

Особенности системы семенного размножения у представителей семейства asteraceae саратовской области
На правах рукописи
Кочанова Ирина Сергеевна
ОСОБЕННОСТИ СИСТЕМЫ СЕМЕННОГО РАЗМНОЖЕНИЯ
У ПРЕДСТАВИТЕЛЕЙ СЕМЕЙСТВА ASTERACEAE
САРАТОВСКОЙ ОБЛАСТИ
03.00.05 – ботаника
Автореферат диссертации на соискание ученой степени
кандидата биологических наук
Саратов – 2008
Работа выполнена в государственном образовательном
учреждении высшего профессионального образования
«Саратовский государственный университет им. Н.Г. Чернышевского»
на базе УНЦ «Ботанический сад» СГУ
| Научный руководитель: | доктор биологических наук, профессор Кашин Александр Степанович |
| Официальные оппоненты: | доктор биологических наук, старший научный сотрудник Эльконин Лев Александрович кандидат биологических наук, доцент Алаторцева Татьяна Алексеевна |
| Ведущая организация: | Ботанический институт им. В. Л. Комарова РАН |
Защита состоится « 3 » июля 2008 г. в 11.00 часов на заседании диссертационного совета Д 212.243.13 при государственном образовательном учреждении высшего профессионального образования «Саратовский государственный университет им. Н.Г. Чернышевского» по адресу: 410012, г. Саратов, ул. Астраханская, д. 83; E-mail: [email protected].
С диссертацией можно ознакомиться в Зональной научной библиотеке ГОУ ВПО «Саратовский государственный университет им. Н.Г. Чернышевского».
Автореферат разослан « 1 » июня 2008 г.

Ученый секретарь
диссертационного совета С. А. Невский
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность темы. Важной проблемой репродуктивной биологии является изучение особенностей процессов воспроизводства в различных таксонах и экологически дифференцированных группах растений на всех уровнях их организации (Терёхин, 1988, 2000; Salisbury, 1942; Wilson, 1983). Эти процессы во многом определяют генетическую структуру популяций, а также адаптационные возможности и эволюционный потенциал растений (Grant, 1981).
Механизмы, или элементы систем размножения, изменение которых оказывает существенное влияние на генетическую структуру популяций вида и характер протекающих в них генетических процессов называют параметрами систем размножения (Куприянов, 1989). Особенно важными для системы семенного размножения оказываются такие основные параметры, как способ опыления (аллогамия, автогамия) и способ образования семян (амфимиксис, апомиксис). Однако степень изученности этих параметров в популяциях покрытосеменных растений остаётся недостаточной. Цитоэмбриологически исследовано менее 20% видов цветковых (Сравнительная…, 1987; Кашин, 2006). К тому же многие из этих видов изучены явно недостаточно для того, чтобы с уверенностью судить о способах опыления или образования семян.
От полноты знаний о параметрах системы семенного размножения зависит сама возможность и эффективность использования этих знаний для решения различных фундаментальных и прикладных проблем. В этом отношении особенно нагляден пример с изучением природы гаметофитного апомиксиса. В последние годы значительно возрос интерес к проблеме гаметофитного апомиксиса в связи с тем, что генетический анализ показал относительно простую генетическую природу апомейоза (Savidan, 1982; Nogler, 1984). Были начаты интенсивные исследования по локализации гена(ов) апомиксиса с целью их передачи методами генной инженерии культурным растениям (Bicknell, Borst, 1994; Kindiger et al., 1996; Grossniklaus et al., 2001) и широкого использования апомиксиса в селекции и растениеводстве. Это важно, в частности, для решения задач сохранения гетерозисного эффекта в ряду поколений, стабилизации отдаленных гибридов и уникальных генных комбинаций, что обещает настоящую революцию в растениеводстве (Koltunov et al., 1995; Savidan, 1995, 2001; Vielle Calzada et al., 1996). Однако существенных успехов в этой области достигнуто не было. Прежде всего, это связано с: 1) почти полным отсутствием близких сородичей культурных растений, у которых был бы известен гаметофитный апомиксис и которые могли бы послужить донорами генов апомиксиса; 2) фрагментарностью знаний о закономерностях распространения этого явления в природе и своеобразия его проявления на уровне системы семенного размножения.
Цель и задачи исследования. Цель настоящей работы заключалась в выявлении частоты апо- и амфимиксиса, авто- и аллогамии в популяциях видов семейства Asteraceae во флоре Саратовской области с оценкой толерантности апомиктов к условиям обитания на антропогенно трансформированных территориях и широты распространения апомиктичных видов в семействе. В задачи работы входило:
- проанализировать семенную продуктивность растений при разных режимах цветения: свободном опылении, в условиях изоляции некастрированных цветков и беспыльцевом режиме;
- цитоэмбриологически исследовать популяции и выявить у растений цитоэмбриологические признаки гаметофитного апомиксиса;
- выявить адаптивный потенциал апомиктичных видов при освоении ими антропогенно трансформированных территорий.
Научная новизна работы. Впервые проведено исследование видов семейства Asteraceae в региональной флоре с целью выявления апомиктичных форм. Гаметофитный апомиксис впервые обнаружен у 13 видов (Hieracium largum, Pilosella echioides, Tragopogon dubius, Scorzonera ensifolia, Chondrilla latifolia, C. сanescens, Aster bessarabicus, Bidens tripartita, Inula britanica, Jurinea cyanoides, J. arachnoidea, Galatella linosyris, Artemisia vulgaris) и в пяти родах (Tragopogon, Scorzonera, Jurinea, Inula и Galatella) семейства Asteraceae.
Показано, что для выявления способности растений вида формировать семена апомиктичным путем необходимо неоднократное изучение данного параметра в популяциях, так как у многих видов апомиксис проявляется не во все годы и не во всех популяциях.
Установлено, что толерантность апомиктичных видов к условиям обитания на антропогенно трансформированных территориях не уступает, а, скорее, оказывается выше толерантности к ним амфимиктичных видов.
Научно-практическая значимость работы. Полученные данные могут использоваться для уточнения списка и характера распространения апомиктичных видов цветковых, а также для подбора доноров генов апомиксиса с целью передачи их культурным сородичам. Разработанная методика выявления частоты амфи- апомиксиса и авто-аллогамии в популяциях может быть использована при определении этих параметров у других представителей цветковых. Материалы диссертации могут быть использованы в университетах России при разработке спецкурсов по репродуктивной биологии, эмбриологии растений, экологии видов цветковых, биоразнообразию. Исследованные видообразцы включены в уникальную коллекцию апомиктичных видов Ботанического сада СГУ.
Апробация работы. Результаты работы были доложены на: Международной научной конференции «Вопросы общей ботаники: традиции и перспективы», посвященной 200-летию Казанской ботанической школы (Казань, 2006); Международной конференции молодых ботаников (Санкт-Петербург, 2006); III съезде ВОГиС «Генетика в XXI веке: современное состояние и перспективы развития» (Москва, 2004); XII молодежной научной конференции «Актуальные проблемы биологии и экологии» (Сыктывкар, 2005); II Международной научной конференции студентов, аспирантов и молодых ученых «Биоразнообразие. Экология. Эволюция. Адаптация», посвященной 140-летию Одесского национального университета им. И.И. Мечникова (Одесса, 2005); V и VI Международных научно-практических конференциях «Проблемы ботаники Южной Сибири и Монголии» (Барнаул, 2006, 2007); Всероссийской научной конференции «Ботанические исследования в Поволжье и на Урале», посвященной 50-летию Ботанического сада СГУ им. Н.Г. Чернышевского (Саратов, 2006); VIII Съезде украинского общества генетиков и селекционеров им. Н.И. Вавилова (Алушта, Украина, 2007), II Международной школе молодых ученых «Эмбриология, генетика и биотехнология» (Уфа, 2007); ежегодных конференциях студентов и аспирантов биологического факультета СГУ им. Н.Г. Чернышевского (Саратов, 2006, 2007).
Публикации. По теме диссертации опубликовано 24 работы, две из которых – в изданиях перечня ВАК РФ.
Декларация личного участия автора. Автор лично провел экспедиционные исследования, включающие сбор и фиксацию экспериментального материала для цитоэмбриологических исследований, кастрацию и изоляцию соцветий для исследования семенной продуктивности. Автором самостоятельно приготовлены и проанализированы микроскопические препараты, сделаны микрофотографии. Анализ и обработка полученных данных осуществлены на 90% самостоятельно, по плану, согласованному с научным руководителем. Доля личного участия в написании совместных публикаций составляла 50-70%.
Объем и структура диссертации. Диссертация состоит из введения, шести глав, заключения, выводов, списка литературы и приложения. Общий объем работы составляет 140 страниц, работа содержит 26 таблиц и 12 рисунков. Список литературы включает 228 источников, в том числе 128 на иностранных языках.
Основные положения, выносимые на защиту:
- гаметофитный апомиксис имеет широкое распространение у видов семейства Asteraceae во флоре Саратовской области;
- апомикты не уступают амфимиктам в толерантности к условиям обитания на антропогенно трансформированных территориях;
- методика определения частоты апомиксиса по семенной продуктивности при беспыльцевом режиме цветения в популяциях автономных апомиктов позволяет выявлять гаметофитный апомиксис с высокой надёжностью.
СОДЕРЖАНИЕ РАБОТЫ
Глава 1. ОСОБЕННОСТИ СИСТЕМЫ СЕМЕННОГО РАЗМНОЖЕНИЯ У ЦВЕТКОВЫХ (обзор литературы)
В главе приводятся сведения о степени изученности цветковых в отношении способа опыления и способа образования семян, широте распространения апомиксиса по данным различных авторов (Поддубная-Арнольди, 1976; Хохлов и др., 1978; Fryxell, 1957; Hanna, Bachaw, 1987, Asker, Jerling, 1992; Carman, 1995, 1997) и об адаптационном потенциале апомиктов. Доказывается, что у растений видов Asteraceae возможны только гаметофитные, но не спорофитные формы апомиксиса.
Глава 2. МАТЕРИАЛ И МЕТОДЫ
Семенную продуктивность при различных режимах цветения определяли по материалам, собранным в 2003-2007 гг. в 157 естественных популяциях 96 видов 44 родов из двух подсемейств (Asteroidea и Cichorioidea) семейства Astе-raceae, произрастающих в 13 районах Саратовской области. Выбор популяций и видов осуществлялся случайным образом. При этом часть одних и тех же популяций исследовалась в течение нескольких лет (от 2 до 8), начиная с 2000 года.
Частоту амфи- и апомиксиса, алло- и автогамии в популяциях выявляли на основе сравнительного анализа данных о семенной продуктивности растений при свободном цветении, цветении в условиях изоляции некастрированных цветков и беспыльцевом режиме цветения. Для анализа завязываемости семян при цветении в условиях изоляции некастрированных цветков и беспыльцевого режима соцветия до начала цветения краевых цветков помещали под пергаментные изоляторы, под которыми они находились до полного созревания семян. Для создания беспыльцевого режима цветения цветки на стадии зрелого бутона предварительно механически кастрировали путем срезания верхней части соцветия вместе с пыльниками на уровне перехода венчика цветка в завязь.
В каждой популяции в среднем исследовали 30 растений, отобранных случайным образом. Процент завязываемости семян в соцветии вычисляли как отношение числа завязавшихся семян к общему числу цветков в соцветии. Частоту апомиксиса в популяции определяли как процентное отношение завязываемости семян при цветении в условиях беспыльцевого режима к завязываемости семян при свободном режиме цветения. Частоту автогамии в популяциях определяли как процентное отношение разности между завязываемостью семян при цветении в условиях изоляции некастрированных цветков и завязываемостью семян при беспыльцевом режиме цветения к семенной продуктивности при свободном цветении.
Виды сложноцветных, у которых обнаружили гаметофитный апомиксис по семенной продуктивности, подвергали дополнительному эмбриологическому контролю. Для этого соцветия за 1-3 суток до цветения (раскрытия бутона) краевых цветков фиксировали в фиксаторе Кларка. Препараты зародышевых мешков готовили по ускоренной методике П.Г. Куприянова (1982, 1989) с использованием мацерирующего агента (цитазы) и микропрепаровальных игл или методики просветления семязачатков (Herr, 1971). Материал окрашивали ацетокармином с предварительной обработкой железоаммонийными квасцами.
Анализ препаратов проводили с помощью микроскопа Axiostar plus (Zeiss) в светлом поле в проходящем свете или при фазовом контрасте под увеличением 10 х 40. Микрофотографирование осуществляли с использованием видеоадаптера 426106-9130-000 при помощи цифровой камеры Canon Power Shot A 620.
Результаты анализа статистически обработаны. Вариационные ряды обсчитывали с помощью программного продукта Microsoft Office Excel 2007. Сравнение вариационных рядов осуществляли по критерию Стьюдента (tst) при уровне значимости Р = 0.95 (Зайцев, 1984). В случае близости величин среднеарифметической к 0 или 100% для определения доверительного интервала использовали метод (Плохинский, 1970). Для сравнения качественных признаков использовали метод вычисления средней ошибки при альтернативном распределении (Зайцев, 1984).
Глава 3. СЕМЕННАЯ ПРОДУКТИВНОСТЬ В ПОПУЛЯЦИЯХ ВИДОВ
СЕМЕЙСТВА ASTERACEAE
Частота гаметофитного апомиксиса в исследованных популяциях видов семейства Asteraceae. Исследовано около 40% видов из почти 75% родов семейства Asteraceae Саратовской области. Как следует из табл. 1, из общего числа исследованных популяций семена в условиях беспыльцевого режима цветения, то есть апомиктично, завязывались у растений популяций 13 видов 9 родов подсемейства Cichorioidea. В 2 родах (Tragopogon, Scorzonera) и у 6 видов (Hieracium largum, Pilosella echioides, Tragopogon dubius, Scorzonera ensifolia, Chondrilla latifolia, C. сanescens ) апомиксис отмечен впервые.
Таблица 1
Частота завязываемости семян путём апомиксиса в популяциях видов Asterасеae
| Популяции видов | Семенная продуктивность при, % | Частота апомиксиса, % | |
| свободном цветении | беспыльцевом режиме | ||
| Aster bessarabicus | 64.3±6.9 | 70.5±5.7 | 100.0 |
| Bidens frondosa | 72.5±3.8 | 20.0±8.8 | 27.6 |
| B. tripartitа | 100.0±0.0 | 96.1±2.5 | 96.1 |
| Chondrilla canescens | 25.9±6.2 | 24.0±8.9 | 92.7 |
| C. graminea | 60.5±12.6 | 30.7±15.3 | 50.7 |
| C. juncea | 53.1±4.1 | 32.4±6.2 | 61.0 |
| C. latifolia | 97.2±1.9 | 76.2±4.3 | 78.4 |
| Galatella linosyris | 60.2±8.0 | 11.5±5.4 | 19.1 |
| Hieracium virosum | 87.9±6.8 | 58.5±10.1 | 66.5 |
| H. largum | 64.7±6.6 | 53.2±8.3 | 82.2 |
| Inula britanica | 72.9±9.5 | 12.5±6.4 | 17.2 |
| Jurinea arachnoidea | 85.3±5.2 | 65.5±9.1 | 76.8 |
| J. cyanoides | 93.5±4.0 | 63.6±9.0 | 68.0 |
| Lactuca serriola | 90.0±10.0 | 14.3±4.3 | 15.9 |
| Pilosella echioides | 53.0±8.2 | 58.4±8.2 | 100.0 |
| P. officinarum | 52.3±5.5 | 73.1 ±11.3 | 100.0 |
| P. praealta | 73.4±5.0 | 55.5±3.0 | 75.6 |
| Scorzonera ensifolia | 63.3±6.1 | 17.5+4.3 | 27.6 |
| Taraxacum officinale | 100.0±0.0 | 78.8±6.9 | 78.8 |
| Tragopogon dubius | 42.1±4.9 | 32.2±5.0 | 76.4 |
В подсемействе Asteroidea апомиксис обнаружен в популяциях 10 видов 7 родов. Впервые апомиктичный способ репродукции зарегистрирован в 3 родах (Jurinea, Inula, Galatella) и у 7 видов (Aster bessarabicus, Bidens frondosa, Inula britanica, Jurinea cyanoides, J. arachnoidea, Galatella linosyris и Artemisia vulgaris). В пределах данного подсемейства апомиксис ранее отмечался крайне редко.
Характер изменчивости частоты гаметофитного апомиксиса на внутрипопуляционном и межпопуляционном уровнях. Показано, что в разных популяциях одного и того же вида и даже в одной и той же популяции в различные годы наблюдений частота апомиксиса может существенно варьировать, вплоть до практически облигатного проявления апо- или амфимиксиса.
Так одна из популяций (22а) апоспорового апомикта P. officinarum, обитающая на влажном лугу в Б.-Карабулакском районе, неизменно вела себя как факультативно апомиктичная с достаточно высокой выраженностью апомиксиса в любой год наблюдений (25.9 - 73.1%). Вторая популяция (33а), обитающая на опушке остепнённого соснового бора в том же районе и удалённая от первой всего лишь на расстояние около 3 км, в различные годы вела себя то как амфимиктичная, то как факультативно апомиктичная. В 2003 г. семена в условиях беспыльцевого режима у растений данной популяции не завязались, в 2004 г. семенная продуктивность при беспыльцевом режиме цветения составила около 10%, а в 2005 г. – более 25%, в 2007 г. часть растений этой популяции оказались апомиктичными, а часть – амфимиктичными (рис. 1.А, Б).
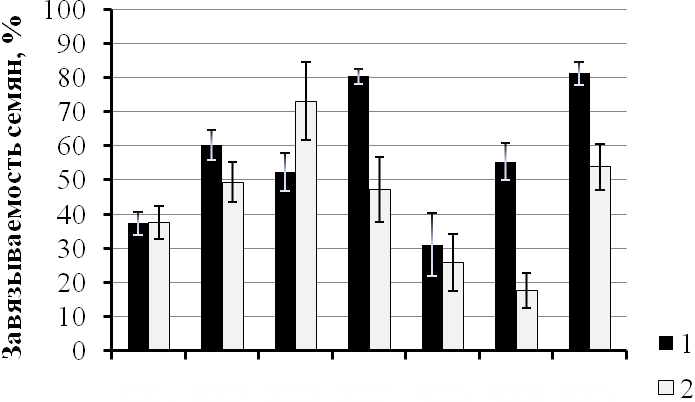
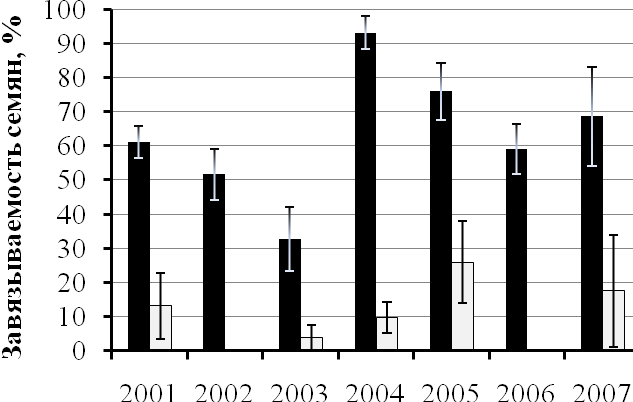
Рис 1. Динамика семенной продуктивности при беспыльцевом режиме цветения растений в популяциях P. officinarum 22а (биотоп влажного луга) (А) и 33а (биотоп остепнённого соснового бора) (Б) по годам: 1 – семенная продуктивность при свободном цветении, 2 – семенная продуктивность при беспыльцевом режиме цветения
Аналогично вели себя популяции C. juncea, P. echioides, J. сyanoides, I. britanica и ряда других видов (табл. 2).
Из полученных результатов следует, что динамика изменчивости способа размножения в популяциях апоспорических видов является существенной как на внутрипопуляционном, так и на межпопуляционном уровнях, причём факторы, её определяющие, носят локальный, биотопный характер проявления.
Таблица 2
Динамика семенной продуктивности популяций некоторых видов при беспыльцевом режиме цветения
| № популяции и вид | Завязываемость семян при беспыльцевом режиме цветения, % | |||||||
| 2000 | 2001 | 2002 | 2003 | 2004 | 2005 | 2006 | 2007 | |
| 48а Taraxacum officinale | 64.2±7.3 | 97.3±9.9 | 53.8±17.0 | 78.8±6.9 | 58.5±9.3 | 42.9±7.6 | ||
| 92 T. officinale | 63.0±9.3 | 93.9±1.3 | 77.4±9.0 | 61.5±8.9 | 54.0±0.2 | 93.4+3.8 | 75.5±9.2 | |
| 67 Chondrila juncea | 52.3±12.8 | 27.7±14.0 | 0 | 3.6±1.4 | 0 | 0 | 0 | |
| 94 C. juncea | 42.5±10.1 | 27.7±7.0 | 17.6±6.9 | 14.2±5.7 | 0 | 0 | 7.5±1.5 | 25.0±8.9 |
| 22ф Pilosella echioides | 0 | 0 | 0 | 0 | 0 | 4.7±0.5 | 7.3±0.7 | |
| 33ф P. echioides. | 0 | 0 | 0 | 0 | 0 | 15.8±1.1 | 6.2±0.6 | |
| 164 P. echioides | 58.4±8.2 | 15.5±1.0 | ||||||
| 203 Jurinea сyanoides | 54.8±11.0 | 0 | 0 | 0 | ||||
| 332 Inula britanica | 12.5±0.6 | 0 | 0 | |||||
Примечание: по незаполненным ячейкам данных нет
В пространственно значительно разделённых популяциях диплоспорового апомикта Taraxacum officinale наблюдалась синхронность изменений выраженности гаметофитного апомиксиса по годам. Сходно вели себя и популяции диплоспорового апомикта C. juncea. Это говорит о том, что факторы, сказывающиеся на динамике изменчивости частоты гаметофитного апомиксиса диплоспорового типа, вероятнее всего, относятся к климатическим факторам относительно глобального характера, т.е. сходно проявляющимся назначительных территориях в тот или иной год и одинаково воздействующим на механизм выбора пути семенной репродукции у растений пространственно разделённых популяций видов.
Характер изменчивости частоты гаметофитного апомиксиса в популяциях видов Asteraceae показывает, что для установления способа семенного размножения у растений тех или иных популяций и видов зачастую недостаточно разового исследования в течение одного полевого сезона. Для выявления способности растений вида формировать семена апомиктичным путем необходимо неоднократное изучение данного параметра в популяциях, так как у многих видов апомиксис проявляется не во все годы и не во всех популяциях.
Частота алло- и автогамии в исследованных популяциях видов семейства Asteraceae. У растений популяций Pulicaria vulgaris, Lactuca serriola, Erigeron acris, Tripleurospermum perforatum, Onopordum acanthium, Carduus acanthoides, Tragopogon ruthenicus, Serratula erucifolia, Arctium lappa, A. tomentosum, Lapsana communis, Inula britanica, Hieracium umbellatum, S. palustris, а в ряде случаев и у Chondrilla juncea, C. graminea, Pilosella officinarum и P. echioides, отмечена высокая завязываемость семян при цветении в условиях изоляции некастрированных цветков, но семена не завязывались, либо с более низкой частотой завя-зывались в условиях беспыльцевого режима цветения (табл. 3). При этом популяции Pulicaria vulgaris, O. acanthium и A. tomentosum являются облигатными автогамами, т.к. семенная продуктивность в них при этом режиме цветения была близка к 100% или достоверно не отличалась от семенной продуктивности при свободном режиме цветения. Остальные из перечисленных популяций факультативно аллогамны, - семенная продуктивность при режиме цветения в условиях изоляции некастрированных цветков у них была на уровне 10 – 20%, либо существенно варьировала по годам или на межпопуляционном уровне.
В число указанных облигатно или факультативно автогамных видов попали и виды, у которых известна или показана способность к факультативному апомиксису (C. juncea, Pilosella officinarum, P. praealta), а также сородичи апомиктичных видов (E. acris). Это ставит под сомнение сложив-шееся представления о том, что апомиктичные растения имеют амфимиктичных
Таблица 3
Исследованные виды семейства Asteraceae, у которых обнаружена автогамия
| № популяции и вид | Семенная продуктивность при, % | Реальная частота автогамии, % | ||
| свободном режиме цветения | изоляции некастрированных цветков | беспыльцевом режиме | ||
| 168 Pulicaria vulgaris | 93.2±5.1 | 93.8±6.3 | 0 | 100.0 |
| 148 Erigeron acris | 63.1±15.9 | 36.9±9.8 | 2.8±0.1 | 54.0 |
| 151 Tripleurospermum perforatum | 48.9±6.2 | 11.3±5.7 | 7.4±0.4 | 8.0 |
| 178 Onopordum acanthium | 100.0±0.0 | 86.9±5.3 | 3.9±0.2 | 83.0 |
| 179 Carduus acanthoides | 72.5±4.3 | 10.0±4.5 | 0 | 13.8 |
| 180 Laсtuca serriola | 90.0±10.0 | 21.8±10.6 | 14.3±4.3 | 8.3 |
| 145 Tragopogon dubius | 63.4±7.3 | 10.5±3.3 | 18.3±5.4 | 0 |
| 223 Hieracium virosum | 87.9±6.8 | 59.3±10.9 | 58.4±10.1 | 1.0 |
| 94 Chondrilla juncea | 84.0±4.6 | 81.3±6.9 | 0 | 96.8 |
| 22а Pilosella officinalrum | 81.2±3.5 | 64.2±5.3 | 47.2±9.6 | 20.9 |
| 22г P. praealta | 73.4±5.0 | 68.0±1.9 | 55.5±3.0 | 17.3 |
| 203 Jurinea cyanoides | 93.6±2.8 | 76.7±9.0 | 54.8±11.0 | 23.4 |
| 332 J. arachnoidea | 85.3±5.2 | 77.5±11.7 | 65.5±9.1 | 14.1 |
| 390 Aster bessarabicum | 64.3±6.9 | 58.4±7.5 | 70.5±5.7 | 0 |
| 212 Arctium tomentosum | 47.9±7.4 | 57.9±10.9 | 0 | 100.0 |
| 275 A. lappa | 54.6±6.7 | 27.1±7.2 | 0 | 49.6 |
| 395 Bidens tripartita | 100.0±0.0 | 96.5±2.5 | 96.1±2.5 | 0 |
| 396 B. frondosa | 72.5±3.8 | 42.6±8.4 | 20.0±8.8 | 31.2 |
| 276 Sonchus arvensis | 32.5±9.2 | 56.6±6.4 | 0 | 100.0 |
| 252 Lapsana communis | 84.8±6.2 | 47.2±8.9 | 0 | 55.7 |
сородичей, неизменно относящихся к перекрёстно опыляющимся видам, у которых перекрёстное оплодотворение обеспечивается самонесовместимостью, двудомностью или какой-либо другой системой семенного воспроизводства по типу ауткроссинга (Gustafsson, 1946, 1947; Grant, 1984).
Глава 4. ЦИТОЭМБРИОЛОГИЧЕСКОЕ ИЗУЧЕНИЕ ЧАСТОТЫ
АПОМИКСИСА В ИССЛЕДОВАННЫХ ПОПУЛЯЦИЯХ
Проведенный цитоэмбриологический анализ показал, что для растений популяций видов семейства Asteraceae, у которых впервые обнаружен апомиксис по семенной продуктивности (H. largum, P. echioides, Tragopogon dubius, Scorzonera ensifolia, C latifolia, C. сanescens, Aster bessarabicus, B. frondosa, I. britanica, J. cyanoides, G. linosyris, Artemisia vulgaris), характерна высокая частота формирования клеток, морфологически подобных апоспорическим инициалям, в присутствии тетрады мегаспор или эуспорических зародышевых мешков разной стадии развития, а также преждевременная эмбриония и (или) автономный эндоспермогенез (табл. 4). У видов, семенная продуктивность которых при
Таблица 4
Результаты эмбриологического исследования видов семейства Asteraceae, у которых впервые обнаружен
апомиксис по семенной продуктивности
| Вид | Норма, % | Дегенерация эуспорических ЗМ, % | С признаками апомиксиса, % | ||||||
| всего | из них | ||||||||
| эндосперм без оплодотворения | преждевременная эмбриония | эуспорический ЗМ и клетки, подобные апоспорическим инициалям | клетки, подобные апоспорическим инициалям, при преждевременной эмбрионии в эуспорическом ЗМ | клетки, подобные апоспорическим инициалям, при дегенерации эуспорического ЗМ | преждевременная эмбриония + эндосперм без оплодотворения | ||||
| Artemisia vulgaris | 69.4 | 0 | 30.6 | 0 | 0 | 30.6 | 0 | 0 | 0 |
| Aster bessarabicus | 51.3 | 0 | 48.7 | 0 | 0 | 48.7 | 0 | 0 | 0 |
| Inula britanica | 94.6 | 3.8 | 1.9 | 1.9 | 0 | 0 | 0 | 0 | 0 |
| Bidens tripartita | 57.9 | 0 | 42.1 | 0 | 0 | 40.3 | 1.8 | 0 | 0 |
| B. frondosa | 42.9 | 12.9 | 44.3 | 1.3 | 2.9 | 31.4 | 1.4 | 7.1 | 0 |
| Chondrilla canescens | 46.2 | 2.6 | 51.2 | 8.6 | 42.6 | 0 | 0 | 0 | 0 |
| C. latifolia | 77.4 | 5.3 | 17.3 | 0 | 17.3 | 0 | 0 | 0 | 0 |
| Galatella linosyris | 71.6 | 5.1 | 23.3 | 0 | 0 | 20.1 | 0 | 3.2 | 0 |
| Jurinea cyanoides | 85.0 | 1.0 | 13.2 | 2.1 | 8.1 | 2.0 | 0 | 0 | 1.0 |
| Scorzonera ensifolia | 69.1 | 15.8 | 15.1 | 3.2 | 9.7 | 0 | 0 | 0 | 2.2 |
| Pilosella echioides | 99.1 | 0 | 0.9 | 0.3 | 0.6 | 0 | 0 | 0 | 0 |
| Lactuca serriola | 99.7 | 0.3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Tragopogon dubius | 97.7 | 0.9 | 2.3 | 0 | 2.3 | 0 | 0 | 0 | 0 |
беспыльцевом режиме цветения равнялась нулю, подобного рода образования в семязачатках растений ни разу не отмечались, несмотря на большой объём цитоэмбриологических исследований. Это говорит в пользу того, что методика определения частоты апомиксиса по семенной продуктивности при беспыльцевом режиме цветения позволяет с высокой надёжностью выявлять данный параметр в популяциях автономных апомиктов.
Результаты проведённых исследований указывают на то, что части видов, у которых гаметофитный апомиксис обнаружен впервые, свойственна апоспория (Aster bessarabicus, B. tripartitа, B. frondosa, G. linosyris, Artemisia vulgaris, J. cyanoides). Растениям видов C. canescens, C. latifolia, S. ensifolia, I. britanica, скорее всего, свойственна диплоспория. Имеют место существенные различия в темпах развития проэмбрио и эндосперма без оплодотворения. Об этом свидетельствует присутствие в исследован-ном материале, в одних случаях, семязачатков с опережающим развитием про-эмбрио, в других, - семязачатков с опережающим развитием эндосперма, в третьих, - семязачатков с одновременным развитием проэмбрио и эндосперма.
Глава 5. ШИРОТА РАСПРОСТРАНЕНИЯ ГАМЕТОФИТНОГО
АПОМИКСИСА У ЦВЕТКОВЫХ
(НА ПРИМЕРЕ СЕМЕЙСТВА ASTERACEAE)
В ходе достаточно локального исследования популяций лишь половины видов одного семейства Asteraceae в одной региональной флоре гаметофитный апомиксис впервые отмечен для 5 родов и 13 видов. Это расширяет список J. Carman (1995, 1997) в отношении апомиктичных родов на четверть, а список апомиктичных видов – более чем на 10% (рис. 2). При этом речь идёт о семействе, к которому принадлежит 1/4 часть выявленных на сегодняшний день в


Рис.2 Графическое изображение соотношения количества родов (1) и видов (3) семейства Asteraceae, у которых апомиксис впервые установлен в данном исследовании, и количества родов (2) и видов (4), у которых апомиксис был известен ранее (Carman, 1995, 1997)
пределах всего отдела цветковых растений видов и 1/5 часть выявленных на сегодняшний день в пределах всего отдела цветковых растений родов.
Таким образом, если исследование лишь половины видов и 3/4 родов в одном только регионе вынуждает так корректировать список апомиктичных форм семейства, обнаруженных более чем за 100 лет предыдущих исследований, то о какой-либо полноте представлений о широте распространения апомиктичных форм, по крайней мере, в семействе Asteraceae, в настоящее время речи быть не может. В данном семействе насчитывается около 1250 родов и порядка 20000 видов (Тахтаджян, 1987) (рис. 3). Поэтому только для данного семейства потенциально можно ожидать наличие гаметофитного апомиксиса, как способа семенного размножения, ещё у 1000 - 1500 видов порядка 150 родов. Очевидно, что это значительно превышает общее число родов и видов цветковых, у которых на сегодняшний день выявлен апомиксис: по числу родов - более чем в пять раз, по числу видов – более чем в 8 – 12 раз.

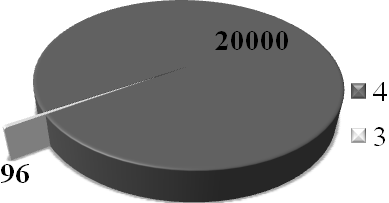
Рис.3 Графическое изображение доли исследованных в данной работе родов (1) и видов (3) семейства от общего числа родов(2) и видов (4), известных в семействе Asteraceae
Учитывая вышеизложенное, следует ожидать, что и в других семействах, - по крайней мере, в тех основных, в которых известен апомиксис, - доля апомиктичных видов и родов окажется выше, чем это известно в настоящее время.
Глава 6. ТОЛЕРАНТНОСТЬ РАСТЕНИЙ АМФИ- И АПОМИКТИЧНЫХ ВИДОВ СЕМЕЙСТВА ASTERACEAE К АНТРОПОГЕННО
ТРАНСФОРМИРОВАННЫМ ТЕРРИТОРИЯМ
При исследовании флоры окрестностей г. Саратова, обнаружено 132 вида из 53 родов семейства Asteraceae. Из этого числа на антропогенных биотопах в границах непрерывной городской застройки обнаружено 88 видов (или 66,6%). Из сложноцветных окрестностей города в отношении семенной продуктивности при трёх режимах цветения исследовано 89 видов 44 родов семейства, т.е. 2/3 от общего числа видов и около 85% родов, обитающих на данной территории.
Интересно, что доля массово и нередко встречающихся в черте городской застройки видов семейства выше среди апомиктов (почти 60% исследованных апомиктичных видов) и существенно ниже среди амфимиктов (лишь 1/3 исследованных амфимиктичных видов) (табл. 5). Доля встречающихся на урбанизированных территориях видов при этом среди амфимиктичных около 50%, в то время как среди апомиктичных – более 85%. Эти данные говорят в пользу того, что апомиктичные виды имеют высокую конкурентноспособность при адаптации к условиям урбанизированной среды. На других основных типах антропогенных биотопах (техногенные местообитания, искусственные лесные насаждения, агрофитоценозы) также доля апомиктичных видов преобладает над амфимиктичными на 10-20% (табл. 6).
В целом на исследованных антропогенно трансформированных биотопах встречается около 75% амфимиктичных и около 90% апомиктичных видов. Это указывает на то, что адаптивный потенциал апомиктов за счёт комплекса
Таблица 5
Встречаемость исследованных в отношении способа семенного размножения видов
Семейства Asteraceae на антропогенных биотопах в границах городской застройки
| Способы семенной репродукции | Число видов, абс. | Из них встречаются на антропогенных биотопах, % | ||||
| всего | массово | нередко | редко | не встречается | ||
| Только амфимиктичный | 73 | 54.8±5.0 | 23.3 | 11.0 | 20.5 | 45.2 |
| Факультативно апомиктичный | 21 | 85.7±6.5 | 19.1 | 38.1 | 28.5 | 14.3 |
Таблица 6
Толерантность к различным типам неурбанизированных антропогенных биотопов
исследованных видов семейства Asteraceae с различными способами размножения
| Способы семенной репродукции | Число видов, абс. | Из них встречаются на антропогенных биотопах, % | |||
| всего | в том числе | ||||
| техногенные местообитания | искусственные лесные насаждения | агроценозы | |||
| Амфимиктичный | 73 | 75.3±5.0 | 61.6±5.7 | 60.3±5.7 | 41.1±5.7 |
| Факультативно апомиктичный | 21 | 90.5±6.4 | 71.4±8.3 | 76.2±9.3 | 61.9±10.6 |
свойств, присущих этой системе семенного размножения, при освоении антропогенно трансформированных территорий отнюдь не уступает адаптивному потенциалу облигатных амфимиктов, а, скорее, превосходит, его.
ВЫВОДЫ
1. Растениям большинства (68.1%) исследованных популяций видов семейства Asteraceae Саратовской области присуще амфимиктичное аллогамное образование семян.
2. Способность к гаметофитному апомиксису по результатам изучения семенной продуктивности при беспыльцевом режиме цветения и цитоэмбриологических признаков выявлена в популяциях Taraxacum officinale, Pilosella officinarum, P. praealta, P. echioides, Hieracium virosum, H. largum, Jurinea cyanoides, J. arachnoidea, Chondrilla juncea, C. canescens, C. latifolia, C. graminea, Tragopogon dubius, Scorzonera ensifolia, Inula britanica, Astra bessarabicum, Bidens tripartita, B. frondosa, Galatella linosyris. При этом гаметофитный апомиксис отмечен впервые для 13 видов и 5 родов.
3. Растения популяций Pulicaria vulgaris, Onopordum acanthium и Arctium tomentosum являются облигатными автогамами. Растения популяций Latuca serriola, Erigeron acris, Matricaria perforata, Carduus acanthoides, Tragopogon ruthenicus, Serratula erucifolium, Arctium lappa, Lapsana communis, Serratula erucifolia, Inula britanica, Hieracium umbellatum, а в ряде случаев и Chondrilla juncea – факультативно аллогамны.
4. В популяциях видов семейства Asteraceae для установления способа семенного размножения у растений зачастую недостаточно разового исследования в течение одного полевого сезона одной популяции вида, т.к. имеет место существенная изменчивость частоты гаметофитного апомиксиса от почти облигатного до полного отсутствия.
5. Апомикты проявляют высокий адаптивный потенциал при освоении антропогенно трансформированных территорий.
СПИСОК ОСНОВНЫХ РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ
ДИССЕРТАЦИИ
* - публикации в печатных изданиях перечня ВАК РФ
- Добрыничева Н.В., Кочанова И.С., Кашин А.С. Основные параметры системы семенного размножения в популяциях ряда видов Asteraceae из Саратовской области // Бюллетень ботанического сада СГУ. Вып. 3. Саратов, 2004. С. 130 – 139.
- Добрыничева Н.В., Кочанова И.С., Кашин А.С. Устойчивость некоторых параметров системы семенного размножения в популяциях двух видов рода Chondrilla // Вопросы биологии, экологии, химии и методики обучения: Сб. науч. трудов. Вып. 7. Саратов, 2004. С. 146 – 150.
- Добрыничева Н.В., Кочанова И.С., Кашин А.С. Цитоэмбриологическое изучение частоты апомиксиса в популяциях Chondrilla juncea L. и C. graminea Bieb.// Бюллетень ботанического сада СГУ. Вып. 4. Саратов, 2005. С. 213 – 220.
- Добрыничева Н.В., Кочанова И.С., Кашин А.С. Основные параметры системы семенного размножения в популяциях видов Asteraceae. // Известия Саратовского университета. Вып.2. Серия Химия. Биология. Экология. Т. 5. Саратов, 2005. С. 30 – 32.
- * Кашин А.С., Добрыничева Н.В., Кочанова И.С., Демочко Ю.А. Особенности семенного размножения в популяциях Chondrilla juncea и Chondrilla graminae (Asteraceae) // Ботан. журн. 2006. Т. 91, № 5. С. 729 – 744.
- Кашин А.С., Березуцкий М.А., Кочанова И.С., Добрыничева Н.В. Особенности системы семенного размножения в популяциях некоторых видов Asteraceae в связи с их толерантностью к антропогенным местообитаниям // Поволжский экологический журнал. № 2/3. Саратов, 2006. С.139 – 146.
- Добрыничева Н.В., Кочанова И.С., Кашин А.С. Сравнительное изучение некоторых параметров системы семенного размножения в популяциях рода Chondrilla L. // Бюллетень ботанического сада СГУ. Вып. 5. Саратов, 2006. С. 307 – 312.
- Кочанова И.С., Добрыничева Н.В., Кашин А.С. Семенная продуктивность в половых и апомиктичных популяциях семейства Asteraceae // Бюллетень ботанического сада СГУ. Выпуск 5. Саратов: Научная книга, 2006. С. 312 – 316.
- Кашин А.С., Березуцкий М.А., Кочанова И.С., Полянская М.В. Характеристика параметров системы семенного размножения, влияющих на генетическую структуру популяций видов Asteraceae // Досягнення i проблеми генетики, селекцiї та бiотехнологiї: Зб. наук. пр. Киев, 2007. С. 86 – 90.
- * Кашин А.С., Березуцкий М.А., Кочанова И.С., Добрыничева Н.В., Полянская М.В. Основные параметры системы семенного размножения в популяциях некоторых видов Asteraceae в связи с действием антропогенных факторов // Ботан. журн. 2007. Т. 92, № 9. С. 1408 – 1427.