично залежных, переходных и инвазийных(или случайно залежных) типов ФКЗ. Долевыесоотношения последних в растительностизалежей имеют индикационное значение поособенностям их демутации. Выявлено всетаксономическое разнообразие степныхсообществ Тувы. Осуществленаэколого-фитоценотическая классификациястепной растительности с характеристикойвыявленных синтаксонов. Выделено 28формаций, 13 групп формаций, относящиеся к 4классам формаций (или подтипам) степнойрастительности.
Впервые дляЦентрально-Тувинской котловины Тывыпроведено комплексное изучение залежнойрастительности, выявлены особенностидемутации, изучена динамикапродуктивности фитоценозов, а такжечисленности микроорганизмов, участвующихв деструкции опада. Определен вкладразличных физиологических групп бактерийв аэробную и анаэробную деструкциюорганического вещества растительныхостатков. Даны количественные показатели оскорости разложения опада в залежныхфитосистемах Тувы, изучено влияниеабиотических факторов (температуры ивлажности) на темпы микробиологическойдеструкции.
Изучены особенностипастбищного использования растительностистепных экосистем, закономерностей ихантропогенной трансформации и процессовдемутации на залежах, а также определеныприоритеты эколого-адаптивногоземлепользования как перспективной длясухостепных ландшафтов межгорных котловинТувы.
Практическаязначимость исследования.В результате проведенныхисследований, впервые наиболее полновыяснена типологическая структура степныхпастбищ Тувы, их продуктивность,хозяйственное состояние. Полученныематериалы по флоре и фитоценотическомуразнообразию степей и залежей могут бытьиспользованы в сравнительныхэкологических и фитогеографическихисследованиях растительности гор югаСибири и Центральной Азии. Содержащиеся вработе материалы могут послужить ценнымисточником при подготовке определителярегиональной флоры, при флористическом игеоботаническом районировании, а также дляэкологического мониторинга исвоевременного принятия мер по сохранениюбиологического разнообразия экосистем ивыявления отрицательных воздействий насреду обитания растений и сообществ.Анализ результатов исследований залежныхэкосистем Тувы позволит обосновать новуюконцепцию экологического земледелия дляаридных и континентальных районовВнутренней Азии. Изменение показателейпочвенного плодородия при выведении пашнив залежь выявили перспективность данногоприема для его восстановления.
В целом, основнымирезультатами научно-исследовательскойработы будут пользоваться другиеведомства по охране и использованиюприродных ресурсов Тувы, Тывинскийгосударственный университет и другиеобразовательные учреждения. Собранныематериалы пополнили гербарные фондыБурятского и Тывинского госуниверситетов.Они могут использоваться при подготовкеспециалистов биологов и экологов в курсахсистематики и экологии растений,геоботаники и биогеографии.
Защищаемыеположения.
1. Флора степей Тувы,насчитывающий 785 вида высших сосудистыхрастений, отражает особенности буферногоположения территории на стыке бореальных ипустынно-степных экосистем. При этомпозиции горностепныхюжносибирско-монгольских ицентральноазиатских видов ксерофитнойэкологии наиболее значительны.
2.Эколого-фитоценотическая классификациястепной растительности с использованиемдоминантно-детерминантного подходапозволила выделить 28 формаций, 13 группформаций и 4 класса формаций (подтипов).Однако, объективизация выделениясинтаксонов может быть достигнута сиспользованием метода экологическойординации сообществ степей.
3. Анализ видовогосостава растительности залежей Тувы (294видов сосудистых растений, 110 – только вЦентрально-Тувинской котловине) сдифференциацией на три типафлористических комплекса залежей (ФКЗ), аименно –типично залежных, переходных и инвазийных(или случайно залежных) позволяетпрогнозировать динамические тенденции впроцессах демутации и могут служитьиндикаторами стадий зацелинения залежей.
4. Микробиологическаядеструкция растительного опада являетсяважнейшим процессом, определяющимбиологический круговорот элементов вприроде. При этом вклад различныхфизиологических групп бактерий в аэробнуюи анаэробную деструкцию органическоговещества растительных остатков имеютрегиональные особенности на различныхстадиях демутации. Это касается преждевсего скорости разложения опада в условияхсухостепных экосистем Тывы.
Апробация работы. Материалыисследования доложены и обсуждены наВсероссийской научно-практическойконференции с международным участием«Опыт и традиции этническогоприродопользования в Байкальской Сибири»(г. Улан-Удэ, 2003), Всероссийской научнойконференции с международным участием«Проблемы сохранения разнообразиярастительного покрова Внутренней Азии» (г.Улан-Удэ, 2004), на VIII МеждународномУбсунурском симпозиуме «Убсунурскаякотловина как индикатор биосферныхпроцессов в Центральной Азии» (г. Кызыл, 2004),на VII Международной конференции «Природныеусловия, история и культура ЗападнойМонголии и сопредельных регионов» (г.Кызыл, 2005), Всероссийской конференции«Биоразнообразие и пространственнаяорганизация растительного мира Сибири,методы изучения и охрана» (г. Новосибирск,2005), Международной научно-практическойконференции «Проблемы ботаники ЮжнойСибири и Монголии» (г. Барнаул, 2002, 2006), Пятоммеждународном симпозиуме «Степи СевернойАзии» (г. Оренбург, 2006), Региональномнаучно-методологическом семинаре«Этническая экология и традиционноеприродопользование на рубеже веков» (г.Улан-Удэ, 2006), V научнойконференции РАЕ с международным участием«Экология и рациональноеприродопользование», (Египет, г. Хургада2007), Всероссийскойконференции с международным участием«Синантропизация растений и животных» (г.Иркутск, 2007), Международной научнойконференции «Мир кочевой цивилизации:история и современность» (г. Чита, 2007),Международной научно-практическойконференции «Биоразнообразие и сохранениегенофонда флоры, фауны и народонаселенияЦентрально-Азиатского региона» (г. Кызыл,2007), заседаниях кафедры ботаники БГУ икафедры общей биологии ТывГУ (2004, 2005, 2008).
Исследования былиподдержаны грантом РФФИ «Разработкастратегии сохранения редких растений ифитоценозов Тувы» (2005-2006, №05-04-97279), ГрантРГНФ «Комплексное социально-экологическоеисследование особенностей номадногоприродопользования аборигенных этносовгорных районов Южной Сибири. (2005-2007, №05-06-06487а), Правительства Республики Тыва (№54, 2005).
Публикации. По теме диссертацииопубликовано 32 работы, в том числе 5монографий и статей, из них 7 врекомендованных ВАК изданиях.
Объем работы. Диссертационнаяработа изложена на 345 страницах. Состоит извведения, 7 глав, выводов, списка литературыи приложения. Материал представлен 19рисунками (фотографии, схемы, диаграммы) и 20таблицами. Список литературы насчитывает312 наименований.
ГЛАВА I. Историяисследования растительности степей изалежей
Первые содержательныегеоботанические ифлористические сведения о растительноммире Тувы появились болееста лет назад в отчетахэкспедиций, исследовавших Центральную Азию. Этопутешествия Г.Н.Потанина(1876-1877 и 1879-1880 гг.),A.В. Адрианова(1881 и 1883гг.), Г.Е.Грум-Гржимайло (1902-1903 гг.) и В.И.Дорогостайского (1907 г.)Исследователи, совершившие специальные ботанические экспедиции –П.Н.Крылов (1903)и Б.К.Шишкин(1914), X. Принтц (Printz, 1921).
Новый этап ботаническихисследований Тувы относится к периодуустановления Танну-Тувинской Народной Республики в 1921 г., и он связан сработами экспедиции ВАСХНИЛ,начиная с 1934 г.,под руководством В.Г.Варварина(1950). В 1928 г. работала Саянская ботаническаяэкспедиция Томского университета подруководством В.В.Ревердатто(Ревердатто, 1931).
Третий этап изучениярастительного покрова, в частности, горных степей Тувы начинается с 1940-хгг. С 1945 по 1949 гг. исследованиямифлоры и растительности Тувы занималась К.А.Соболевская (1950,1953, 1958). КромеК.А. Соболевской в 1940-1960-е гг. ботаническиеисследования в Туве проводят А.И.Шретер(1953), А.В.Калинина (1957),С.В.Гудошников (1964), М.А.Савич (1975).
Четвертый, современныйпериод ботанических исследований в Туве, начинается с 1970-хгг. Ониначинаются с флористическихработ И.Ю.Коропачинского иИ.М.Красноборова. Начиная с 1975 по 1981гг., изучением растительного покроваТувы занималась лаборатория геоботаники ЦСБС СО АН СССР, подруководством А.В.Куминовой.Исследования степнойрастительности Хемчикской котловины вЗападной Туве ведет Б.Б.Намзалов (1978; 1979, 1982, 1985). Различным аспектам анализа структуры растительности ифлоры горных степей региона посвятилисвои работы С.А.Тимохина иВ.М.Ханминчун (1976, 1977), В.М.Ханминчун (1975, 1980), М.Н. Ломоносова (1977),Э.А. Ершова (1982а, б),В.А.Шоба (1982),Г.А. Зверева (1982), М.П. Данилов(1981), А.В.Куминова (1982), Г.Г.Павлова и др.,(1982), Д.Н. Шауло (1982), И.М.Красноборов (1986) и др.
После некоторогоперерыва, начиная с 1995 г. и,особенно с 2000 г. возобновляются исследованиярастительности степных экосистем регионас приоритетом анализа трансформациистепей,изучения структурно-функциональных ихособенностей. В постановке и реализацииисследовательских программ степныхэкосистем велика роль Убсу-Нурского центрабиосферных исследований под руководствомВ.В.Бугровского и С.С.Курбатской, а такжеестественнонаучных кафедр Тывинскогогосуниверситета с участием и координациейС.О.Ондара, Ч.Т.Сагды, Б.Б.Намзалова,Л.К.Аракчаа и Н.Г.Дубровского. Итоги исследованийпоследнего времени отражены в рядедиссертационных работ. ПубликацииА.Д.Самбуу (2003,2004) и Ч.С.Кыргыс (2002), посвященны анализупродукционно-деструкционных процессов сухихстепей Убсу-Нурской котловины.Рассмотрению геоэкологических аспектовфункционирования степных ландшафтовпесчаных массивов Тувы посвятила своиработы С.К.Кужегет (2006). Степи нагорьяСангилен, особенности ее флористическогосостава, фитоценотического разнообразия ипространственной организации сталиобъектом исследования К.В.Кыргыс (2006).Флористические исследования в регионепродолжил А.М.Самдан (2007), раскрывшийструктуру флоры плато Алаш.
Таким образом, степень изученностистепной растительности региона такова, чтонеобходимо обобщениеимеющихся материалов. Приэтом особого внимания заслуживают анализрезультатов трансформации степей(пастбищного и залежного)и выявление их динамических тенденций. Залежнаярастительность Тувы до сих пор не былапредметом специальных исследований.
ГЛАВА II.Природно-климатические особенности Тувы(орография, климат, почвы)
Тува,расположенная на юге Средней Сибири ипримыкая к территории Монголии,приурочена между 49° и 52° с.ш. и 87°и 96° в.д., характеризуетсясильно расчлененным рельефом. Современный макрорельеф Тувысформировался напротяжении мезозоя и кайнозоя, причем орогенетическая активизация территории происходила по линиям древнейшихтектонических разломов (Ефимцев, 1961; Девяткин, 1965; Чернов, 1988). В мезозое, вконце юры или раннего мела, устанавливается платформенный режим.В этот период на фонемедленногосводовогоподнятиятерриториипреобладалипроцессыденудационного выравнивания. Речной сток из Тувинской котловины былнаправлен на юг, в сторону Гоби. Рельефвыгляделв виде низких гряд, холмисто-увалистых возвышений на фоне обширных выровненных равнин.
Новейшийорогенныйэтап развития рельефа Алтае-Саянскойобласти –этап альпийского тектогенеза вызвали блоковые поднятия, выявившие уже достаточно четко основные черты современного рельефа Тувы.Это оформление Саян, образование горстового хребта Западный Танну-Ола (Маслов, 1948,Зятькова,1969). В этоже время в результате слабых восходящих движений сформировался ряд внутригорныхвпадин в Тувинской котловине, обособившие Хемчикскую, Улуг-Хемскую иКызыльскуювпадины, поднявшимися низкогорными грядами -Адар-Таш, Берт-Дат. Горные сооружения, испытавшие постепенное поднятие, приобреличерты резко расчлененного рельефа(Западный Саян, Шапшальский). Отдельные хребты, испытавшие более резкое вздымание, сохранили остатки поверхности выравнивания навысотах2500-3000 м над ур.м. (Западный Танну-Ола, нагорьеСангилен). В пределах плато и нагорий характернанаибольшая дифференцированность поднятий, на фонесреднегорного сильно расчлененного рельефа возвышаются отдельныевершины до 3200-3600 м (Сангилен, Алашское плато) (рис. 1).

Рис. 1.Орографическая схема Тувы
Сложно организованнаяорография территории, располагающаяся вантициклоническом фасе крупнейших горныхсооружений юга Сибири - Саян и ЦентральногоАлтая, предопределила климатическиеусловия Тувы. В целом территориянаходится в условиях умеренного и недостаточногоувлажнения (Поликарпов и др., 1986). Распределе-ние осадков порегиону чрезвычайно неравномерное. С одной стороны,территория недоступна воздействию влажныхвоздушных масс западного переноса; сдругой, регион открыт мощным влияниямаридного и экстраконтинентального климатаЦентральной Азии. Наиболее экстремальныйгидротермический режим создается вдождевой тени экранирующих хребтов(юго-западные части Хемчикской иУбсу-Нурской котловин, где сумма осадковдостигает 80-150 мм в год). С другой стороны,создаются оптимальные условия на наиболееудаленных от магистральных хребтов, горныхвозвышениях, где транзитные воздушныепотоки верхнего яруса тропосферыоставляют большую часть своей влаги. Такиеситуации создаются на северныхмакросклонах восточных отрогов ЗападногоТанну-Ола, где выпадает до 700-800 (900) ммосадков, тогда как на шлейфах южногомакросклона этих хребтов – всего около 100-150 мм.
Сложный рельеф иконтрастный климатическийрежим территории определяют большоеразнообразие ипространственную неоднородностьпочвенного покрова Тувы. Для всех типов почвхарактерна малая мощностьпочвенного профиля из-за неглубокогопроникновенияпочвообразовательных процессов в толщуматеринских пород (Кононова, 1963). Почти все типы почв в горностепных ландшафтах отличаются грубым механическимсоставом, содержат большое количествощебнистого материала, на небольшой глубине подстилаются осадочными иликристаллическими материнскими горными породами (Зятькова, 1969).
Степныепочвы тувинских котловинотносятсяк почвам каштанового ряда, представленныхподтипамитемно-каштановых, каштановых исветлокаштановых почв. На первом месте по распространениюстоят каштановыепочвы (Носин,1963), по механическомусоставу в основном легкосуглинистые исупесчаные разновидности снеглубоким(в пределах 50 см от поверхности) залеганием щебня или песка. В целом для каштановых почв характернымалая мощность, легкиймеханический состав, высокое содержаниескелетных включений (до 50-80%),незначительные запасыгумуса - 2,3-4,0 %. Среди степныхпочв особого внимания заслуживают почвыверхних поясов аридных горТувы. Характерными чертамиэтих почв являютсяукороченный профиль, присутствие на поверхностях органно-генногогрубогумусного горизонта свысоким содержанием в нем гумуса (до 12 %),слабокислой реакциейверхних горизонтов ищелочной - в нижних (Максимович, Ногина,1984).
ГЛАВА III. Методологияизучения разнообразия фитосистем
Биологическое разнообразие какфундаментальное понятие классических науко биоте (ботаника, зоология) включает всемногообразие живых организмов растений,животных и микроорганизмов, а также ихсообществ и ландшафтных комплексов. Одноиз самых лаконичных определений понятия«биоразнообразие» (БР) дано Б.А. Юрцевым «БР- разнообразие организмов и их природныхсочетаний» (1992). Однако, автор принимая забазовый уровень видовое разнообразие посути развивает (альфа) - таксономическую концепциюБР, означающая разнообразие генофонда,начиная с популяционного уровня до фено- игенотипов.
За детализацию видовогоразнообразия на уровне особей и популяций,полагая что они отличаются погенетическим, фенотипическим и возрастнымструктурным особенностям, ратуетЮ.А.Злобин (1996). Им обосновывается тезис отом, что «БР проявляется на всех уровняхорганизации живой материи от особей добиосферы» (с.31). Однако столь дробноеподразделение категорий БР может привестик такому явлению как континуум БР. При этоммы можем «потерять» такую узловую точкупроявления БР как вид. Вид, являясьсистемой популяций, включает всемногообразие особей как структурнуюоснову биологического вида и являетсяудобной «инвентаризационной единицей»(Мэгарран, 1992).
Другая позиция вориентации приоритетов к определению БРвыработана в подходе А.А.Тишкова (1992). Помнению автора ключевым является«разнообразие элементовпространственно-временной структурыбиомов». Это прежде всего означаетразнообразие экосистем как проявлениеценофонда (Седельников,1997). Понятие«ценофонд» отражает разнообразиебиохорологических структур от уровняэлементарных экосистем (сообществ), ихсукцессионных серий до биомов. Сутьподхода - через выявление ценофонда,разнообразия зональных экосистем с учетомредких и исчезающих его компонентов,обеспечить сохранение потенциала БР видови их генофонда. Этих же позицийпридерживается А.Ю.Королюк (1994), по егомнению «зная структуру комплексарастительности, выражающуюся в наборе иплощадном соотношении слагающих егорастительных сообществ, мы можем оцениватьи прогнозировать присутствие наисследуемой территории как групп, так иотдельных видов растений» (с.590). Данныйподход в какой-то мере раскрывает (бета) компоненту БРкак разнообразие сообществ
Наиболее полным, с точкизрения принципов системности ииерархичности, остается подход Р.Уиттекера(1972). Понятие «Биоразнообразие» авторрассматривал как системувзаимодополняющих на разных уровняхструктурных элементов экосистем (в данномслучае экосистема рассматривается какнадценотическая, ландшафтная единица). Приэтом уровень разнообразия видов (), на следующемуровне рассматривается как ценотический() и наконец,более высокий уровень интеграциипредусматривает БР определенныхландшафтов (/гамма/- разнообразие). Причем,разнообразие в пределах ландшафтаозначает многообразие как видов, так исообществ (рис. 2).

Рис. 2.Схема узловых уровней биоразнообразияфитосистемы
По мнению многихэкологов видовое разнообразие намногосильнее связано с разнообразием сообществи архитектурной сложностью местообитаний,т.е. ландшафтной структурой. Чемразнообразнее ландшафт, тем вышеэкологический объем местообитаний.Многообразие местообитаний по образномувыражению Б.М.Миркина (1981, 1986), являетсяследствием "увеличения гиперпространстваниш сообщества" (1986, с.55). Последнее означаетобеспечение полноты популяционногоспектра видов, включая и временной факторсвязанный с микроэволюционными процессамив сообществах (формирование видов).Выявление разнообразия сообществ какследствия реакций биоты на воздействияразличных факторов среды и составляет цельизмерения В-разнообразия. В данном случаеценоз или сообщество организмов являетсяконкретным выражением -разнообразия, такжекак особь - при определении -разнообразия.Разнообразие сообществ выражается числомтаксонов или типов сообществ (ассоциаций,формаций, классов и т.д.).
Особый интерес вызываетследующий уровень БР (-разнообразие) биоты,разнообразие наименьшейбиохорологической единицы - ландшафта.Данный уровень понимания БР до сих поростается достаточно неразработанным. Вчастности В.И.Василевич (1992) считаетцелесообразным рассматривать и - разнообразия какодин тип БР. По мнению автора, сменасообществ по каким-либо градиентам(например, по ряду увлажнения) означаетэкологический диапазон изменений всетого-же -разнообразия, комбинации сообществв границах ландшафтов. Подобныетерриториальные системы синтаксоновназываются ценохорами (Сочава, 1972) илисигматаксонами (в подходах флористическойкласс-сификации растительности). Сам Р.Уиттекер (1972) -разнообразие рассматривал какобщее разнообразие группы участков и -разнообразий, но приэтом не дал четких критериев: "что нужносчитать базовой счетной единицей данногоразнообразия?". Однако, Р.Уиттекер выделялособую категорию разнообразия (-эпсилон), какрегиональную. Сюда включалисьразнообразие группы территорий соднотипной генетической "конструкцией"ландшафтов. По сути это было разнообразиекрупных биогеографических единиц (районов,провинций, областей).
После краткогорассмотрения общих теоретических позицийконцепции биоразнообразия, рассмотримсхематическое представление методологиианализа биоразнообразия растительногомира (рис. 2). Уровень -разнообразия показываеттаксономическое разнообразие видов,которые отражены в региональныхфлористических сводках, определителяхрастений.
-разнообразие - разнообразиерастительных сообществ или фитоценозов.Познание данного уровня начинается сустановления группировок из сочетанийособей видов растений и систем ихпопуляций в границах однородного контураземной поверхности минимальнойразмерности. Это исследованияпарцеллярной и синузиальной конструкциисообществ, явлений мозаичности и ярусностиструктуре сообществ, что крайне важно дляопределения механизмов их устойчивости.Пространственная разнообразие и временнаядинамика сообществ, выраженная вфенологических спектрах, сукцесссионныхсхемах и картографических моделяхрастительного покрова, а такжетипологическое разнообразие фитоценозов,представленная в виде иерархических схемклассификации синтаксонов составляетсодержание раскрытие B-разнообразиярастительности.
Ландшафтноеразнообразие растительности, котороетрактуется как -разнообразие, остается ещемалоразработанной. Пока не выработанединый подход в раскрытии данного уровня. Внашей трактовке этоландшафтно-геоботанические исследования,ориентированые на выявление ценотическогоразнообразия сообществ ивнутриландшафтных структурных категорий вединстве с физико-географическимиусловиями - почвенными,геоморфологическими, климатическими,литологическими и т.д. Важнейшим являетсяанализ пространственной структуры территориальныхединиц растительности (ландшафтныхкомбинаций) с целью познания механизмовструктурирования и интегрированиясообществ и комбинаций в единую "ткань"растительного покрова. Непременнымусловием работ является заложениеэталонных полигонов с постановкой на нихдетального крупномасштабногокартографирования растительности.Дальнейшая типизация (классификация)выделенных однородных (сообществ) инеоднородных (комбинаций) единицрастительности полигонов дает системноепредставление о структуре растительного покроварегионов.
В целом, из трех базовыхуровней биоразнообразия (,, ) ключевымпредставляется ландшафтная, котораявключает в себя элементы первых двух -конкретные особи видов растений ифитоценозы. Действительно, вид можносохранить лишь сохранив сообщества, вкотором он отмечается, а разнообразиеценозов является следствием разнообразияместообитаний, морфогенетическихповерхностей рельефа. Отсюда каждаяприродная комбинация обеспечивает нетолько разнообразие ценозов, но и системыпопуляции видов растений, как структурныхчастей сообществ. В этом заключаетсясистемообразующая роль комбинацийландшафтного уровня и поэтому, значимостьисследований типов структур РП будетвызывать все больший интерес (Намзалов,Дубровский, 2007б). В настоящей работе всоответствии с изложенной выше концепциейбиоразнообразия раскрыта видовая () и ценотическая() организациярастительности степей Тувы, включая ихпастбищно- и залежно трансформированныеварианты.
ГЛАВА IV. Степи Тувы,анализ структуры флоры степей и залежныхземель
Степи в Туве являютсяшироко распространенным типомрастительности и занимают 2811 тыс. га.(Ершова, Намзалов, 1985). Как все островныестепи Южной Сибири, ониприурочены к межгорным понижениям иокаймляющими их со всехсторон предгорьями хребтов и нагорий. ВТуве они слагают основуландшафтов Тувинской (Хемчикская иУлуг-Хемская) и Убсунурской котловин, рядапериферийных небольших межгорных впадин (Эдыгейская,Карахольская, Турано-Уюкская) ивстречаются по южнымсклонам в лесном поясе всех горныхсистем.
Основныестепныерегионы:
- Тувинская котловина (Хемчикская,Улуг-Хемская и Турано-Уюкская впадины).
- Убсунурский (северная окраина Убсунурскойкотловины и южныймакросклон Западного и ВосточногоТанну-Ола).
- Монгун-Тайгинский (горный массивМонгун-Тайга и южный макросклон хребтаЦаган-Шибэту).
Флора степей Тувывключает 785 видов, принадлежащих к 272 родам и 54 семействам. Список флоры составлен на основании гербарных сборовавтора, а также анализапубликаций К.А. Соболевской (1953),И.М.Красноборова ссоавторами (1973, 1975, 1980), А.В.Положий (1974, 1980), А.С.Ревушкина (1981), К.В.Кыргыс(2006), А.М.Самдан (2007) и других. Флора степей Тувыбогата, что особенно наглядно видно, еслисопоставить ее с флорамиостровных степей Алтая, Хакасии,Забайкалья. Согласно даннымсистематического анализа флоры, к многородовым семействам относятся:Asteraceae (31 род),Роасеае (25), Fabaceae (14), Rosaceae (10), Chenopodiaceae (13), Brassicaceae (23). По видовому составу всепредставленные семейства можно разделить на три основныегруппы:
- В группу ведущихсемейств, включающих от 20 видов и более,входят 13 семейств, ихудельный вес в сложении флоры высокий– 83,7% (655 видов). Сюда входятследующие семейства: Asteraceae(120), Роасеае (89),Fabaceae (79), Rosaceae (42), Chenopodiaceae (37),Liliaceae(30).
- Группа семейств,включающая от 4 до 19 видов, состоит из 13 семейств, всоставе которых 80 видов (10,2% от флорыстепей): Primulaceae (10), Umbelliferae (10), Polygonaceae (13), Caryophyllaceae (11), Gentianaceae (9), Campanulaceae (12) и др. Виды, объединенные в этисемейства, образуют группу разнотравья,ценотическая роль которой внастоящих крупнозлаковыхи луговых степях становится одной из ведущих.
- Группа семейств снебольшим числом видов (от 1 до 4) пред-ставительна: 25семейств и 47 видов (6,1%). Многовидовыми родами являются: Artemisia (38), Astragalus (27), Potentilla (24), Allium (22)(табл.1). Для познания флоры тех или иных регионов не менее важныроды с небольшим числом видов, особенно монотипные. Во флорестепей Тувы это - Arctogeron gramineum, Cymbaria daurica, Nitrariasibirica, Nanophyton erinaceum и др. Эти виды имеют разный генезис ив комплексе хорошо отражают сложныйпроцесс становления флоры исследуемогорегиона (Дубровский, Намзалов, Кыргыс,Самдан, 2007).
Таблица 1
Количество видовосновных родов флоры степей Тувы иосновных районов
Род | Количество видов | Числовидов в районах | |||
| шт. | % от общегосостава | I | II | III | |
| Stipa Festuca Carex Chenopodium Allium Potentilla Astragalus Oxytropis Saussurea Artemisia | 12 10 16 9 22 24 27 20 9 38 | 1,50 1,27 2,00 1,15 2,80 3,06 3,40 2,50 1,15 4,80 | 10 3 11 7 18 11 15 8 5 30 | 8 6 7 7 9 17 20 6 4 26 | 5 9 7 3 14 8 6 10 5 13 |
| Общее числовидов | 785 | 503 | 367 | 263 | |
Примечание. I - Тувинскаякотловина, II -Убсунурский, III -Монгун-Тайгинский
Экологическийанализ. Разнообразиеместообитаний, обусловленное расчлененностью рельефагорной страны, характером микроклимата, почв и подстилающих горныхпород, наложило глубокий отпечаток на особенности флорыстепей Тувы. Экологическаяобстановкаспособствовала развитию чертксероморфизма почти во всех биологических группах растений.
Своеобразие степямТувы придают высокогорные степи, сложенные криофитами(криоксерофитами), формирование которых шло в условияхгляциальных и перигляциальных ландшафтов ледникового периода(Ревердатто, 1934, 1940; Соболевская, 1948; Положий, 1964 и др.). Среди них выделяются Festuca tschujensis, F. kryloviana, Kobresia filifolia, Oxytropis macrosema, Artemisia depauperata и др.
Анализ экологическогоспектра флоры степей показалпреобладающее значение группы ксерофитов,а также мезоксерофитов и ксеропетрофитов,что в целом составляет 81% от всех видов. Примерно такое же соотношение сохраняется во флорахкрупнодерновинных,мелкодерновинных настоящих, опустыненныхи луговых степей. Многиевиды из этой группы являютсяэдификаторами степныхсообществ. Группаксеропетрофитов как растений каменистыхместообитаний составляет26,9% от флоры степей. К этой группе мыотносим следующие виды:Alyssum lenense, Orostachys spinosa, Eritrichium pectinatum, Peucedanum baicalense, Thalictrum petaloideum, Allium stelleranum, Potenlilla soongarica, Chamaerhodos altaica, Ch. erecta, Oxytropis eriocarpa и др.
Небольшая по числувидов группа псаммоксерофитов (2,3%) образует несомкнутуюрастительность на песчаныхместообитаниях. Злаковуюоснову песчаных степей слагают Leymus racemosus, Calamagrostis epigeios,Agropyron michnoi идр.
Галоксерофиты создаютструктурную основу солонцеватых степей и составляют 6,5% от всей флорыстепей. Для днищ котловин, надпойменных террас рек с близкимзалеганием грунтовых вод характерны ассоциациисолонцеватых степей (чиевые, волоснецовые,прутняково-чиевые). Помимоопределяющей экологической группыгалоксерофитов участие в сложениисолонцеватых степей принимают настоящие ксерофиты (21,7%),мезоксерофиты (37,2), ксеропетрофиты (12,4%).
Анализ составажизненных форм в исследуемых степях Тувытакже показал ихособенности и своеобразие. Прежде всего, большое значение в степяхТувы имеют ксерофильные кустарники, особенно виды рода Caragana. Из семи видов караган, известных вТуве, в степных сообществах широкораспространены три: Caragana pygmaea, С. bungei, С. spinosa. По количеству видов различных кустарников (35) степи Тувыбогаче, чем соседние островные степи (Хакасские - 20 видов,Алтайские - 22), и по ценотической роли в степных фитоценозахзанимают одно из ведущих мест (Куминова, Намзалов, 1982).
Основу степной флорыТувы составляет жизненная форматравянистых многолетников (79,5%). Онапредставлена дерновиннымитравянистыми многолетниками, причем, кромеобычных в степях Евразииплотнодерновинных злаков, характернымелкие перистые ковыли- Stipa glareosa,S. orientalis. В опустыненных степях заметноповышается роль луков (Allium tenuissimum,. A. mongolicum, A. anisopodium), образующих дерновины за счетнескольких сближенныхлуковиц. Из корневищных биоморфтравянистых многолетников ценотическуюроль играют ксерофильная осочка - Carex duriusculaи виды рода Leymus всолонцеватых злаковыхстепях.
Группаполукустарничков представлена 27 видами, из них доминантамиценозов выступают Artemisia frigida, Kochia prostrata, Ceratoides papposa, реже -Thymus mongolicus. Кустарнички представлены Nitraria sibirica, Ephedra distachya, Oxytropis tragacanthoides, Nanophytonerinaceum. Последние три вида выступают какэдификаторы сообществ. Незначительное участие в травостоестепных сообществ Тувы занимает группа одно- и двулетников(10,5%). Многие из них относятсяк числу адвентивных видов (сорняки).Наиболее обычны Chenopodium aristatum, Kochia sieversiana, Artemisia anethifolia, Tribulus terrestris.
Всяфлора степей делитсяна несколько эколого-фитоценотических групп: 1 - Высокогорно-степные виды,свойственные криофитным (высокогорным) степям. 2 - Горно-степные растения характерныместообитаниям по южнымкрутым склонам 3 - Лесостепные растения - виды, обычные в луговых, разнотравно-дерновиннозлаковых степях. 4 – степные растения представлена видамидерновинно-злаковыхнастоящих степей. 5 - пустынно-степные растения -виды, слагающие сообщества формаций опустыненных степей. 6 -солонцевато-степные видысоставляют в основном гетерогенный флористический комплекс сильноостепненных чиевников. 7 - придаточные растения образуютмалотипичные для степейвиды, встречающиеся в качестве сорных илизаносных. В среднем по всемвыделенным группам более половины (67,8%)состава флоры приходится нагорно-степные и лесостепные виды (табл. 4), собственно степные видысоставляют лишь 28,0%. Однакоанализ по районам показывает иныехарактеристики. С однойстороны, для Убсунурской котловины долястепных (15,8%) ипустынно-степных (9,5%) групп заметно выше,чем в других районах. Сдругой стороны, для нагорья Сангиленосновное значение имеютлесостепные виды (30,6%) при незначительныхпоказателях степных (13,1%) ипустынно-степных (2,5%). Это закономерно, ибоУбсунурская котловина образует северныеоконечности пустынных экосистемСеверо-Западной Монголии, аВосточно-Тувинское нагорье и Сангилен - области древнегогорообразования и издавна были горно-лесными районами, что иопределило господство лесостепных видов.
5.2. Характеристикасистематического, биоморфологического иэколого-фитоценотического состава флорызалежной растительности
Специального изученияфлоры и растительности залежных земельТувы еще не проводились. Однако изучениестепной флоры проводились К.А. Соболевской(1953), А.В. Куминовой (1960), И.М. Красноборовым(1984). Заметный вклад в изучение флоры Тувытакже внесли И.Ю. Коропачинский (1975), С.А.Тимохина (1977), В.П. Седельников (1985), Э.А.Ершова (1985), Б.Б. Намзалов (1981, 1985, 1994), А.Ю.Королюк (1991) и др. Важные и содержательныеидеи и данные в изучении адвентивной флоры,сорной и залежной растительности Тувывнесли А.В. Куминова (1985), Т.А. Терехина (2000).
Список растенийописанных и изученных нами в залежныхфитосистемах в Центрально-Тувинскойкотловине включает 110 видов сосудистыхрастений, относящихся к 27 семействам и 80родам.
По количеству видовлидирующее положение занимают семействаAsteraceae, Poaceae, Brassicaceae, Fabaceae, Rosaceae. Видовое богатство семейств Asteraceae, Poaceae, Fabaceae иChenopodiaceaeхарактеризуют аридные черты флоры. (Юрцев,1988). В то же время в видовом богатствеRosaceae, Brassicaceae, Caryophyllaceae, Ranunculaceaeпроявляются черты гумидности климата.Обилие видов семействах Asteraceae, Poaceae и отчастиFabaceae указываетна бореальный характер флоры, а видовоебогатство семейств Brassicaceae,Rosaceae, Caryophylaceae, Boraginaceae, Chenopodiaceae подчеркивает ее аридные черты, афлора носит смешанный, бореально-степнойхарактер (табл. 2).
Таблица 2
Ведущие семействазалежной флоры
| № | Семейство | Числовидов | % от общегочисла видов | Числородов |
| 1 | Asteraceae | 23 | 20 | 13 |
| 2 | Poaceae | 18 | 16 | 14 |
| 3 | Fabaceae | 10 | 9 | 7 |
| 4 | Brassicaceae | 9 | 8 | 9 |
| 5 | Rosaceae | 7 | 6 | 3 |
| 6 | Caryophyllaceae | 6 | 5 | 4 |
| 7 | Chenopodiaceae | 5 | 4 | 4 |
| 8 | Boraginaceae | 3 | 2 | 3 |
| 9 | Apiaceae | 3 | 2 | 3 |
| 10 | Ranunculaceae | 2 | 1 | 2 |
| Всего: | 86 | 73 | 62 |
Анализ жизненных формзалежной флоры Тувы на моделиЦентрально-Тувинской котловины позволяетнам заключить следующее: численнопреобладают травянистые многолетние(поликарпы) растения (57%). Второй группой поколичеству видов жизненных форм являютсяоднолетники (монокарпы) (22%). Сюда жепримыкают двулетники (9%) и 1-2 летние (6,4%)травянистые растения. Залежная флора малопредставлена полукустарничками (Potentilla bifurca, P. acaulis, Artemisia frigida) (2,7%) икустарниками (Spiraea media, Caraganapygmaea, C. bungei) (2,7%). Они уступаютместо травянистым растениям, так как средисорнополевых растений кустарничков иполукустарничков очень ограниченноеколичество видов и их роль в засоренииполей небольшая.
Итак, в залежной флоре вЦентрально-Тувинской котловине мыотмечаем разнообразие жизненных форм ипреобладание многолетников иоднолетников. Широко распространены видырастений стержнекорневой (53/48%),корневищной (26/24%), длиннокорневищной (13/12%),кисте-корневой (11/10%), корнеотпрысковой (6/5,5%)растений, что является результатомприспособления к резким колебаниямтемператур и недостатку влаги. Ощутимоеколичество одно-, двулетних растенийопределяется максимумом осадков впоздне-летнее время, а также антро-погеннымвоздействием. На каждый год на почвеостается их генети-ческий фонд – семена. Здесьмонокарпы переживают неблагоприятныепериоды. Среди залежных безрозеточныхрастений (48/43%), выделяются стержнекорневыеиз семейств Asteraceae, Brassicaceae,Fabaceae; розеточных (36/33%) изсемейства Poaceae, Cyperaceae, а также полурозеточных (26/24%).
Экологическаяструктура залежной флоры отражаетособенности региональных флор с позицийсоответствия ее состава современнымклиматическим и орографическим условиям,выявляет черты, свидетель-ствующие обэтапах эволюции флоры (Холбоева, Намзалов,2000).
Анализ экологическогоспектра показал господствующие позицииксерофитов (50/46%), мезоксерофитов (31/29%),ксеропетрофитов (8/6%), что свидетельствует орезкоконтинентальном климате Тувы.Мезофиты (17/16%), растения более умеренновлажных почв представлены средне.
В составепоясно-зональных подразделений залежнойфлоры выделяются следующиеэколого-фитоценотических группы:собственно-степные, лесостепные,горно-степные, пустынно-степные.
В целом, во флорепреобладают собственно-степная (47/42%)группа, что связано со сглаженностьюрельефа, отсутствием щебнисто-каменистыхсклонов. Иногда залежи размещаются насклонах увалов и шлейфах предгорий вмежгорных понижениях. Эти условияотличаются стабильным увлажнением,способствующим развитию видовмезоксеро-фитной экологии лугово-степной(40/37%) группы. Они имеют широкуюэкологическую амплитуду, могутпроизрастать в разных экологических нишах,что объясняет обширный ареал большинствавидов группы.
Наибольший интересвызывают горно-степная и пустынно-степнаягруппы. Горно-степной (20/18%) группехарактерны преобладание видов срозеточной формой роста с развитием мощныхмногоглавых корней. Они отличаются хорошосбалансированным режимом и сочетание ванатоматической структуре листьев ксеро- имезоморфных признаков (Горшкова, 1966).Пустынно-степные (43%) растения приурочены квыходам на поверхность карбонатных породили к сильно засоленным почвам по долинамстепных рек и озер (Пешкова, 2001). Анализпоказал, что главнейшее значение взалежной флоре Центральной Тувы имеютстепные, горно-степные и лугово-степныевиды при заметном снижениипустынно-степных.
5.3. Особенности флорызалежей как индикаторов в процессахдемутации растительности;типично-залежный, переходный и инвазионный(или случайно-залежный) элементы во флоре.
В настоящее время навсех континентах происходят интенсивныепроцессы синантропизации флоры ирастительности, в результате чегоадвентивные виды вытесняют аборигенные(Ульянова, 2005).
Под термином«адвентивная флора» мы, вслед за А.Я.Григорьев-ской, Е.А. Стародубцевой, Н.Ю.Хлызовым, В.А. Агафоновым (2004), М.М.Силантьевой (2006) понимаем гетерогенную попроисхождению и гетерохронную по временипроникновения группу видов в составерегиональной флоры, которая формируется врезультате трансконтинентальных,трансзональных и межзональных миграций,осуществляющихся благодаря прямому иликосвенному воздействию человека.
Адвентивная флора Тувыеще не была предметом специальныхисследований. В последние годы начатыработы по залежным экосистемам Тувы(Дубровский, Ооржак 2004). Расширение ареаласинантропных растений объясняетсяустранением конкуренции со сторонытуземных видов вследствие нарушенияестественных условий их произрастания илив связи с созданием искусственныхместообитаний. Первоначально занесенныерастения поселяются на нарушенныхчеловеком или природными факторами средыместообитаниях, в которых у нихотсутствует конкуренция с другими видами.Данную экологическую нишу в РеспубликеТыва освоили такие адвентивные виды как:Ceratocarpus arenarius, Cannabis ruderalis, Utricacannabina, U. dioica, Tribulus terrestris, Chenopodium aristatum, Salsolacollina, Artemisia jacutica, Neopalassia pectinata, Kochia scoparia,Convolvulus ammanii.
За счет заноса семян спосевным материалом сформировалась группарастений с широким ареалом (Куминова и др.,1985), обычные и для других районов Сибири:Setaria viridis, Avena fatua, Convolvulus arvensis,Chenopodium album, Axyris amaranthoides, Melandrium album, Brassica campestris,Thlapsi arvense, Camelina microcarpa, Neslia paniculata, Convolvulus fischerianus, Nonea pulla,Galeopsis bifida, Cirsium setosum, Sonchus arvensis.Все эти виды требуют значительнойвлажности почвы и свойственныпреимущественно предгорным районам.Исключение составляет Setariaviridis, встречающийсяповсеместно и в большом количестве.Расселение растений с целинных земель,особенно с эродированных участков снарушенным растительным покровом,способствовала выработке специфичных дляТувы комплекса адвентивных видов: Corispermum delicinatum, Ceratocarpus arenarius, Tribulus terrestris, Chenopodium aristatum,Lappula sguarrosa, Salsola collina, Kochia laniflora, Amaranthus blitoides,Eragrostis minor, Euphorbia humifusa, Artemisia jacutica, Neopalassiapectinata, Convolvulus ammanii. Особенночасто на легких песчаных почвах в массевстречается рогач песчаный (Ceratocarpus arenarius),проникающий с соседних участковзаброшенных пашен и обочин дорог иразрастающийся на полях особенно пышно.Таким же образом ведут себя якорцыстелющиеся (Tribulus terrestris). При улучшении водного режима иразвитии культурного растения эти сорнякине выносят конкуренции и из посевоввытесняются.
Кромевышеперечисленных видов часто на большихмассивах основной фон создают малолетниепрутняки - Kochia scoparia, K.siversiana, к ним примешиваютсяCorispermum delicatum,Agriophyllum sguarrosum, Artemisiajacutica, на отдельных участкахразрастается Scutellaria scordifoliaили Dracocephalumfoetidum.
Нами в ходе полевыхисследований и анализа полученныхрезультатов предложена систематизацияфлористического комплекса залежей (ФКЗ) натри группы с учетом их встречаемости иэкологических особенностей видоврастений:
А. Типичнозалежные растения (Setaria viridis, Chenopodium album, Camelina microcarpa, Lappulasquarrosa, Convolvulus arvensis, Nonnea pulla, Artemisia scoparia и другие);
Б. Залежно-степные (переходные)растения, одинакововстречающиеся как в степных сообществах,так и на залежах (Cleistogenes squarrosa,Psathyrostachys juncea, Carex duriuscula, Allium anisopodium, Potentillabifurca, Artemisia frigida и другие);
В. Спонтанно-инвазионные (случайнозалежные) растения, связанныес редкими случаями внедрения видов, несвойственных залежам (Stipacapillata, Carex pediformis, Dianthus versicolor, Potentilla nudicaulis,Astragalus adsurgens, Oxytropis strobilacea, Aster alpinus и другие).
Выявленные группы видовв определенной степени относятся киндикаторам состояния залежныхфитоценозов, по их относительному участиюможно прогнозировать не только возрастзалежей, но и позиции их в стадиивосстановления. Для анализа состояниязалежной растительности Тувы мы разделилитерриторию внутренней части региона на триприродных сектора. ЗападнаяТува (ЗТ) рассматриваетсяпреимущественного в границах Хемчикскойкотловины, Центральная Тува(ЦТ) включает Чаа-Хольскую,Улуг-Хемскую котловины с включениемТуранской депрессии; Восточная Тува (ВТ)охватывает восточные и юго-восточные частиЦентрально-Тувинской котловины свключением части предгорий нагорьяСангилен.
В результатесравнительно-сопоставительного анализа,доля всех трех компонентов ФКЗ оказаласьнаиболее высокой в ЦТ, соответственно А– 21,0%, Б – 6,5%, В – 9,6%. Более близкипоказатели по ЦТ и ВТ, в последней участиеслучайно-залежных видов наиболее низка(7,6%). Это указывает на низкую степеньинвазии коренных степных видов на залежи,по сравнению с ЗТ и ЦТ, что может бытьсвязана с одной стороны, с меньшимиплощадями залежей или с более лучшимиусловиями для богарного земледелия. Вболее аридных условиях ЗТ и ЦТ более 90%пахотных земель запущены в залежь ипоэтому естественна усиленная инвазиятипично степных видов.
В целом, выявленныегруппы видов относятся к четкиминдикаторам состояния залежныхфитоценозов, по их соотношению можнопрогнозировать в определенной степенивозраст залежей, а также стадии их развитияв демутационной динамике. Дальнейшийглубокий анализ залежных экосистем Тувыпозволит обосновать концепциюэкологического земледелия для аридных икрайне континентальных районов ВнутреннейАзии.
ГЛАВА IV. Степная изалежная растительность, характеристикаразнообразия и место их в системегеоботанического районированиярастительного покрова Тувы
При классификациистепной растительности Тувы мы,придерживаясь основныхпринципов Е.М. Лавренко (1940), следуем эколого-фитоценотическим построениямА.В.Куминовой (1960). Основные классы формаций(опустыненные, настоящие, луговые,высокогорные) степнойрастительности отличаются по структурефитоценозов, видовомусоставу, особенностям происхождения. Привыделении групп формацииучитывалась роль доминирующих жизненныхформ. В настоящих степяхгруппы формаций разграничиваются поопределяющей роли злаковойсинузии как наиболее важного и устойчивого компонента жизненнойформы (Ревердатто, 1928; Куминова идр., 1976). Вусловиях Тувы группа дерновинных злаковобогащается мелкимиперистыми ковылями (Stipa glareosa, S. orientalis), которые могутбыть охарактеризованы как доминантытипичныхопустыненно-степных сообществ.
Формации выделяютсяпо преобладающим видам основного ярусасообществ, ассоциации, как основные низшиетаксономические единицы -по доминантам основных ярусов, а также сучетом характерных видов. Каменистые, солонцеватые и песчаные степи рассматриваютсянами как эдафические (почвенно-грунтовые)варианты основных классовформаций.
Классификация степнойрастительности проводилась в тесной связис экологической ординацией сообществ,позволившей выявить важныеэколого-географические закономерностиформирования этих степей и датьэкологическую интерпретацию выделенныхсинтаксономических единиц (Кыргыс,Дубровский, Намзалов, 2006). На основаниианализа и обработки геоботаническихописаний с использованием пакетовпрограмм TURBOVEG и MegaTab, выделено 28 формаций(не включая серии дигрессионного ипетрофитного ряда сообществ), 13 группформаций, относящиеся к 4 классам формаций(или подтипам) степной растительности (см.ниже). При выделении низших единицрастительности – ассоциаций были использованырезультаты ординации фитоценозов в полеведущих экологических факторов - степениувлажнения и богатства почв сиспользованием методических подходов А.Ю.Королюка и др. (Экологическая оценка…,2005).Последние показаны на примере ординациисообществ настоящих степей (рис. 3, табл. 3).

Рис. 3. Экологическая ординация настоящихдерновинно-злаковых степей
Условныеобозначения. Ассоциации: 1–типчаково-мятликовые; 2 –крылово-ковыльные; 3 –караганово-мятликово-ковыльные; 4 –караганово-тонконогово-типчаковые
Таблица 3
Структура и видовойсостав ассоциаций настоящихдерновинно-злаковых степей(фрагмент)
| Класс формаций | настоящие дерновинно– злаковыестепи | ||||||||||||||||||
| Формации | полидоминантно-мелкодерновиннозлаковая | ||||||||||||||||||
| Ассоциации | 1 | 2 | 3 | 4 | |||||||||||||||
| номера описаний | 11 | 14 | 27 | 31 | 26 | 27 | 10 | 13 | 46 | 16 | 47 | 61 | 78 | 80 | 49 | 50 | |||
| число видов | 16 | 16 | 20 | 24 | 24 | 18 | 16 | 15 | 16 | 18 | 12 | 17 | 15 | 14 | 20 | 18 | |||
| проект.покрытие | 30 | 25 | 50 | 50 | 35 | 30 | 30 | 30 | 30 | 35 | 30 | 30 | 20 | 30 | 35 | 30 | |||
| Доминантныевиды | |||||||||||||||||||
| 1 | Stipa krylovii | 3 | 3 | 2 | 3 | 1 | + | 1 | 1 | 2 | 2 | 1 | 2 | 1 | |||||
| 2 | Poa attenuata | + | 1 | 1 | 1 | + | 2 | 2 | 1 | 2 | 2 | 2 | 2 | 2 | 2 | ||||
| 3 | Agropyroncristatum | 1 | 1 | 1 | 1 | 2 | 2 | 1 | + | + | + | 1 | 1 | 1 | 1 | ||||
| 4 | Koeleria cristata | + | 1 | + | 1 | 1 | 2 | 2 | 1 | 1 | 2 | + | 1 | + | 2 | 1 | 2 | ||
| 5 | Festuca valesiaca | 1 | 1 | 2 | 1 | 2 | 2 | 1 | 2 | 1 | + | 3 | 1 | 2 | 2 | ||||
| 6 | Caragana bungei | 3 | 3 | 3 | 3 | + | + | ||||||||||||
| 7 | Caragana pygmaea | + | 1 | 1 | 3 | 2 | 2 | 2 | |||||||||||
| Дифференциальныевиды | |||||||||||||||||||
| 8 | Oxytropiseriocarpa | 1 | + | 2 | 1 | ||||||||||||||
| 9 | Alliumvodopjanovae | 1 | 1 | + | + | ||||||||||||||
| 11 | Artemisia dolosa | + | 1 | 1 | |||||||||||||||
| 12 | Stipa orientalis | 1 | + | + | |||||||||||||||
| 13 | Schizonepeta multifida | + | 1 | 1 | 1 | ||||||||||||||
| 14 | Kochia prostrata | 1 | 1 | + | + | ||||||||||||||
| 15 | Thalictrum foetudum | 1 | + | 1 | 1 | ||||||||||||||
| 16 | Hedysarum gmelinii | 1 | + | 1 | |||||||||||||||
| 17 | Carex korshinskyi | 1 | 1 | 1 | 1 | ||||||||||||||
| Сопутствующиевиды | |||||||||||||||||||
| 19 | Coluria geoides | + | 1 | 1 | 2 | 1 | + | + | 1 | 1 | + | ||||||||
| 20 | Thymus mongolicus | 1 | 1 | 2 | 1 | 1 | 1 | + | + | 1 | + | 1 | |||||||
| 22 | Bupleurum scorzonerif. | + | 1 | 2 | 1 | 1 | 2 | + | 1 | + | 1 | 1 | 1 | ||||||
| Индифферентныевиды | |||||||||||||||||||
| 23 | Artemisia frigida | 1 | 1 | + | 1 | + | 1 | + | 1 | 1 | 1 | 1 | + | 1 | + | + | 1 | ||
| 24 | Potentilla acaulis | + | 1 | 1 | 1 | 1 | + | + | 1 | 1 | + | 1 | 1 | + | 1 | 1 | + | ||
| 25 | Carex duriuscula | + | 1 | 1 | + | 1 | + | 1 | + | 1 | 1 | 1 | 1 | 1 | 1 | + | 1 | ||
Примечание: Цифрами даются баллы проективногопокрытия: + –менее 1 %; 1 –от 1до 5 %; 2 –от 6 до 10%; 3 –от 11 до 15 %. ![]() – блокидифференциальных видов. Ассоциации: 1–крылоковыльные; 2 –караганово-тонконогово-типчаковые; 3–караганово-мятликово-ковыльные; 4 –типчаково-мятликовые
– блокидифференциальных видов. Ассоциации: 1–крылоковыльные; 2 –караганово-тонконогово-типчаковые; 3–караганово-мятликово-ковыльные; 4 –типчаково-мятликовые
Классификация степейТувы
Класс формаций - опустыненные степи. Группа формаций -дерновинно-злаковые опустыненные степиФормациягалечно-ковыльковая (Stipaglareosa) Группа формаций -полукустарничковые опустыненные степиФормации:нанофитоновая (Nanophyton erinaceum), прутняковая (Kochiaprostrata), терескеновая(Ceratoides papposa). Группа формаций - кустарниковыенастоящиестепи Формация -карагановая полынно-злаковая с караганойкарликовой (Stipa glareosa, Cleistogenes squarrosa, Agropyron cristatum, Artemisia frigida,Caragana pygmaea).
Серии петрофитныхгруппировок опустыненных степей:колючеостролодочниковая (Oxytropis tragacanthoides, О. aciphylla), дернистополынная (Artemisia caespitosa), курчавковаяпольшно-злаковая (Atraphaxis pungens, A. frutescens, Stipa orientalis, Agropyron cristatum, Artemisia santolinifolia), восточно-ковыльковая (Stipaorientalis)
Класс формаций - настоящие степи. Группа формаций – мелко-дерновинныестепи. Формации: мелкодерновиннозлаковаяполидоминантная (Poa attenuata, Festucavalesiaca, Stipa krylovii, Koeleria cristata, Agropyron cristatum), змеевковая (Cleistogenes squarrosa), крыловоковыльная(Stipa krylovii), житняковая (Agropyron cristatum), тонконоговая (Koeleria cristata), типчаковая (Festucavalesiaca).Группа формаций —крупнодерновинные степи. Формации: ковыльная(тырсовая) со Stipa capillata, овсецовая (Helictotrichon desertorum), стоповидноосоковая(Carex pediformis) Группа формаций—кустарниковые настоящие степи Формации — карагановая мелкодерновинная(Caragana pygmaea, С.bungei, Cleistogenes squarrosa, Stipa krylovii, Festucavalesiaca, Koeleria cristata), карагановая крупнодерновинная(Caragana bungei,С. pygmaea, Helictotrichon desertorum, Stipacapillata, Carex pediformis) Формации дигрессионного ряданастоящих степей: полынная(Artemisia frigida), бесстебельнолапчатковая (Potentilla acaulis), твердоватоосоковая(Carex duriuscula). Группаформаций - солонцеватыекрупно-дерновинно-корневищные степи Формации - чиевая (Achnatherum splendens), волоснецовая (Leymuspaboanus). Группа формаций -песчаные крупнодерновинно-корневищныестепи Формации: волоснецовая (Leymusracemosus), вейниковая (Calamagrostis epigeios), пырейная (Agropyron desertorum,A. michnoi)
Серии петрофитныхгруппировок настоящих степей: пырейная(Elytrigia gmelinii), коленчатопырейная (Elytrigia geniculata), арктогероновая (Arctogeron gramineum), хамеродосовая (Chamaerhodos ataica)
Класс формаций - луговые степи. Группаформаций - осоково-злаковые луговые степи Формация:злаковая (Phleumphleoides, Bromopsis pumpelliana, Poa stepposa, Helictotrichon schellianum) Группа формаций - разнотравныелуговыестепи Формация -ирисовая (Irisruthenica), простреловая(Pulsatilla patens)
Серии петрофитныхгруппировок луговых степей: плаунковые(Selaginella sanguinolenta) мезопетрофитные сообщества,злаково-разнотравные закустаренныемезопетрофитные сообщества (Spiraeamedia, Pentaphylloides fruticosa, Cotoneaster melanocarpus, Phleum phleoides, Helictotrichon schellianum, Poa attenuata, Carex pedijormis, Bupleurum multinerve).
Класс формаций - высокогорные криоксерофильныестепи. Группа формаций - мелкозлаковыекриоксерофильные степи Формации: чуйскотипчаковая (Festucatschujensis),ленскотипчаковая (Festucalenensis). Группаформаций — кобрезиевыекриоксерофильные степи.Формация -нителистнокобрезиевая(Kobresia filifolia.). Группа формаций -разнотравные криоксерофильные степи.Формация -злаково-разнотравная (Eremogone formosa, Potentillasericea, Oxytropis macrosema, Artemisia depauperata, Festuca kryloviana,F. lenensis, Poa attenuata).
Серии петрофитныхгруппировок высокогорных степей:разнотравно-осоково-злаковая (Poaattenuata, Koeleria altaica, Carex rupestris, Potentilla sericea), трагакантово-остролодочниковая(Oxytropistragacanthoides,Smelovskia calycina).
Класс формаций - Настоящие дерновинно-злаковыестепи. Настоящие степиобъединяют растительные сообщества,эдификаторную роль вкоторых играют ксерофитные дерновинныезлаки. Среди них наиболееобычны типчак, тонконог, житняк, овсец,ковыли, волоснецы. Эти степив Туве получили широкое распространение, занимая 2209,4тыс. га. (Ершова, Намзалов, 1985). В котловинахи межгорных депрессиях на выровненных местообитанияхони занимают огромные площади, формируяустойчивые зональные сообщества. В горныхрайонах настоящие степипредставлены исключительно петрофитнымивариантами или такназываемыми каменистыми степями. В зависимости от экологическихусловий местообитаний в Туве отмеченыварианты настоящих степей:псаммофитные,господствующие на развеваемых песках игалофитные – на солонцеватыхтеррасах долин рек и в приозерныхкотловинах.
По видовому составуформации настоящих степей характеризуютсянезначительным разнообразием и заметнообеднены по сравнению сдругими островными степями Сибири, на чторанее указывали К.А. Соболевская (1950, 1958)и А.В. Калинина (1957). В экологическом спектре преобладаютксерофиты (до 80%), по характеру ареалов ведущее место занимаютсевероазиатские и евразийские виды (50%).Формационный состав этих степейразнообразен и представлен16 формациями.
Среди формацийнастоящих степей широко распространенымелкодерновиннозлаковые полидоминантныев обширных межгорных котловинах и насклоновых местообитаниях, характерны нагорных каштановых малоразвитых ищебнистых почвах. Основныеэдификаторы: Poa attenuata, Festucavalesiaca, Stipa krylovii, Koeleria cristata, Agropyron cristatum. Среди соэдификаторов наиболееобычна Caragana pygmaea, а в лесостепном поясе – C.bungei. Для сообществ этойформации, не нарушенных интенсивнымвыпасом, устойчивая полидоминантностьзлаковой основы травостоя. Выделеныассоциации: типчаково-мятликовые (Poa attenuata, Festuca valesiaca);крыловоковыльные (Stipakrylovii);караганово-мятликово-ковыльные (Caragana pygmaea, Poa attenuata, Stipa krylovii); караганово-тонконогово-типчаковые(Caragana bungei,Festuca valesiaca, Koeleria cristata).
По результатамэкологической ординации сообществнастоящих дерновинно-злаковых степейвидно, что ассоциациякараганово-тонконогово-типчаковыезанимают влажные местообитания (51-56), атипчаково-мятликовые и крыловоковыльныерасполагаются в сухихместообитаниях (42-48), промежуточное положениезанимают караганово-мятликово-ковыльные. Полученные в ходе ординациикластеры сообществ диагностируют синтаксоны рангаассоциаций (табл. 7).
В процессеинтенсивного и длительного использованиязлаковых мелкодерновинныхстепей под выпас скота происходятразрушение злаковой основы,нарушение общей структуры и деградациятравостоя.
Крупнодерновинныестепи занимают в Туве меньшие площади, чеммелкодерновинные, и сосредоточены восновном по окраинамкотловин и низкогорным элементамрельефа. В качествеценозообразователей выступают ковыли,овсец пустынный, осокастоповидная и другие виды.
Кустарниковыенастоящие степи представлены в Туве формациями: караганниковаямелкодерновинная икараганниковая крупнодерновинная.Эдификаторную роль в этих степях играют два вида караган:Caragana pygmaea и С. bungei. Караганниковые мелкодерновинныестепи встречаются в пределах степногопояса и лишь небольшими островками поюжным склонам проникают впояс лесостепи. Фитоценозы этой формации приурочены к щебнистымпочвам выровненных местообитаний. В травостое доминируютKoeleria cristata, Cleistogenes squarrosa, Stipakrylovii.
Класс формаций – Опустыненные степишироко распространены вТуве, сосредоточены впределах Тувинской и Убсунурской котловинпо террасам рек, пологимшлейфам предгорий, каменистым южнымсклонам мелкосопочников.Это крайне аридные сообщества в условияхстепного пояса,характеризующиеся низкотравным,обедненным разреженнымтравостоем. В экологическом спектрехарактерно господствоксеропетрофитов (до 70%). Географическийанализ флоры показал явноепреобладание центрально-азиатских видов (до 45%).
Группа формацийдерновинно-злаковых опустыненных степейкак по структуре, так и поосновным компонентам составляющих их растений, наиболее близка кзональным пустынным степямсеверогобийского типа, впервые описаннымА.А. Юнатовым (1974). В составе формацийвыделяются галечноковыльковые инанофитоновые степи. В Туве Nanophyton erinaceum (джунгаро-туранский вид) в рангеэдификатора имеет свой северо-восточный пределраспространения.
Класс формаций -Луговые степи. Луговые степи - наиболеемезофильный вариант степной растительности в условиях сухого ирезко континентального климата Тувы— занимаютплощадь 244,5 тыс. га. Они отмечены посклоновым микропонижениям,впадинам между холмами и нижним частямсклонов северныхэкспозиций на хребтах Западный Саян,Уюкский, Шапшальский,Цаган-Шибэту. Флористический состав луговых степей значительноразнообразнее, чем настоящих степей,и беднее (заметнопреобладают растения-петрофиты), чем вподобных формациях луговыхстепей Хакасии (Куминова и др., 1976), Алтая(Куминова, 1960), Забайкалья(Рещиков, 1961; Пешкова, 1972). Втравостое луговых степей в значительномобилии выступают как дерновинные (Роа attenuata, Helictotrichon desertorum), так и корневищные ксеромезофильныезлаки (Phleum phleoides, Helictotrichon schellianum, Poapratensis, Bromopsis inermis), высок процент осок(Carex pediformis, С. kirilowii, С. supina, С. obtusata); менее разнообразна группа бобовых(Onobrychis tanaitica, Oxytropis strobilacea, Astragalus adsurgens). Красочный аспектлуговым степям придаетдовольно богатая группа разнотравья– Iris ruthenica, Aconitum barbatum, Pulsatilla patens, Bupleurum multinerve, Galium verum. Основу экологического спектратравостоя луговых степей составляет группа мезоксерофитов(40-50%). Погеографическому распространению преобладаютевразийские и североазиатские виды (25-35%). Фитоценотический спектрлуговых степей представлен злаковой, ирисовой и простреловойформациями.
Класс формаций – Высокогорные криофитные степи.На высоких выположенныхводоразделах (от 2000 до 2500 м над ур. м.) по склонам Монгун-Тайги июжным макросклонам хрзб-товЦаган-Шибэту, Западный и ВосточныйТанну-Ола, нагорья Сангилен в особыхкриофитно-ксерофитных условиях,создающихся в полосеконтакта тундровых и горно-степныхландшафтов, формируются своеобразные сообщества—высокогорные степи.
Вопросытаксономического ранга высокогорныхстепей и место их в типестепной растительности интересовалимногих исследователей.П.Н. Овчинников (1947,1948) в высокогорнойрастительности СреднейАзии выделяет подтип - криофитные степи.А.А. Юнатов (1948,1950),характеризуя степи Монголии, обосновываетособую группу степей сдоминированием высокогорных овсяниц икобрезий.
Сообществавысокогорных степей мыотносим к классу формаций (подтипу) криофильных степей. Выделенычетыре формации: мелкозлаковые (чуйскотипчаковая иленскотипчаковая), кобрезиевая и разнотравная(Намзалов, Дубровский,2007а).
4.2. Залежнаярастительность и процессы их зацелинения,основные стадии демутации
В условиях резкоконтинентального и аридного климата Тывыразвитие богарного земледелия себя неоправдала. Интерес к залежным системамособенно усилился в 90-ых годах ХХ столетия,в связи с развитием идейэколого-адаптивного природопользования(Помишин, 1993, Намзалов, Доржиев, 1999, Аракчаа,2003).
Для растительностизалежей Тувы характернопятнисто-мозаичное сложение, в структуресообществ которой трудно уловить иходновозрастность с позиций общепринятойконцепции 4-ех стадийности в залежнойдемутации. Парадоксальнаяразнонаправленная векторностьдемутационного процесса по-видимомуявляется следствием не толькомикроклиматических контрастов, но иисключительной неоднородности субстрата,механического, термического игидрологического режимов поверхностныхгоризонтов почв. Виды растений различнойэкологии, обеспечивающие первичные стадиизалежной сукцессии, оказываются в условияхдостаточно неоднородных по эдафическомуфону (рис. 4).
На представленномтрансекте были зафиксированы ценозы какбурьянистой, так и корневищной стадий сучастием плотнокустовых злаков. В процесседемутации растительности в условияхгорносклонового и контрастного поэкологическим условиям богарногоземледелия в Тыве и Забайкалье имеютразличные временные (по протяженности) иколичественные (по этапам восстановления)стадии.
Демутация как сложныйпроцесс восстановления не толькоестественной растительности, но ипочвенного плодородия после определенногопериода ее освоения под посевы культурныхрастений изучены в различных секторахЕвразийского пространства (Голубинцева,1930; Глумов,1953; Микляева,1996; Быков и др.,2003).Многими исследователями подчеркивается,что наличие общих черт в динамикевосстановления не исключает проявленияряда региональных особенностей. Последниеявляется следствием не толькосамобытности длительного этапафилоценогенеза растительности, но иприродно-климатических условийсовременности. В результате, в демутацияхмогут различаться количество временныхстадий и их продолжительность, состав иразнообразие видов растений в сообществах.Нередко могут выпадать отдельные стадииили напротив, могут усложняться за счетформирования особых промежуточныхстадий.

Рис. 4. Карта-схема трансекты залежнойрастительности Улуг-Хемской котловины (вокрестностях с. Арыг-Узуу), июль 2004 г.
Классикамистепеведения (Залесский, 1918; Лавренко, 1940;Семенова-Тян-Шанская, 1966) даны обобщающиесхемы зацелинения степных залежей, они вцелом состоят из 4 стадий: 1. Бурьянистаястадия (стадия полевых сорняков); 2.Корневищная стадия; 3. Стадия рыхлокустовыхзлаков; 4. Стадия плотнокустовых злаков(вторичная целина).
Сукцессия на залежныхземлях на месте сухих степей в условияхгорно-котловинного рельефа Тувы имеет своюспецифику и часто не укладывается вобщепринятую схему, установленную вевропейской части страны. Касаясьрегиональных особенностей демутационныхпроцессов, следует отметить факт развитияособой мелкобурьянистой стадии,предшествующей развитию крупного бурьяна,в условиях Абаканских и Тувинских степей(Голубинцева, 1930; Дымина, 1983; Куминова, 1985;Дубровский и др., 2007). С другой стороны, назалежах по фону каштановых почв в Бурятиинаблюдаются случаи выпадения бурьянистыхгруппировок (Быков и др., 2003), при этомдлиннокорневищные злаки (Elytrigia repens, Leymus chinensis)формируют монодоминантные сообщества снебольшим участием видов-монокарпиковпервичной стадии, что свойственна и вусловиях Тувы.
Мелкобурьянистаястадия (описание 21) – Основным компонентом травостояявляются однолетние растения, именно ониявляются пионерами зарастания.Доминантами являются: Setariaviridis, Artemisiaglauca, Hypecoum erectum,Субдоминант – Convolvulus arvensis, Potentilla bifurca.
Бурьянистая стадия(описание 24) –Доминанты одно-двулетние виды семействаAsteraceae: Artemisia scoparia, Artemisia sieversiana, Artemisia obtusiloba, Artemisia glauca.Субдоминантные виды: Medicago falcata, Heteropappus altaicus, Lappula microcarpa.Характерным признаком этой стадииявляется преобладание разнотравья.Структура травостоя трехъярусная: I ярус -Artemisia siversiana, Artemisia scoparia, II ярус –Heteropappus altaicus, IIIярус – Convolvulus arvensis, Linaria vulgaris.По жизненным формам преобладаютстержнекорневые растения. В практическихцелях залежи с этой стадией неиспользуются, так как имеют низкуюхозяйственную оценку или низкийкормозапас. Кормовое достоинствобурьянистых залежей низкое.
Корневищная стадия(описание 7) –Отмечается незначительноеувеличение сорных видов. I ярус представлен - Artemisia scoparia, II ярус– Elytrigia repens, III ярус– Convolvulus arvensis, Astragalusadsurgens, Medicagofalcata, Melilotus albus, Vicia cracca, Heteropappusaltaicus. Доминантом этой стадииявляется Elytrigia repens. На этой стадии можно наблюдатьобразование большого количества пятенпырея, которые постепенно расширяются ивытесняют стержнекорневые растения.Залежи этой стадии используются длясенокошения, где в большей степенивстречаются растения из семейства бобовых.Однако качество сена среднее, из-заприсутствия сорных и ядовитыхрастений.
Стадия рыхлокустовыхзлаков (описание 16) – Ярусность слабовыражена. Доминанты – Elytrigia repens, Convolvulus arvensis, Medicago falcate, Artemisiascoparia. Вариантстаровозрастной залежи на ранних стадияхзацелинения плотнокустовыми злаками, поокраине основной пахотной полосы. Этизалежи местное население в основномиспользуют для сенокошения. Качество сенахорошее.
Стадия плотнокустовыхзлаков (описание 15) – Ярусность слабовыражена. Доминантами являются Carex pediformis, Cleistogenessguarrosa, Stipa capillata, Gallum verum, Heteropappus altaicus. Послезалежная целина.Крупнокрутинная от дерновинок ковылейзалежь – стадиязацелинения плотнокустовыми злаками.Сообществ, находящихся на данной стадиидемутации отмечено нами в незначительномколичестве, одна из ключевых точек – в окрестности селаАрыг-Узуу.
Схемы зарастаниязалежей на каштановых и светлокаштановыхпочвах Центральной Тувы (рис. 5).
| НА КАШТАНОВЫХ ЛЕГКО СУГЛИНИСТЫХПОЧВАХ | НАСВЕТЛОКАШТАНОВЫХ СУ- ПЕСЧАНЫХ ПОЧВАХ |
| I | Липучково-веничнополынная (Lappula squarrosa + Artemisia scoparia) | Полынно- коноплевая (Artemisia obtusiloba + Artemisia scoparia - Cannabisruderalis) | Неопалласиево-рогачевая (Convolvulus ammanii – Neopallassia pectinata +Ceratocarpus arenarius) |
| II | Вьюнково-пырейная(Convolvulus arvensis + Elytrigiarepens) | Гетеропаппусово-вьюнковая (Artemisia sieversiana +Heteropappus altaicus + Convolvulus ammanii) |
| III | Пырейно-змеевково-разнотравная (Elytrigia repens + Cleistogenessquarrosa + variaherbosa [Potentilla bifurca, Artemisia frigida, Alliumanisopodium]) | Полынно-вильчатолапчатково-змеевковая (Artemisia glauca, A. frigida +Potentilla difurca + Cleistogenes squarrosa) |
| Гетеропаппусово-лапчатково-ковыльная (Heteropappus altaicus + Potentilla bifurca – P. acaulis +Stipacapillata) |
IV
Рис. 5. Схемы зарастания залежей
Примечание к схемедемутации
| I -IV - | стадии залежной сукцессии(зацелинения) |
| ______- | векторы направленностисукцессии |
| - - -- - - | вектор вероятностного направлениясукцессии (не всегда наблюдаемая сменасообществ) |
| Квадрат с жирнойлинией - | заключительная стадия сукцессии(вторичная целина) |
| Квадратс тонкими линиями- | условно-заключительная стадия назалежах по фону светлокаштановыхсупесчаных почв |
Так, по результатамнаших исследований и данным И.П. Быкова поБурятии (2003) на залежах по фону каштановыхпочв наблюдаются случаи выпадениябурьянистых группировок, при этомдлиннокорневищные злаки (Elytrigia repens, Leymus chinensis)формируют почти монодоминантныесообщества с небольшим участием видов -монокарпиков первичной стадии. Нами в ходеполевых, маршрутно-рекогносцировочныхисследований залежной растительности былинеоднократно описаны сообщества различныхстадий в границах одновозрастной пахотнойполосы, запущенной в режим демутации.
На рыхлокустовойстадии, вместо пырея появляются рыхло-иплотнокустовые злаки и сопутствующие имвиды (Stipa capillata, Cleistogenes squarrosa,Agropyron cristatum, Convolvulus ammanii, Рotentilla bifurca и др.), болееприспособленные к создавшимся новымусловиям почвенной среды. Процесснакопления органического веществапроходит замедленным темпом, чем напредыдущей стадии и может длиться напротяжении 10-15 лет. В этот период временипроисходит перевод всего запаса элементовпочвенной пищи в органическую форму,недоступную для усвоения еерыхлокустовыми злаками (Намзалов и др.,1998;Быков, 2000).
Таким образом,демутация растительности на залежахпоказывает происходящие изменения вструктуре ценозов, где на начальнойбурьянистой стадии преобладаютстержнекорневые малолетники – полыни и конопля. В дальнейшем идетнасыщение ценозов видами коротко- идлиннокорневищных форм. Наблюдаетсяобразование большого количестварыхлокустовых и плотнокустовых злаков,которые постепенно расширяются ивытесняют растения не приспособленных ксухим условиям среды. Комплексное изучениеэкологии залежных земель, еерастительности имеет не только большоетеоретическое значение, но и практическое.
4.3. Фитогеографическоерайонирование важнейших степных массивовТувы (Монгун-Тайгинскаявысокогорно-степная, Центрально-Тувинскаясухостепная, Убсунурскаяопустыненно-степная) и ихособенности
Выделение территорийразных рангов, растительность которыхсоставлена неоднородными структурнымитипами, является целью геоботаническогорайонирования (Сочава, 1952). Проводится онопо признаку экологической связирастительности с геофизической средой и сучетом современной динамики растительныхсообществ (Сочава,1979). Впервые дробноеботанико-географическое районированиеТувы сделано К.А. Соболевской (1950).Используя положения и принципырайонирования, высказанные В.Б. Сочавой (1979)и А.В. Куминовой (1960,1971), и составленныекрупно- и среднемасштабные картырастительности, на территории Тувы намивыделено пять провинций и двенадцатьокругов (рис. 6). Две южные провинции и дваокруга в них продолжаются в Монголии.Большая часть территории Тывы находится впределах Алтае-Саянской горной области изахватывает узкую полосу на северо-западеЦентральной Азии (Маскаев, Намзалов,Седельников, 1985).
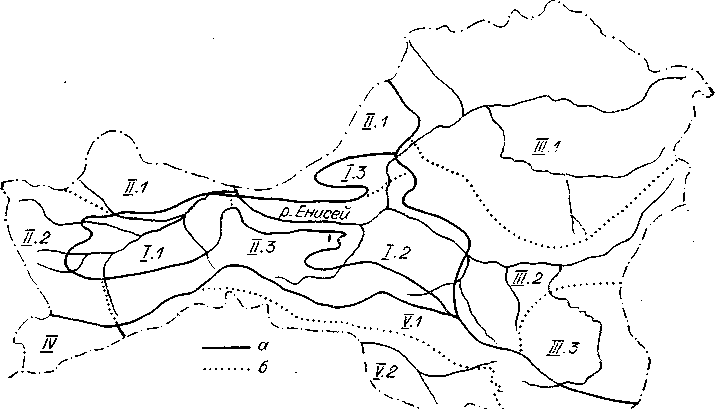
Рис. 6.Схема геоботаническогорайонирования Тувы (по Маскаеву, Намзалову,Седельникову, 1985).
I— Тувинская котловинная степнаяпровинция, округ: 1.1 — Хемчикскийсухостепной, 1.2 —Центральнотувинский лугово-степной,1.3 — Турано-Уюкский лиственничныйлугово-степной; II — Западно-Саянскаягорная таежная провинция, округ: II.1 — Куртушибинский остепненныйкедрово-лиственничный, II.2—Верхнехемчикский лиственничный остепненный горно-тундровый, II.3 — Таннуольский тундровый,остепненный лиственничный; 111 —Восточно-Саянская горная таежнаяпровинция, округ: 111.1 — Тоджинскийкедрово-лиственничный,111.2 — Восточно-Тувинскийгорно-тундровый лиственничный, III.3 — Сангиленскийлиственничный горно-тундровый; IV — Юго-Восточная Алтайская горнаястепнаяпровинция, Монгун-Тайгинскийгорно-тундровый опустыненный округ; V—Убсунурская равниннаяопустыненно-степная провинция, округ:V.1 —Южно-Таннуольско-Сан-гиленский степной, V.2 — Убсунурскийопустыненный степной.Границы: а — провинций, б —округов.
Степи – одна из важнейших ишироко распространенных типоврастительности в Туве, онидифференцируются на три крупныхрегиональных варианта. Крупнейшая из них– степиЦентрально-Тувинской котловины, степиюжных предгорий Танну-Ола и нагорьяСангилен составляют лишь северные окраиныпустынной Убсунурской депрессии и степноймассив горного узла Монгун-Тайга наюго-западе территории, в приалтайскойчасти Тувы.
Ниже подробнорассмотрим особенности этих трех степныхпровинций региона.
Тувинская котловиннаястепная провинция. В неевходят обширные пространства Хемчикской и Улуг-Хемскойкотловин, нескольконебольших периферических межгорныхдепрессий (Туранская,Уюкская, Эдыгейская, Чаа-Хольская) ипредгорья хребтов Танну-Ола,Шапшальского, Западного Саяна. Днища котловин покрываюткаштановые почвы, чаще всего их петрофильные варианты. В рядугорно-степных почв господствуют горные каштановые почвы, обыкновенныеи южные черноземы, нередкомалоразвитые и щебнистые.
Господствуют степи (50%от общей площади провинции). Они представлены разреженными инизкотравными опустыненными группировками - ковыльковыми (Stipaglareosa), полынно-змеевковыми (Cleistogenes squarrosa, Stipaglareosa), полынно-змеевково-ковыльковыми (с караганой карликовой) иковыльково-нанофитоновыми, а также сухими настоящимидерновинно-злаковыми степями предгорий (ковыльные с ковылем Крылова,житняковые, тонконогово-типчаковые с караганой Бунге).Небольшие массивы песчаныхзлаковых степей локализованы в восточной части котловины, вХадынской озерной депрессии и в междуречье Чаргык — Шеми в Хемчикскойкотловине.
Луговые угодьязанимают 15,2% и представлены пойменными злаковыми (ячменевые,полевицевые, бескильницевые),низинно-заболоченными осоковыми (Carexenervis, С.caespitosa, С.dichroa), вейниковыми (Calamagrostis neglecia) лугами. Лесапредставлены долинной тополевой формациейс участием лиственницысибирской, березы мелколистной. Общиеплощади ихнезначительны. Тувинскаякотловина - район интенсивногохозяйственного освоения.Здесь сосредоточены основные массивысенокосных угодий, пастбищи пахотных земель.
На территориипровинции выделено три округа: Хемчикскийсухостепной,Центральнотувинский лугово-степной иУюкский лиственничныйлугово-степной. Хемчикскийсухостепной округ находится в пределах Хемчикскойкотловины. С восточной стороны округ отделяется от Улуг-Хемскойкотловины средневысотным хр. Адар-Таш.
Юго-ВосточнаяАлтайская горная степная провинция расположена вюго-западной части Тувы и отгорожена отостальной части республикина севере хр. Цаган-Шибэту.В провинции выделен одинокруг - Монгун-Тайгинский горнотундровый опустыненный (377,3 тыс.га). Он соприкасается с Чуйским высокогорно-степным округомподпровинции Юго-Восточный Алтай, по схеме районированияАлтая А.В.Куминовой (1960). Округ являетсянаиболее высокогорной территорией в Туве.Его низшие отметки лежат навысотах около 1900 м, а высшие достигают 3970 м (гора Монгун-Тайга). Врельефе ярко выражено сочетание альпинотипных и гольцовых форм,что обусловливает большую комплексность в растительномпокрове.
Растительный покровокруга характеризуется господствомвысокогорнойрастительности и почти отсутствием лесов,занимающих всего 1 тыс. га(рис. 7). Высокогорнаярастительность (с высот более 2300 м)занимает более половинытерритории округа. Господствуют дриадовые,лишайниковые, овсяницевые икобрезиевые тундры. Распределение их по территорииокруга неравномерно. Степнаярастительность на территории округазанимает высоты ниже 2100 м,формируяхорошо выраженный пояс. Господствующее положение в нем имеютопустыненные и каменистые степи, приуроченные к склонам южнойэкспозиции со сдуваемым в зимний периодснегом. В местах интенсивного выпасаформируются злаково-лапчатковые,холоднополынные, чиево-холоднополынныестепи. Опустыненные и каменистые степныесообщества Монгун-Тайгинского округа аналогичны подобнымсообществам Юго-Восточного Алтая и Северной Монголии. По севернымсклонам распространены настоящие илуговые (в местообитаниях свыраженным снеговым покровом) степи,представленныекрупнодерновинными тырсовыми ценозами,мелкодерновинными,полидоминантными, злаково-осочковымимелкодерновинными сообществами настоящих степей.Среди луговых степей преобладают разнотравные, разнотравно-злаковые,злаково-осоковые сообщества. По долинамрек в степном поясе небольшими участкамираспространены злаковыеостепненные луга и различные ихварианты. Леснаярастительность представленалиственничными и тополевыми лесами, встречающимисяотдельными участками в наиболее теплых долинах рек.

Рис. 7.Фрагмент картырастительности Монгун-Тайгинского округа (М 1: 100 000).
Условныеобозначения. Тундры: 1 - Щебнистыелишайниковые и криофитно-разнотравныетундры; 2 - Лишайниково-дриадовые тундры; 3 -Травянистые тундры с участием овсяниц,осок и кобрезий; 4 - Ерниковые,рододендроновые тундры; Леса: 5 - Высокогорныелиственничные редколесья; 6 - Лиственничныекустарниково-травяные; Кустарниковые заросли: 7 - Ерниковые;Степи: 8 - Высокогорныекриофитные степи (крыловотипчаковые,мятликовые, кобрезиевые); 9 -Холоднополынно-змеевковые опустыненныестепи; Луга: 10- Альпийские разнотравные луга; 11 -Субальпийские разнотравно-злаковые луга; 12- Осоковые заболоченные высокогорные луга;Растительность скал иосыпей: 13, 14 - Петрофитныегруппировки и единичные поселенцы наскалах; 15 - Растительные группировки нагалечных наносах рек. 16 - Ледники.
В хозяйственномотношении округ являетсяживотноводческим, пашнизанимают немногим более 300 га. Основноенаправление животноводства–овцеводство, в значительной мере – крупный рогатый скот (яки). Пастбищарасположены как в степном, так и ввысокогорном поясе, гдевыпасают овец и яков. В целом растительныйпокров используется подвыпас относительно равномерно. Пастбищная перегрузка наблюдаетсятолько вблизи населенных пунктов, около стоянок чабанов и подолинам рек Каргы и Моген-Бурен.
Убсунурская равниннаяопустыненно-степная провинция занимает южныемакросклоны хребтов Западный и ВосточныйТанну-Ола, юго-западнуюпокатость нагорья Сангилен и севернуючасть Убсунурскойкотловины. Южная границапровинции совпадает с государственнойграницей СССР, северная -проходит по водораздельной части хребтовЗападный и ВосточныйТанну-Ола, восточная - по водоразделу рекТес-Хем и Балыктыг-Хем,западная - по р. Барлык.
В растительномпокрове хорошо выражена высотнаяпоясность. Впровинции выделяются два округа:Убсунурский опустыненно-степной и Южно-Таннуольско-Сангиленскийстепной.
Растительность округахарактеризуется господствомопустыненных степей, а попериферии, в границах нижней частигорно-степного пояса, ониприобретают характер сухих настоящих,дерновинно-злаковых степей.Полосу опустыненных степейформируют ковыльковые, змеевково-ковыльковые, нанофитоновые,ковыльково-нанофитоновые степи,занимающие обширные низкогорные части отприозерных равнин допредгорных шлейфов хребтов Танну-Ола. Наболее легких почвах шлейфовв водораздельных пологовсхолмленныхравнинах междуречий(Холь-Ежу, Арасканниг-Хем, Улуг-Оруг и др.)широко распространенызмеевковые степи, поднимающиеся здесь довысоты 1700 м (Ханминчун, 1980). Внаиболее пониженных притеррасных частях отмечены пустынныековыльковые степи, часто закустаренныекараганойкарликовой. Начиная с высоты 1300-1400 м на склонахразличных экспозицийгосподствуют различные варианты горныхпетрофитных степей. Склоныюжных экспозиций носят более опустыненныйхарактер, среди разнообразиясоставляющих их фитоценозов выделяютсяполынно-ковыльковые,разнотравно-злаковые закустаренные,разнонотравно-ковыльковые степи. Насклонах северных ориентации распространены более мезофитныесообщества: осоково-злаковые, стоповидноосоковые,разнотравно-мелкозлаковые степи. Вгорно-степном поясе навысотах 1200-1600 м выделяются плаунковыефитоценозы.
Леса занимаютнезначительные площади и представленыдолинными тополевыми лесами иногда сучастием березы мелколистной и некоторых видов ив.
В хозяйственномотношении провинция имеетживотноводческое направление,преимущественно развито овцеводство икозоводство. Каменистыезлаковые степи по горным склонамиспользуются как естественные пастбища. Обширныеопустыненные степи на шлейфах и подгорных равнинах являютсяпозднелетними и осенними пастбищами. Значительная часть ихиспользуется зимой. Существенное значение имеют долинные злаковыелуга. Они высокоурожайны, хорошего качества, но занимаютнебольшую площадь. Здесь сосредоточены основные сенокосныеугодья.
ГЛАВА VI. Продуктивностьи деструкция растительного опада взалежных сообществах
Восстановлениерастительности на залежах протекает потипу вторичных сукцессий, относящихся ксингенетическим сменам растительности(Зайченко, Хакимзянова, 1999; Титлянова,Афанасьева, Наумова, 1993). Изучениеособенностей смены фитоценозовпроводились на многолетних залежах (5-10 иболее лет) в Центрально-Тувинскойкотловине с 2004-2007 гг. Работа по динамикепродуктивности залежных фитоценозовпроводилась в окрестности озера ЧедерКызылского кожууна на пробных площадках 1м2. Дляисследования были выбраны сообщества постадиям их восстановления: 1.Полынно-коноплевая залежь (бурьянистаястадия); 2. Пырейно-гетеропаппусовая залежь(корневищная стадия); 3.Гетеропаппусово-змеевковая залежь (стадиярыхлокустовых злаков).
Общие запасы живойфитомассы в полынно-коноплевом сообществев среднем за 2005 год составило 11,07 ц/га, а 2006год – 11,9 ц/га.Повышение надземной биомассы уполынно-коноплевого сообществанаблюдается с июня по июль месяцы. Потомидет постепенное снижение урожайности.Запасы живой фитомассыпырейно-гетеропаппусово-вьюнковогосообщества за 2005 год составило 9,09 ц/га, а за2006 равно 11,01 ц/га. Повышение продуктивностиэтого участка наблюдается с июля по августмесяцы. Затем идет постепенное снижениеурожайности (Ооржак, 2007). Общие запасы живойфитомассы в гетеропаппусово-змеевковомсообществе в среднем за 2005 год составило 11,5ц/га, а 2006 год –12,05 ц/га. Для этой залежи характерноеувеличение продуктивности с июля посентябрь месяцы.
Продуктивность в целом,на каштановых легкосуглинистых почвах,мало меняется и варьирует в пределах от 4,0до 7,0 ц/га, однако качественный составтравостоя существенно изменился.Наибольшая продуктивность надземнойфитомассы характерна длягетеропаппусово-змеевкового сообщества в2005-2006 годы, наименьшая - впырейно-гетеропаппусовом сообществе. Пообщему запасу фитомассы все изученныезалежи согласно десятибалльной шкалеН.И.Базилевич и Л.Е. Родина (1964), можноохарактеризовать как оченьмалопродуктивные (2 балла – общее количествобиомассы от 25-50 ц/га).
В качестве основногокритерия интенсивности разложениярастительного материала нами была принятаубыль массы образцов за определенныйпериод. Скорость разложения растительныхостатков определяется химическим составомразлагающегося материала. Высокоесодержание легкорастворимых органическихсоединений благоприятствуют быстройминерализации растительного опада. Убыльмассы происходит за счет вымывания ивыщелачивания водорастворимых веществ иих последующей утилизациимикроорганизмами. Растительные остаткипырейно-гетеропаппусовом сообществе (70% загод), наиболее богаты водорастворимымиорганическими веществами и зольнымиэлементами. А минимальная потеря массынаблюдается у полынно-коноплевогосообщества (50,4 % за год).
Интенсивностьразложения модельных субстратов: белка ицеллюлозы проводились с целью оценкиэкологических условий разложенияорганического вещества в различныхприродных условиях. Интенсивностьразложения белка и целлюлозы колеблется взависимости от времен года. В теплыйпериод, т.е. в мае разложение целлюлозырезко увеличивается в полынно-коноплевомсообществе. В августе с увеличениеммикробного пула целлюлолитиков скоростьразложения целлюлозы повышается до 0,12-0,13%.С понижением температуры окружающей средыинтенсивность процесса падает 0,08-0,09%, а всутки 0,003%.
Преобладающей группоймикроорганизмов в микробном сообществерастительного опада являются бактерии.Большую роль в разложении растительногоопада играет функциональная группабактерий –гидролитики. В пробах растительного опадабыли изучены различные физиологическиегруппы бактерий: сапрофиты (аэробы ианаэробы), протеолитики – участвующие вразложении белка (аэробы и анаэробы),амилолитики-крахмалразрушающие,целлюлолитики – целлюлозоразрушающие (аэробы ианаэробы).
В микрофлоре висследуемых объектах преобладающейгруппой микроорганизмов были сапрофиты.Максимальная численность сапрофитов(аэробов и анаэробов) в растительных опадахобнаружена в осенний период во всехисследуемых залежных фитосистемах.
Численностьмикроорганизмов подвержена сезоннойдинамике. В аэробных условиях разложениецеллюлозы ведут микроорганизмы разныхтаксономических групп: бактерии, грибы,актиномицеты. Наиболее ярко это выражено уцеллюлозоразлагающих бактерий (ЦРБ). Втечение года наблюдается рост численностиЦРБ – в начале иконце лета. Наиболее активное разрушениеклетчатки отмечается при 28-30С. Весной, сповышением температуры в воде и почве,численность бактерий начинает резкоувеличиваться. Наиболее интенсивноклетчатка разлагается при влажности 70% отполной влагоемкости почвы. При осеннемпонижении температуры воздуха, воды ипочвы, рост микроорганизмов падает. Принизких температурах (менее 8°С) и привысоких (более 55С) разложение целлюлозызамедляется.
Таким образом,основными абиотическими факторами,определяющими активностьмикроорганизмов-деструктороврастительного опада, являются влажность итемпература. Наиболее высокая скоростьдеструкции растительного опаданаблюдается в пырейно-гетеропаппусовомсообществе. Полученныеколичественные данные о распространениимикроорганизмов разных физиологическихгрупп и об их активности в растительныхопадах характеризует темпы минерализацииорганического вещества в залежныхфитоценозах Центральной Тувы.
Глава VI. Залежь какфактор экологизации земледелия:ретроспективный анализ и состояниепроблемы
На конференции вРио-де-Жанейро (1992), на Саммите Тысячелетияв ООН (2000), на Всемирном Саммите поУстойчивому Развитию (ВСУР) вИоханнесбурге (2002) и на Третьем Всемирномконгрессе по охране природы Всемирногосоюза по охране природы (МСОП) (Таиланд, 2004)мировое сообщество признало, чтодостижение Устойчивого Развития являетсяобщемировым приоритетом.
Деградацияисключительно сильно трансформированныхландшафтов степной зоны, ведущая кснижению рентабельности сельскогохозяйства, с одной стороны, и быстраяпотеря биоразнообразия степного биома, сдругой, требуют перехода кнеразрушительному использованию исохранению степных экосистем насельскохозяйственных землях.
Общая площадьземельного фонда Республики Тыва на 1января 2005 года г. составила 16860,4 тыс. га.Пашни занимают 111,6 тыс. га, что составляет 5,6% от всех сельскохозяйственных угодий или0,7 % от общей площади земельного фондареспублики. Тыва является одним из ведущихпроизводителей животноводческойпродукции Сибири, но удельный весреспублики в производстве зернасоставляет 0,9 %.
Данные о динамикезалежной растительности в условиях Тывы влитературе отсутствуют, хотя залежнаясистема земледелия на территориисовременной Тывы существовала со временивхождения в состав СССР (1944 г.). Средисоседних регионов Восточной СибириРеспублика Тыва характеризуется менееблагоприятными для земледелияпочвенно-климатическими условиями, в 1950годы - незначительной распаханностьюсельхозугодий, в которых почти 90 %приходится на долю пастбищ (Солдатов, 1967). В1930 году в валовой продукции кочевыхаратских хозяйств Тувы на долю земледелияприходилось всего 11%. Всю остальнуюпродукцию давало животноводство.
В период массовогоосвоения новых земель в использованииземельного фонда республики были допущеныошибки. В ряде случаев распашка земельпроводилось без тщательных почвенныхисследований. Также отсутствиеобоснованной системы целинного земледелияпривело к выпадению из оборота около 20 тыс.га пашни. По данным 1973 г. из общей площадипашни республики подвержено эрозии,засолено, засорено камнями около 25 %.
Современное кризисноесостояние сельского хозяйства Тывыявляется следствием деградациисельскохозяйственных угодий в периодконца 70-х начала 90-х годов XX века. В условияхрезко континентального климата Тывыразвитие богарного земледелия себя неоправдала. По данным Кондратьева А.Б. иШкоркина А.И. (1969) в Тувинской АССР пшеницаво многих районах возделывается наорошаемых землях. Урожай пшеницы приорошении, в 2-3 раза выше, чем набогаре.
По данным В.А.Носина (1963)чероземы Тывы в 1960 году имели: гумуса всреднем 6,2%, а пахотный горизонт составлял 46см., соответственно каштановые почвы: 4,8– 40 см. К началу1980-х годов содержание гумуса в черноземахснизилось до 2,1-2,5 %, а мощность пахотногогоризонта до 31 см. Подобная картинаотмечается и на каштановых почвах. Этопроизошло в силу того, что повсеместно безучета экологии почв внедрялась системаземледелия, в которой значительное местобыло уделено мобилизации потенциальногоплодородия за счет минерализацииорганического вещества пашни.
Земледелие натерритории современной Тывы зародилось в1950-х годах XX в. С переходом АПК в рыночныеотношения на низко плодородных почвахвозделывание зерновых культур сталоэкономически неэффективным, поэтому около50% пашни запущено в залежь. Исходя из этого,интересно проследить, как происходитвосстановление плодородия почвы.
Период бурьянистого перелога представлен группировками: а) сучастием Artemisia sieversiana, A.vulgaris, Sonchus oleraeceus,Barbarea arcuata и др.; б) принедостаточном увлажнении -мелкобурьянистые: Artemisia scoparia,Chenopodium album и другие. Видырастений имеют хорошо развитую корневуюсистему, проникающую иногда на глубину дометра и более, обладающие повышеннойпоглотительной способностью. Онипоглощают из почвы те минеральныевещества, которые не доступны культурнымрастениям и переводят их в доступные формыв процессе биологического круговоротавеществ в биогеоценозе агроэкосистемы. PHпахотного горизонта бурьянистого перелогаможет достигать 7,5 – 7,7; то есть повышаться по сравнению скультурной пашней на 0,4 – 0,5 единиц.
Основным доминантомкорневищной стадии является Elytigiarepens. Это растение встречаетсятакже в бурьянистой стадии, но оно угнетенокрупно- и мелкобурьянными сорняками. Впырейном перелоге pH снижается до 7,3 – 7,35.
Рыхлодерновиннаястадия характеризуетсяналичием злаков и разнотравья. Доминантомявляется Poa attenuata, P.pratensis. Из злаков встречаютсяKoeleria cristata, Phleum phleoides, из разнотравья Artemisiaannua, A. siversiana.
Стадияплотнокустовых растений,представлена отдельными пятнами Stipa krylovii. Пофитоценотическому и физиономическомусоставу она медленно, но приближается кфитоценозам целинных степей и pH почвыстановится близкой к нейтральной.
Агроландшафтно-экологическоерайонирование представляет собой единуюнаучно-обоснованную систему делениятерритории, учитывающую закономерностираспределения природных комплексов, атакже особенности их использования вкормопроизводстве, оптимизации земельныхресурсов, рациональном природопользованиив сельском хозяйстве. Единицырайонирования обоснованы системойпоказателей, включающей климат, структуруземельных и кормовых угодий, рельеф, почвы,растительность и их продуктивность, атакже мероприятия по управлению и созданиюагроэкосистем. Новые показатели, впервыеиспользуемые при районировании, делают егоболее наукоемким, системным и актуальным,имеющим большое практическоезначение.
Увеличениепроизводства продукции растениеводствапроисходит в основном за счет ростаурожайности. В решении этой задачи большоезначение имеет разработка и применениеагротехнических мероприятий с учетомособенностей природных зон. Совершенноочевидно, что в этом важном деле большуюроль играет правильное деление территориина природные зоны. Следует отметить, чтоопытом природного районированиятерритории республики в разное времязанимались многие ученые – естествоиспытателии специалисты сельского хозяйства.Природное районирование Тывы на основематериалов Тувинской комплекснойэкспедиции и почвенного районированиядано В.А. Носиным (1963). Солдатовым В.П. в 1967году в соответствии с группировкой районовпо сходным природным и экономическимусловиям на территории Тывы выделено 4 зоныи 3 подзоны: I. Центральная зона -земледельческо-животноводческая; подзоны– подтаежная,преимущественно земледельческая и степная–животноводческо-зерновая. II. Западная зона–животноводческо-земледельческая; подзонаживотноводческая Монгун-Тайга. III. Южнаязона –преимущественно овцеводческая. IV.Восточная зона -животноводческо-промысловая.
Тем не менее,общепринятая система деления наагрохозяйственные природные зоны врегионе все еще не сложилась. Принятое с 1963года разделение территории республики надесять почвенно-климатических зон с цельюразработки мероприятий по увеличениюпроизводства сельскохозяйственнойпродукции и получившее известноераспространение, нельзя признать научнообоснованным. Подходы эколого-адаптивногоприродопользования должна стать основойагро-экологического районированияземельных ресурсов Республики Тыва.
ВЫВОДЫ
1. Природные особенностиТувы выявили определяющую рольорографических и климатических факторовна процессы формирования степнойрастительности региона. Степи в Тывесоставляет основу ландшафтов какмежгорных котловин, так и обрамляющих иххребтов и нагорий. При этом выявленызакономерности, в условиях плакоров,подгорных равнин господствуют сообществанастоящих дерновиннозлаковых иопустыненных полукустарничковых степей.По периферии межгорных депрессий, вграницах лесного и лесостепного поясовраспространены фитоценозы луговых степей.В более континентальных и сухихвысокогорьях юго-западной Тувы характернысообщества криофитных степей. Во всехпоясах широко развиты серии петрофитныхгруппировок степной растительности.
2. Степная флора Тувынасчитывает 785 видов высших сосудистыхрастений (272 рода и 54 семейства). Анализ эколого-фитоценотических групппоказал, что главнейшеезначение в степях Тувы имеют горно-степныеи лесостепные виды призаметном снижении степных ипустынно-степных. Пограничное положение территорииТувы на стыке лесной (Южной Сибири) ипустынно-степной (Центральной Азии)областей объясняет значительное участиево флоре северо-азиатских мезоксерофитныхбореальных и южносибирско-монгольскихксерофитных горностепных элементов, чтопозволяет охарактеризовать ее степнуюфлору как горнуюбореально-гемиаридную.
3. Список растенийзалежных фитосистем вЦентрально-Тувинской котловине включает 110видов сосудистых растений, относящихся к 27семействам и 80 родам. Однако, видовоебогатство семейств Asteraceae,Poaceae, Fabaceae характеризуютаридные черты флоры. В то же время вразнообразии видов в семействах Rosaceae, Brassicaceae, Caryophyllaceae, Ranunculaceaeпроявляются черты гумидности. Богатствопредствителей Asteraceae (33%) выражается в обилии адвентивныхвидов. Высокий удельный вес злаковыхрастений Poaceae(27%), показатель восстановления степнойрастительности. Оригинальнаяклассификация видового составарастительности залежей Тувы сустановлением трех категорийфлористического комплекса залежей (ФКЗ), аименно –типично залежных, переходных и инвазийных(или случайно залежных) типов ФКЗ имеютиндикационное значение по особенностям ихдемутации.
4.Эколого-фитоценотическая классификациястепной растительности с использованиемдоминантно-детерминантного подходапозволила выделить 28 формаций, 13 группформаций, относящиеся к 4 классам формаций(или подтипам) степной растительности. Приэтом объективизация выделения синтаксоновдостигнута с использованием методаэкологической ординации сообществ степей.Своеобразие и оригинальность степям Тувыпридают сообщества опустыненных степей сэдификаторами – нанофитоном, полынью дернистой,ковылем галечным, а также заметное участиезападных степных элементов (ковыльперистый, осока низкая, скабиозабледно-желтая) в сочетании сдауро-монгольскими горностепными видами(цимбария даурская, володушкакозелецелистная).
5. При изучениипроцессов естественного восстановлениязалежей Центрально-Тувинской котловиныТывы выявлены залежные стадиивосстановления: бурьянистая(мелкобурьянистая), корневищная,рыхлокустовая и плотнокустовая. Уровеньбиологической продуктивности залежныхсообществ зависит от их возраста, видовогосостава растений, слагающих конкретныесообщества, а также почвенно-экологическихусловий их местообитания. Наибольшаяпродуктивность надземной фитомассыхарактерна для сообществ рыхлокустовойстадии - гетеропаппусово-змеевковомсообществе в 2005-2006 годы. Наименьшаяпродуктивность наблюдается впырейно-гетеропаппусовом сообществе,свойственной мелкобурьянисто-корневищнойстадии демутации.
6. Микробиологическаядеструкция растительного опада являетсяважнейшим процессом, определяющимбиологический круговорот элементов вприроде. При этом вклад различныхфизиологических групп бактерий в аэробнуюи анаэробную деструкцию органическоговещества растительных остатков имеютособенности на различных стадияхдемутации. Динамика численностимикроорганизмов-деструктороворганического вещества растительногоопада залежных экосистем Центральной Тувыв зависимости от влияния абиотическихфакторов (температуры и влажности) посезонам показали, что наибольшая скоростьразложения белка наблюдается вранне-осенний период, а целлюлозы – летом. При этомактивность целлюлозоразлагающих бактерийнаблюдается в начале и особенно, в концелета с повышением влажности почвы.
7. Растительный покровстепей в настоящее время испытываетсильное влияние антропогенного фактора.Для рационального использованиягорностепной растительности в качествекормовых угодий необходимо введениесезонной системы пастбищеоборота,соблюдение норм и правил традиционногоопыта в использовании пастбищ. Особенностипроцессов демутации на залежах, а такжерезультаты ретроспективного анализа вземлепользовании в регионе позволилиопределить приоритеты эколого-адаптивногоприродопользования как перспективной длясухостепных ландшафтов межгорных котловинТувы.
Списокосновных работ, опубликованных по темедиссертации
Монографии:
Ондар С.О., ДубровскийН.Г.Биологические системы в разных временныхмасштабах. Ч. I. Организация, интеграция,регуляция и эволюция биологических системв экологическом времени. Новосибирск:СибАГС, 2005. - 558 с.
Дубровский Н.Г., Назын-оол О.А. Зерновые культуры,возделываемые в Туве. – Кызыл: РИО ТывГУ,2005. – 56с.
Дубровский Н.Г.,Назын-оол О.А.,Черепанов М.Е. Применениеудобрений на дефлированных почвахРеспублики Тыва. Кызыл: ТывГУ КЦО «Аныяк»,2006. - 255 с.
Ондар С.О., ДубровскийН.Г.Биологические системы в разных временныхмасштабах. Ч. II, III. Организация, интеграция,регуляция и эволюция биологических системв экологическом времени. Новосибирск:СибАГС, 2006. - 787 с.
Определитель растенийРеспублики Тыва / Красноборов И.М.,Ломоносова М.Н., Лайдып А.М., Шауло Д.Н.,Дубровский Н.Г. и др. – Новосибирск: Наука, 2007. – 335 с.
Статьи врекомендованных ВАК изданиях:
Дубровский Н.Г. Удобрения надефлированных почвах и продуктивностьяровой пшеницы / Н.П.Аюшинов,А.А.Атыгаев, Н.Г.Дубровский, Н.Г.Солдатова,В.М.Соловьева, О.А.Назын-оол, Н.А.Ховалыг,Е.А.Порядина // Земледелие,№2, 2005. - С.11-12.
Дубровский Н.Г. Особенностизалежной сукцессии в Туве / Н.Г.Дубровский,Б.Б.Намзалов, А.В.Ооржак// Вестник Бурятскогоуниверситета: сер.2, вып.№7, 2005. - С.200-205.
Дубровский Н.Г. К классификациинастоящих дерновинно-злаковых степейнагорья Сангилен (Юго-Восточная Тыва) /К.В.Кыргыс, Н.Г.Дубровский,Б.Б.Намзалов // Вестник Бурятского университета:сер.2, вып.№8, 2006. - С. 122-127.
Дубровский Н.Г. К характеристикефлористического комплекса залежнойрастительности Тывы /А.В.Ооржак, Н.Г.Дубровский //Вестник Бурятского госуниверситета:вып.№3, 2007. –С. 169-172.
Дубровский Н.Г. Продуктивность идеструкция растительного опада в залежныхсообществах Тывы /А.В.Ооржак, Н.Г.Дубровский, В.Б.Дамбаев,Б.Б.Намзалов // ВестникБурятского госуниверситета: вып.№3, 2007.– С.184-188.
Дубровский Н.Г. Особенности территориальногораспределения и фитоценотическогоразнообразия растительности залежейТувы / Н.Г.Дубровский, А.В.Ооржак //Вестник ЧГПУ: № 3, 2009. – С. 263-275.
Дубровский Н.Г. Классификация и особенностидемутации залежной растительностиЦентральной Тувы / Н.Г.Дубровский,А.В.Ооржак, Б.Б.Намзалов // Вестник ЧГПУ:№ 3, 2009. –С. 276-292.
В другихизданиях:
Дубровский Н.Г. Рациональноеиспользование преобразование и охранабиологических ресурсов Тувы /Н.Г.Дубровский // Сборник статей. – Кызыл, 1990. – С. 7-10.
Дубровский Н.Г.Природопользование и экологическоевоспитание / Н.Г.Дубровский, Е.Э.Ондар //Материалы Международной конференции,посвященной 70-летию тувинскойписьменности (11-14 сентября). - Кызыл, 2000.– С.105-106.
Дубровский Н.Г. К развитию теориидемутационного процесса и особенностифлоры залежной растительности Тувы / Б.Б.Намзалов., Н.Г.Дубровский,А.В.Ооржак // МатериалыВсероссийской научно-практическойконференции «Опыт и традиции этническогоприродопользования» (17-19 сент. 2003). – Улан-Удэ: Изд-воБГСХА, 2003. –С. 48-52.
Дубровский Н.Г. Состав удобрения«биогумус» и его влияние на рост и развитиерастений в неблагоприятных условияхпроизрастания» /С.С.Потапова, Н.Г.Дубровский // Материалы IV съезда Докучаевскогообщества почвоведов «Почвы национальноедостояние России» (9-13 августа 2004).Новосибирск: Наука – Центр, 2004.– С. 296-297.
Дубровский Н.Г. К анализу флорызалежной растительности Тувы / А.В.Ооржак, Н.Г.Дубровский //. Материалы Всероссийской научнойконференции с международным участием«Проблемы сохранения разнообразиярастительного покрова внутренней Азии» (7-10сентября 2004 г.) – г. Улан-Удэ: Изд-во Бурятскогонаучного центра СО РАН, 2004, С. 128-130.
Андрейчик М.Ф., Дубровский Н.Г.Атлас облаков РеспубликиТыва (в летнее время года). – Кызыл, 2004. – 100 с.
Дубровский Н.Г. Условиязагрязнения атмосферы горных котловинРеспублики Тыва /М.Ф.Андрейчик, Н.Г.Дубровский // Материалы VII Международнойконференции «Природные условия, история икультура западной Монголии и сопредельныхрегионов» (20-22 сентября 2005 г., г. Кызыл,Россия). –Кызыл: Изд-во ТувИКОПР СО РАН, 2005. Т.2., С.217-219.
Дубровский Н.Г. Современноесостояние и вопросы охраны редких степныхсообществ нагорья Сангилен / К.В. Кыргыс,Н.Г.,Дубровский // Материалы VII Междунар.конф. «Природные условия, история икультура Западной Монголиии сопредельных регионов». - Кызыл, 2005. - Т.1. -С. 144-148.
Дубровский Н.Г. Особенности использования степныхпастбищ нагорья Сангилен (на примеретрадиций тувинского этноса) / К.В.Кыргыс, Н.Г.Дубровский,Б.Б.Намзалов // МатериалыПятой международной научно-практическойконференции “Проблемы ботаники ЮжнойСибири и Монголии” (Барнаул, 21-23 ноября 2006г.). - Барнаул:Изд-во «АзБука», 2006. – С.141-143.
Дубровский Н.Г. Основные этапывзаимоотношений в системе «ЭТНОСЫ(популяции людей) – ПРИРОДА (экосистема)» / Б.Б.Намзалов, Л.К.Аракчаа,Н.Г.Дубровский // Материалынаучно-методического семинара «Этническаяэкология и традиционноеприродопользование на рубеже веков»(Улан-Удэ, 30 ноября 2006 г.). - Улан-Удэ: изд-воБурятского госуниверситета, 2006. - С.5-10.
Дубровский Н.Г. Залежные экосистемы Тувы:особенности флоры и демутационногопроцесса / Н.Г.Дубровский, Б.Б.Намзалов //Флора и растительность Алтая. Т. 11, 2006.– С. 89-93.
Дубровский Н.Г. Краткая история исследованиярастительности степей и залежей Тувы /Н.Г.Дубровский, Б.Б.Намзалов, А.Д.Самбуу //Биота в экосистемах гор Южной Сибири:состояние и проблемы. Байкальскийэкологический вестник. – Вып. 4. – Улан-Удэ: Изд-воБурятского госуниверситета, 2007. – С. 4 -15.
Дубровский Н.Г. Методологияизучения разнообразия фитосистем (на примерерастительности Республики Тыва) /Б.Б.Намзалов, Н.Г.Дубровский // Биота вэкосистемах гор Южной Сибири: состояние ипроблемы. Байкальский экологическийвестник. –Вып. 4. –Улан-Удэ: Изд-во Бурятскогогосуниверситета, 2007б. – С. 15 - 28.
Дубровский Н.Г. Анализ структуры флоры степей Тувы/ Н.Г. Дубровский, Б.Б.Намзалов, К.В.Кыргыс,А.М.Самдан // Биота в экосистемах гор ЮжнойСибири: состояние и проблемы. Байкальскийэкологический вестник. – Вып. 4. – Улан-Удэ: Изд-воБурятского госуниверситета, 2007. – С. 28 - 39.
Дубровский Н.Г. Степнаярастительность Тувы, классификация ихарактеристика разнообразия / Б.Б.Намзалов,Н.Г.Дубровский, К.В.Кыргыс // Биота вэкосистемах гор Южной Сибири: состояние ипроблемы. Байкальский экологическийвестник. –Вып. 4. –Улан-Удэ: Изд-во Бурятскогогосуниверситета, 2007. – С. 42 - 54.
Намзалов Б.Б., Дубровский Н.Г.Очерки о высокогорных степях Тувы иЮго-Восточного Алтая – Улан-Удэ: Изд-воБурятского госуниверситета, 2007а. – 37 с.
Дубровский Н.Г. О некоторых теоретическихаспектах изучения залежной растительности(на примере Республики Тыва) / Н.Г.Дубровский, Б.Б.Намзалов,А.В.Ооржак // МатериалыВсероссийской конференции с международнымучастием (Иркутск, 21-25 мая 2007 г.)“Синантропизация растений и животных”.Иркутск: изд-во Института географии СО РАНим. В.Б. Сочавы, 2007. - С.35-37.
Дубровский Н.Г. К анализу степной растительностигорной лесостепи хр. Тану-Ола / Б.Б.Намзалов,Н.Г.Дубровский, А.М.Монгуш // Материалы второй Международнойнаучно-практической конференции.«Биоразнообразие и сохранение генофондафлоры, фауны и народонаселенияЦентрально-Азиатского региона». - Кызыл, 2007.- С. 36-38.
Дубровский Н.Г. Род Artemisia L. на залежнойфлоре Центрально-Тувинской котловины / А.В.Ооржак, Н.Г.Дубровский, Б.Б. Намзалов// Материалы второйМеждународной научно-практическойконференции. «Биоразнообразие исохранение генофонда флоры, фауны инародонаселения Центрально-Азиатскогорегиона». - Кызыл, 2007. - С. 53-55.
Дубровский Н.Г. О некоторыхаспектах эколого-исторического анализаземлепользования в Республике Тыва /К.В.Кыргыс, Н.Г.Дубровский, Б.Б.Намзалов// Мир кочевойцивилизации: история и современность.Матер. межд. научн. конференции (10-12 октября2007, Чита –Агинское). –Чита: Экспресс изд-во, 2007. – С. 327-330.