

Состав, структура и динамика зоопланктонного сообщества тауйской губы охотского моря
На правах рукописи
ВАКАТОВ Алексей Владимирович
состав, структура и динамика
зоопланктонного сообщества Тауйской
губы Охотского моря
03.00.16 - Экология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Казань – 2008
Работа выполнена на кафедре зоологии беспозвоночных ФГОУ ВПО «Казанский государственный университет им. В. И. Ульянова-Ленина» и в лаборатории морских промысловых рыб ФГУП «Магаданский научно-исследовательский институт рыбного хозяйства и океанографии»
Научный руководитель: кандидат биологических наук, доцент
Сабиров Рушан Мирзович
Официальные оппоненты: доктор биологических наук, профессор
Мингазова Нафиса Мансуровна
Казанский государственный университет
кандидат биологических наук, старший
научный сотрудник
Примаков Игорь Михайлович
Зоологический институт РАН,
г. Санкт-Петербург
Ведущая организация: ФГОУ ВПО «Дальневосточный государственный
университет», г. Владивосток
Защита состоится 4 декабря 2008 г. в __________ часов на заседании диссертационного совета ДМ212.081.19 при ФГОУ ВПО «Казанский государственный университет им. В. И. Ульянова-Ленина» по адресу: г. Казань, ул. Кремлевская, 18, биолого-почвенный факультет, аудитория 211. Факс (843)238-71-21, (843)231-52-40, электронная почта [email protected]
С диссертацией можно ознакомиться в Научной библиотеке им. Н.И. Лобачевского Казанского государственного университета по адресу: г. Казань, ул. Кремлевская, 35.
Автореферат разослан «______» ноября 2008 г.
Ученый секретарь диссертационного совета,
кандидат биологических наук, доцент Р.М. Зелеев
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность темы. Тауйская губа Охотского моря имеет важное экономическое значение как район традиционного промысла лососевых рыб, воспроизводящихся в бассейнах рек Тауй и Яна, тихоокеанской сельди, камбал, колючего краба. На побережье губы находятся крупные промышленные центры Магаданской области, проходит граница Магаданского заповедника. На прилегающем к Тауйской губе участке Охотского моря сосредоточены значительные запасы минтая и сельди (Шунтов и др., 1990а, 1993а, 2003; Дулепова, 2002).
Планктон является основой кормовой базы массовых промысловых пелагических рыб минтая и сельди в Охотском море. Знание его количественных характеристик позволяет судить об условиях питания в районах нагула и воспроизводства рыб (Виноградов, Шушкина, 1987; Шунтов и др., 1993а). Пространственное распределение многих видов планктона, особенно калянид, служит хорошим индикатором течений и дислокации водных масс (Бродский, 1956, 1957; Беклемишев, 1957; Раймонт, 1983).
Помимо этого, в настоящее время очевидна недооценка индикаторной роли зоопланктона для Охотского моря. При сходном наборе видов зоопланктона в дальневосточных морях и северо-западной части Тихого океана (Бродский, 1957; Кун, 1975; Волков, 1996) большое индикаторное значение приобретают структурные характеристики сообществ, основанные на видовом составе, как наиболее чувствительные к изменениям окружающей среды, связанным с антропогенным воздействием (Одум, 1986). Сведения о составе планктона, его пространственном распределении и динамике представляют собой необходимую научно-теоретическую основу, без которой невозможно решение вопросов, связанных с изучением экологии пелагических планктоноядных рыб и с функционированием морских сообществ и экосистем.
По большинству районов Охотского моря информация о планктоне внутренней части неритической зоны отсутствует или скудная (Шунтов, 2001). Проведенные ранее исследования северной части Охотского моря оставляли без внимания прибрежную часть, где сосредоточены значительные запасы биоресурсов, представляющие важный промысловый интерес. Так сведения о планктоценозе Тауйской губы являются отрывочными и недостаточными для оценки функционирования водного сообщества. В последнее десятилетие прошлого века исследования планктоценоза не проводились. Следует отметить также, что за последние десятилетия климат и гидрологический режим Охотского моря претерпели заметные изменения (Груза, Ранькова, 1996; Матвеев, Мороз, 1997).
Цель работы: изучение зоопланктонного сообщества Тауйской губы и прилегающего участка Охотского моря, его видового состава, экологии, пространственно-временной изменчивости в сезонном и межгодовом аспектах.
Для достижения цели решались следующие задачи:
1. Изучить видовой состав фитопланктона (до родов) и его горизонтальное распределение в Тауйской губе в весенне-летний период.
2. Изучить качественные и количественные показатели зоопланктона Тауйской губы и прилегающего участка Охотского моря.
3. Выявить закономерности горизонтального распределения зоопланктона.
4. Изучить сезонную и межгодовую динамику зоопланктона Тауйской губы и прилегающего участка Охотского моря.
5. Исследовать значение зоопланктона в питании минтая и сельди.
Положения, выносимые на защиту.
1. Новые данные о составе, горизонтальном распределении, сезонной и межгодовой динамике зоопланктона Тауйской губы Охотского моря.
2. Планктонное сообщество Тауйской губы является гетерогенным по своему составу и происхождению и включает автохтонный комплекс прибрежного и надшельфового зоопланктона и аллохтонные виды из открытых вод северной части Охотского моря, привносимые восточной ветвью Ямского течения.
3. Прибрежное планктонное сообщество Тауйской губы участвует в формировании продуктивной надшельфовой зоны северной части Охотского моря (Притауйский промрайон) через систему западной выносящей ветви Ямского течения.
Научная новизна. Впервые проведено подробное исследование зоопланктонного сообщества Тауйской губы Охотского моря, дана полная экологическая характеристика планктона, исследованы количественные и качественные изменения в зоопланктонном сообществе в межгодовом аспекте, изучено влияние гидрологических условий на состав и структуру сообщества. Впервые показано, что планктонное сообщество Тауйской губы является гетерогенным по своему составу и происхождению и формируется из автохтонного прибрежного и надшельфового зоопланктона и привносимых Ямским течением видов из открытых вод северной части Охотского моря.
Научно-практическая значимость. Полученные данные дополняют общие представления о структуре планктонного сообщества неритической зоны северной части Охотского моря, включающего прибрежный и надшельфовый планктоценозы, и могут быть использованы в уточнении промыслового районирования Охотского моря.
Прибрежный планктоценоз Тауйской губы принимает непосредственное участие в формировании зон высокой биологической продуктивности в северной части Охотского моря, в связи, с чем промысел массовых промысловых рыб (сельдь, минтай) необходимо планировать с учетом продуктивности прибрежного планктонного сообщества Тауйской губы. Полученные данные о структуре и динамике планктонного сообщества Тауйской губы могут быть использованы при организации мониторинга за состоянием экосистемы северной части Охотского моря в условиях активного развития поисково-разведывательных работ и перспективы масштабной добычи углеводородного сырья на данном участке шельфа.
Апробация работы и публикации. Результаты исследований докладывались на Итоговых научных конференциях Казанского университета (2005-2008 гг.), на Отчетных конференциях МагаданНИРО (2005, 2006 гг.), представлялись на VI научной конференции «Сохранение биоразнообразия Камчатки и прилегающих морей» (Петропавловск-Камчатский, 2005 г.), XI и X Международных школах-конференциях молодых ученых «Биология – наука XXI века» (Пущино, 2005, 2006 гг.), Всероссийской конференции «Эколого-биологические проблемы вод и биоресурсов: пути решения» (Ульяновск, 2007 г.), Международных чтениях памяти В.Л. Вагина «Морская зоология и паразитология: поиски и открытия» (Казань, 2007), VI Всероссийской школе по морской биологии «Биоразнообразие сообществ морских и пресноводных экосистем России» (Мурманск, 2007), Международном симпозиуме ECSA-42 «Estuarine Ecosystems: structure, function and management» (Светлогорск, 2007).
Декларация личного участия автора. Автор принял личное участие в сборе, обработке и анализе материала по Тауйской губе и прилегающего участка северной части Охотского моря, собранного в рейсах БГК-627 (июнь 2005 г., 116 проб планктона; июль 2005 г., 50 проб планктона), НИС «Профессор Кагановский» (май 2006 г., 50 проб планктона, 74 пробы на питание рыб; май 2008 г., 50 проб планктона, 65 проб на питание рыб), на РЗ «Арманский» (май-июнь 2005 г., 20 проб на питание рыб). Автор обработал и проанализировал переданные материалы из рейсов СРТМк «Мастер» (октябрь-ноябрь 2003 г., 28 проб планктона), БГК-627 (июнь 2004 г., 116 проб планктона; июнь 2006 г., 30 проб планктона), на РЗ «Арманский» (май-июнь 2002 г., 25 проб на питание рыб; май-июнь 2006 г., 14 проб на питание рыб). Мы также проанализировали переданные нам карточки обработки 77 планктонных проб из рейса НИС «Зодиак» (август-сентябрь 2000 г.). В целом доля личного участия автора в сборе, обработке и анализе использованных данных составляет около от общего объема материала, задействованного при написании диссертации.
Публикации. По теме диссертации опубликованы 9 работ, в т.ч. 3 статьи в изданиях, рекомендованных ВАК. Одна статья принята к печати.
Структура и объем диссертации. Диссертация состоит из введения, 4 глав, заключения, выводов и списка литературы, включающего 213 источников, в т.ч. 7 на иностранном языке. Работа изложена на 191 странице, включает 51 рисунок, 24 таблицы, 17 приложений с результатами статистической обработки данных.
Благодарности. Автор выражает глубокую благодарность своему научному руководителю к.б.н., доценту Р. М. Сабирову за неоценимую помощь в работе над диссертацией. Я особо признателен зав. сектором фоновых исследований Магаданского научно-исследовательского института рыбного хозяйства и океанографии С. А. Шершенковой, с.н.с. В. Д. Жарниковой за передачу части материалов, помощь в обработке гидробиологических проб, ценные советы и замечания. Данное исследование было бы невозможным без поддержки и неоценимой помощи в сборе и обработке материалов зав. лабораторией морских промысловых рыб МагаданНИРО к.б.н. А. А. Смирнова, сотрудников данной лаборатории С. Ю. Шершенкова и Е. В. Кащенко, за что я выражаю им самую искреннюю благодарность. Я также очень благодарен зав. лабораторией гидробиологии Тихоокеанского института рыбного хозяйства и океанографии (ТИНРО-центр, г. Владивосток), д.б.н. В. И. Чучукало, а также к.б.н., в.н.с. лаборатории гидробиологии К. М. Горбатенко, к.б.н., н.с. А. Е. Лаженцеву за передачу в наше пользование гидробиологических материалов по северной части Охотского моря, ценные советы и замечания, высказанные при обсуждении работы. Автор считает своим приятным долгом выразить признательность всему коллективу кафедры зоологии беспозвоночных Казанского университета за постоянную поддержку и всяческое содействие в выполнении данной работы.
- ИСТОРИЯ ИЗУЧЕНИЯ ПЛАНКТОНА
ОХОТСКОГО МОРЯ
Широкомасштабные исследования Охотского моря были начаты в ряде комплексных экспедиций с начала 30-х гг. под руководством К. М. Дерюгина, П. В.Ушакова, Г. У. Линдберга и др. (Дерюгин, 1935; Кусморская, 1940; Киселев, 1947; Кун, 1981). В результате экспедиций были получены данные по видовому составу зоопланктона, его пространственному распределению, сезонной изменчивости. С середины 1980-х гг. начался новый этап изучения планктонных сообществ, связанный с экосистемными исследованиями биоресурсов Охотского моря. При экосистемном подходе в изучении пелагиали одновременно учитывали гидрологическую обстановку, гидрохимические характеристики, уловы рыб, количество планктона и питание планктофагов. (Волков, 1986, 1988, 1995, 1997; Волков, Ефимкин, 1990, 2002; Шунтов и др., 1993а, 1998; Чучукало и др., 1998). Достаточно подробное описание макропланктона северной части Охотского моря дается в работе Н. Н. Афанасьева (1985), состав, структура и динамика зоопланктона Охотского моря описаны К.М. Горбатенко (1997), зоопланктон Охотского моря его роль в питании нектона обсуждается в работе А.Ф. Волкова (1986).
В целом, в истории изучения планктона Охотского моря прослеживаются три основных направления. Первый – период изучения микроводорослей и беспозвоночных, составляющих планктон, описание и установление их положения в таксономической системе. Важнейшие работы этого периода – И. А. Киселева «Фитопланктон дальневосточных морей как показатель особенностей их гидрологического режима» (1947), К. А. Бродского «Фауна веслоногих рачков (Calanoida) и зоогеографическое районирование северной части Тихого океана и сопредельных вод» (1957). Второе направление связано с выявлением количественных характеристик планктонных организмов. Впервые работы по учету количества планктона стали проводиться в комплексе с определением количества биогенных элементов и гидрологических характеристик. Наиболее значимые работы этого периода – К. В. Беклемишева «Связь распределения планктона с распределением водных масс в зоне фронтов северо-западной части Тихого океана» (1958), Котляр Л.К., Чернявского В.И. «Распределение характерных представителей Calanoida в зависимости от режима года и некоторые черты их экологии» (1970). Третье направление – экспериментальное. Это изучение жизненных циклов, роста, поведения, физиологии, размножения, которое направлено на решение проблемы круговорота веществ в море и его продуктивности.
- МАТЕРИАЛ И МЕТОДЫ
Материалом для данной работы послужили планктонные сборы весенне-летних и летне-осенних комплексных съемок Тауйской губы и прилегающего участка северной части Охотского моря в период с 2000 по 2008 гг. Материал был собран в экспедициях (2000, 2003-2006, 2008 гг.) Общее количество проанализированных проб составило: планктона и ихтиопланктона - 517, проб на питание – 198 (3647 желудков рыб) (рис. 1а,б).
Планктон облавливался сетью «Джеди» (площадь входного отверстия 0,1 м2, ячея фильтрующего конуса 0,168 мм). Скорость подъема сети составляла 1м/с. На каждой из станций измеряли температуру, соленость, глубину зондом SBE-25 и батитермографом. Камеральную обработку проб производили в лабораторных условиях по фракциям в соответствии с методикой, принятой в ТИНРО-центре (Волков, 1996). Крупная фракция просчитывалась полностью. Численность мелкой фракции копепод подсчитывалась в камере Богорова по видам и стадиям развития. Затем их биомасса рассчитывалась по стандартным сырым весам согласно методике Е. А. Лубны–Герцык (1959). Общая биомасса пробы планктона определялась путем взвешивания на электронных весах FX-320 с точностью до 0,001 г. Обработку проб по питанию рыб проводили в соответствии с «Методическим пособием…» (1974) и «Руководством…» (Чучукало, Волков, 1986). Статистическую обработку данных проводили с использованием программ Excel, Statistica 5, анализ и представление результатов проводили с использованием программ Surfer 8 и Power Point.
Для оценки значимости видов нами был использован индекс плотности населения рассчитанный как корень квадратный из произведения биомассы
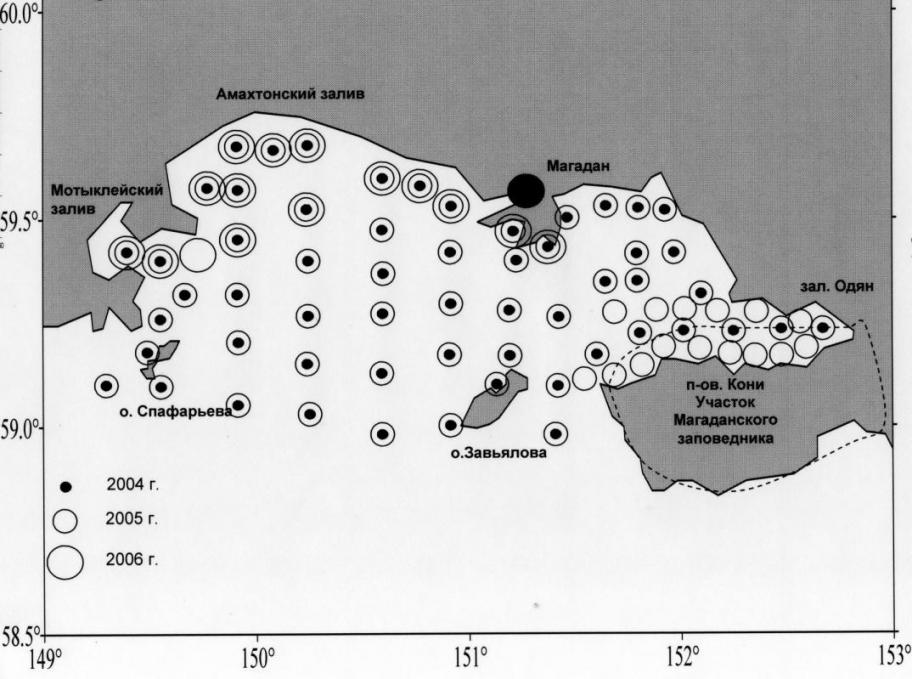
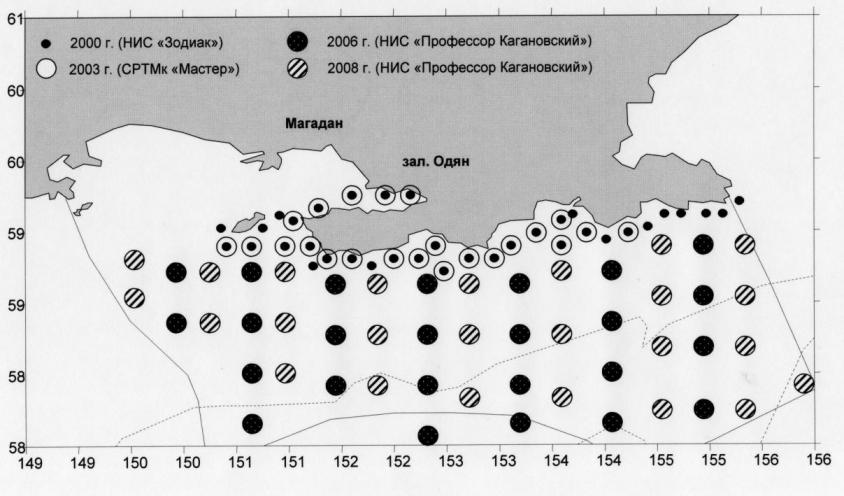
Рис.1. Карта-схема района работ в Тауйской губе (а) и
в прилегающем участке Охотского моря (б).
вида и плотности населения выраженный в процентах (Одум, 1986; Шитиков, 2003). Для сравнения степени видового сходства зоопланктона прибрежного, надшельфового и глубоководного сообществ использовали коэффициент общности видового состава Т. Съёренсена K=2с/(a+b) и коэффициент К. Чекановского:
 ,
,
где Xi и Yi – количественные значения вида i в пробах X и Y; S – общее число видов, и коэффициент видового сходства Константинова: K= i=1min (ai1,ai2), где ai1 и ai2 – соответственно биомасса вида i в долях от общей численности в 1-м и во 2-м сравниваемых сообществах; N – общее число видов в сравниваемых сообществах.
Для оценки видового биоразнообразия нами был использован индекс Шеннона-Уивера:
 ,
,
где, Pi – доля вида от общей биомассы зоопланктона (Шитиков, 2003).
3. результаты иССледований
3.1. Гидрологические условия в Тауйской губе
в период исследований
Период выполнения съемок, конец июня - начало июля, на североохотоморском шельфе относится к переходному сезону от поздней весны к лету. В первую очередь прогревается мелководье, холодные зоны отмечаются в динамически активных за счет интенсивного перемешивания районах, а также в районах подтока охотоморских вод. Минимальные значения отмечались в мористой части губы вокруг о. Завьялова и далее в центральной части до п-ова Старицкого (4,3-5,0°С), что было связано с затоком вод Ямского течения. Максимумы температуры в поверхностном слое были отмечены в западной части в заливах Амахтонский (11,9°С), Мотыклейский (13,9°С) и в восточной части в заливе Одян (10,2-11,0°С) (рис. 2). В придонном слое температура составляла от 2 до 5°С. Область минимальных значений (-1,1°С; 1,4°С) располагалась между островами Завьялова и Спафарьева и была ориентирована к п-ову Старицкого. Вторая область минимальных значений была расположена в восточной части Тауйской губы со стороны п-ова Кони (<0,5°С). Заток вод Ямского течения через Ольский пролив характеризовался температурой от 0,5 до 1,0°С. Минимальные значения солености отмечались в западной части Тауйской губы, где распресненный шлейф захватывал Амахтонский и Мотыклейский заливы (22,50-24,50‰) и распространялся вдоль берега через пролив Лихачева в открытое море. Минимальные значения (22,07‰) располагались в
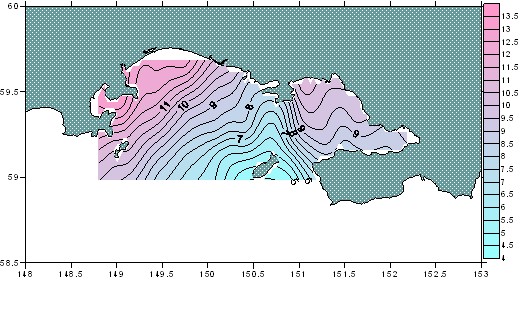
Рис. 2. Распределение температуры воды
поверхностного слоя в Тауйской губе в июле 2005 г.
приустьевых районах крупных рек, впадающих в Тауйскую губу (реки Ола, Армань, Яна и Тауй). Максимальные значения солености в поверхностном слое отмечались в Ольском проливе и вокруг о. Завьялова, где осуществлялся основной подток морских субарктических вод, в частности, Ямского течения (31,80-32,16‰) (рис. 3).

Рис. 3. Распределение солености в поверхностном
cлое в Тауйской губе в июле 2005 г.
Распределение солености у дна характеризовалось, в целом, увеличением абсолютных значений от берега (31,30-31,56‰) в глубоководную часть (32,86-32,97‰). Глубина распространения распресненного шлейфа в западной половине губы не превышала 7-9 метров.
3.2. Состав планктона и закономерности
его горизонтального распределения
Фитопланктон. В течение всего периода наблюдений на обследованной акватории Тауйской губы в фитопланктоне доминируют диатомовые водоросли: Coscinodicus, Thalassiozira, Rhizosolenia и Chaetoceros. Представители других систематических групп в составе фитопланктона не играли заметной роли. Фитопланктон был представлен, в основном, холодноводными формами неритического комплекса.
Образование циклонического круговорота в центральной части губы способствовало развитию максимумов биомассы фитопланктона: в центральной части Тауйской губы до 11471 мг/м и на выходе из губы между островами Завьялова, Спафарьева до 3140 мг/м. Средняя биомасса фитопланктона в Тауйской губе в весенне-летний период составила 1087 мг/м. С высоким содержанием фитопланктона выделялись прибрежные районы лиманов крупных рек, впадающих в Тауйскую губу. Высокая биомасса фитопланктона (более 10000 мг/м3) отмечена в заливе Одян. Значительных концентраций (свыше 5000 мг/м3) фитопланктон достигал в районе заливов Мотыклейский и Амахтонский. Биомасса микроводорослей от 500 до 1000 мг/м3 отмечалась между о. Завъялова и п-вом Старицкого, где осуществлялся приток более холодных субарктических вод. В центральной части Тауйской губы отмечались низкие биомассы фитопланктона (до 100 мг/м) которые были приурочены к водам с высокой соленостью (31-32‰).
Зоопланктон. Зоопланктон Тауйской губы представлен аркто-бореальными и бореальными холодноводными видами. В распределении доминирующих видов на акватории проявляется их приверженность водам определенной гидрологической структуры, где они образуют четко выраженные поля высокой плотности биомассы. Наиболее широкое распространение на акватории губы имеют веслоногие ракообразные (Copepoda) и эвфаузииды (Euphausiaceae), которые формируют от 70 до 90 % от общей биомассы зоопланктона Тауйской губы. Среди 30 видов, постоянно встречающихся в составе зоопланктонного сообщества Тауйской губы в весенне-летний период, в качестве структурообразующих нами были выделены 10 видов. Из них к доминантам относятся: Thysanoessa raschii, Metridia okhotensis, Pseudocalanus minutus, личинки Decapoda (рис. 4).

Рис. 4. Видовая структура зоопланктонного сообщества
Тауйской губы в весенне-летний период
К субдоминантам нами отнесены Neocalanus plumchrus, Calanus glacialis, Acartia longiremis, личинки Cirripedia, Microsetella rosea и Oithona similis остальные виды являются второстепенными.
В период исследований в весенне-летний период зоопланктон активно развивался на фоне вегетации диатомовых водорослей. Биомасса зоопланктона на акватории губы колебалась от 15 до 5691 мг/м, при среднем значении 953,6 мг/м. Максимальная биомасса зоопланктона (более 1000 мг/м3) отмечалась южнее и восточнее п-ова Старицкого и была приурочена к фронтальной зоне (район схождения распресненных вод Тауйской губы и вод Ямского течения) (рис. 5).
Максимальные концентрации (более 500 мг/м) мелкого зоопланктона были сосредоточены в Мотыклейском заливе, в прибрежье устья р. Тауй и Ола и заливе Одян. На этих участках зафиксирована максимальная температура поверхности воды (8,5-10,5С). На остальной акватории биомасса мелкой фракции колебалась от 50 до 150 мг/м3.
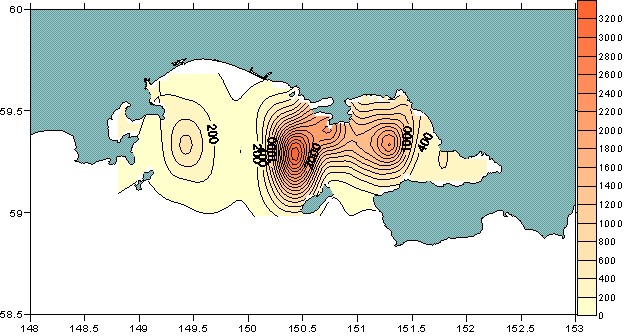
Рис. 5. Горизонтальное распределение общей биомассы
зоопланктона в Тауйской губе в весенне-летний период.
Основу мелкой фракции по биомассе составляли эвфаузииды (оva, nauplii, cаlyptopis), а по численности – копеподы. Основу средней фракции составляли крупные половозрелые особи P. minutus, A. longiremis и 3-4 копеподитных стадий N. plumchrus и M. okhotensis. Максимальная биомасса крупной фракции (более 2000 мг/м) отмечена в юго-восточной половине губы и была приурочена к периферии фронтальной зоны со стороны вод Тауйской губы.
Copepoda. Среди копепод доминировали P. minutus и М. okhotensis. Незначительную часть мелкой фракции составляли N. plumchrus, C.glacialis (1-2 копеподитные стадии), а также O.similis и A.longiremis.
M. оkhotensis. На долю этого вида приходилось до 50 % от биомассы копепод и 10 % от общей биомассы зоопланктона. Максимальные концентрации M. оkhotensis (от 100 до 323 мг/м3 и от 500 до 2460 экз./м3) в виде отдельных пятен встречалась в заливе Одян, Ольском лимане, в центральной части у п-ва Старицкого, в Амахтонском заливе на 3 и 4 копеподитных стадиях. В глубоководных участках на выходе из западной части губы (более 70 м) биомасса M. оkhotensis не превышала 100 мг/м3 и была представлена старшими копеподитными стадиями. Вид, по мере перехода к старшим стадиям мигрирует на глубину и выносится с западной выносящей ветвью Ямского течения в надшельфовую зону Притауйского пром. района. Здесь M. оkhotensis участвует в формировании зоны высокой биологической продуктивности являясь важным компонентом в питании пелагических рыб.
N. plumchrus. Незначительное присутствие его в планктоне (от 1 до 50 мг/м3) отмечались практически на всей акватории Тауйской губы. Возрастная структура была представлена копеподитами II и III стадий. Однако в западной части губы рачок образовывал значительные скопления восточнее о. Спафарьева. Численность этого рачка составила 382 экз./м3, а биомасса до 670 мг/м3. Здесь возрастная структура популяции N. plumchrus была представлена особями IV и V копеподитных стадий (74%), а также взрослыми рачками – 26%. Все они содержали небольшое количество жира. Концентрация рачков старших стадий в западной части губы свидетельствует о выносе их по мере взросления в надшельфовую зону Притауйского пром. района.
Calanus glacialis. На акватории Тауйской губы биомасса рачка изменялась в пределах от 1 до 50 мг/м3. Максимальные скопления вида отмечались восточнее о-ва. Спафарьева на выходе из губы, где биомасса его достигала 69 мг/м3, а численность - 1736 экз./м3. На остальной акватории половозрелые формы встречались в единичных экземплярах (от 1 до 46), но, в основном, это были мелкие формы. Значение C. glacialis как и N. plumchrus, крайне велико, т.к. они служат основной пищей пелагических рыб.
Зоопланктонное сообщество Тауйской губы активно участвует в формировании зоны высокой биологической продуктивности в прилегающем участке шельфа северной части Охотского моря. При участии западной выносящей ветви Ямского течения происходит вынос зоопланктона из губы в прилегающую губе акваторию, о чем свидетельствует высокая концентрация биомассы отдельных видов (М. okhotensis, N. plumchrus, C. glacialis) в западной части на выходе из губы. По мере взросления копеподиты M. okhotensis, C. glacialis, N. plumchrus, личинки эвфаузиид, декапод мигрируют и в дальнейшем становятся участниками надшельфового сообщества прилегающей к Тауйской губе акватории (рис.6).
В составе сообщества Тауйской губы также на отдельных станциях (глубина более 70 метров) наблюдались единичные экземпляры видов присущих сообществам открытых вод. Их распространение, четко коррелировало с гидрологическими данными, определяющими воды Ямского течения. Это Eucalanus bungii, С. cristatus, T. libellula (рис.7). Стоит также

Рис. 6. Распределение биомассы C. glacialis в Тауйской губе в весенне-летний период (1 - направление западной выносящей ветви Ямского течения, 2 – локализация зоны высокой биологической продуктивности).

Рис. 7. Распределение биомассы E. bungii в Тауйской губе в весенне-летний период (стрелкой показано направление восточной приносящей ветви Ямского течения).
отметить, что вышеперечисленные виды встречались в Тауйской губе только на взрослых стадиях, что подтверждает утверждение об их привнесении с восточной ветвью Ямского течения.
Euphausiaceae. Общая биомасса эвфаузиевых (личинок и взрослых форм) была достаточно велика и составляла в среднем 140,56 мг/м3, численность 46685 экз./м3. Основные скопления эвфаузиид наблюдались в прибрежной зоне п-ова Старицкого, где и отмечался максимальный прогрев вод до 11,5С, что способствовало интенсивному размножению и развитию.
Amphipoda. Бокоплавы были представлены гипериидами T. libellula и T.pacifica. Биомасса гипериид варьировала от 5 до 16 мг/м, а их доля в планктоне составляла от 0,6 до 1,2 %. Распространение T. libellula на акватории губы ограничивалось зоной прогрева воды - до 7С. Зоны распределения этого вида на акватории достаточно четко отражали границы влияния затоков холодных водных масс с юга (Ямское течение) и теплых прогретых вод с северной части Тауйской губы. Биомасса T. libellula варьировала от 4 до 32 мг/м, доля в планктоне колебалась от 0,5 до 4,3 %. Decapoda. Основу биомассы крупной фракции в весенне-летний период составляли молодь и личинки десятиногих ракообразных. Встречаемость молоди десятиногих раков и их личинок была достаточно высокой (до 74%), большая их часть присутствовала, в основном в мелководной зоне Тауйской губы, составляя в среднем 173 экз./м, формируя 62,7 % биомассы крупной фракции по всему району и до 90 % - в восточной половине губы; в 2005 году биомасса колебалась от 1,2 до 285 мг/м, составляя в среднем 30,8 мг/м, около 12,0 % от биомассы крупной фракции по всему району и до 85% - в восточной половине губы. Максимальные концентрации молоди десятиногих ракообразных и их распределение были приурочены к фронтальной зоне к району схождения распресненных вод Тауйской губы и вод Ямского течения и по его северной периферии. Отмечено, что более поздние стадии развития декапод находились на большей глубине, чем младшие стадии.
3.3. Сезонная динамика зоопланктонного сообщества
Тауйской губы и прилегающего участка Охотского моря
Доля голопланктона прибрежного сообщества Тауйской губы составляла от 53% в июле и до 90 % в октябре. В течение всего периода исследований среди голопланктона доминировала группа Copepoda. В июне она составила 34,8%, 371,7 мг/м3, в июле доля от общей биомассы снижается и составляет 24,1%, 146 мг/м3 (рис. 8.).
Меропланктон составляет в июне 24,4%, в июле его доля возрастает до 46,2%, а осенью составляет не больше 11%. В июне 19% составляют личинки десятиногих ракообразных, Decapoda - 210 мг/м3, 4% науплии и циприсовидные личинки усоногих раков (Cirripedia) 43,3 мг/м3; в июле доля Decapoda возрастает до 44,5% и составляет уже 269,8 мг/м3, биомасса Cirripedia снижается до 5 мг/м3. В октябре Decapoda также составляли основу меропланктона – 10% от общей биомассы зоопланктона, 81,7 мг/м3. В июне появляются эхино- и офиуплютеусы (Echinodermata), в июле их доля возрастает.

Рис. 8. Сезонные соотношения различных групп зоопланктона
в Тауйской губе и прилегающем участке акватории
В июне, наблюдается массовое размножение холоднолюбивых видов, что характеризует этот период как конец биологической весны. Из Copepoda это M. okhotensis, C.glacialis, P. minutus. В большом количестве в планктоне присутствуют их науплиусы и молодь, численность которых очень быстро уже в июле достигает максимума. В это же время в планктоне встречаются теплолюбивые Copepoda: C. abdominalis, C. bipinata и другие, а у эврибионтных M. rosea и O. similis наблюдаются массовые преднерестовые скопления. Большое количество самок с яйцевыми мешками. Ниже перечисленные процессы, наблюдаемые в июне, могут служить надежными критериями перехода от биологической весны к лету в Тауйской губе:
а) массовое размножение Cirripedia, большое количество науплиусов и циприсовидных личинок;
б) большое количество молоди и науплиев C. glacialis;
в) большое количество молоди P. minutus;
г) наличие большой биомассы яйценосных самок M.rosea.
В июле появляются ювенильные стадии теплолюбивого C. abdominalis. В планктоне появляются Cladocera – P. leucartii и E. nordmanni. В августе в разгар лета наблюдается исчезновение в планктоне молоди C. glacialis; максимум численности Cladocera и максимум численности планктонных личинок моллюсков Bivalvia; наблюдается также исчезновение личинок Echinodermata и науплиусов Cirripedia.
Касаясь вопроса сезонной изменчивости в видовом составе планктона, следует отметить, что весной достигают высокой численности (до конца июня) умеренно-холодноводные виды, а в теплые годы уже в июле численность их сокращается, и в планктоне появляются более тепловодные виды E. nordmannii, P. leucartii и веслоногие ракообразные прибрежного тепловодного комплекса: C. abdominalis,, C. bipinata. На стабильном уровне представлены личинки полихет - 3,5 мг/м.
3.4. Межгодовая динамика видовой структуры зоопланктона
Межгодовые изменения зоопланктона Тауйской губы хорошо прослеживается не только по общим количественным показателям в целом, но и по соотношению численности и биомассы отдельных групп и видов. Сравнение динамики развития зоопланктона в весенне-летний период в Тауйской губе в теплые годы 1986, 1989 (Афанасьев, Михайлов, 1994) и 2004, 2005, 2006 гг. показывает, что общий ход развития планктона весьма сходен. Однако общие значения биомассы зоопланктона в 2004-2006 гг. были выше, составляя в среднем до 953 мг/м, в сравнении с биомассой в ранее исследованные годы (470-770 мг/м в 1986 и 1989 гг.).
В различные годы исследований Тауйской губы в весенне-летний период, численности и по биомассе среди голопланктона преобладали две таксономические группы Copepoda и Euphausiidae. Численность этих групп ракообразных претерпевают значительные межгодовые колебания (рис. 9).
Copepoda. В 1989 году доля в общей биомассе группы Copepoda составила 94 %. Доминируют M. okhotensis, N. plumchrus, A. longiremis, P. minutus, C. glacialis.
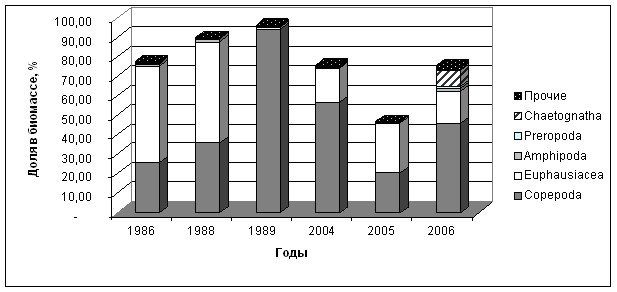
Рис. 9. Межгодовая динамика биомассы различных групп
голопланктона в Тауйской губе в весенне-летний период
(1986-1989 гг. - по Н.Н. Афанасьеву, В.И. Михайлову, 1994)
Копеподы в июле 1989 г. составляли - 726,5 мг/м, в июне 2004 г. - 539,1 мг/м, а в первой декаде июля 2005 г. отмечалось заметное снижение концентрации копепод до 203,76 мг/м. В первую очередь это снижение отразилось на С. glacialis, M. okhotensis, O. similis и N. plumchrus. В годы более низких температур численность копепод – P. minutus и O. similis уменьшалась (в июне 1988 г. P. minutus - до 825 экз./м, а O. similis до 243 экз./м) и увеличивалась в теплые годы. Резкое снижение биомассы в 2005 г. объясняется, поздним нерестом копепод (C. glacialis и M. okhotensis) в конце мая, что подтверждается наличием в планктоне большого количества яиц и науплий.
Euphausiaceae. Доля в биомассе составляла от 14 до 50 % от общей биомассы зоопланктона. В теплые годы наблюдалось увеличение биомассы и численности. Численность Th. raschii в теплые годы колебалась от 65103 экз./м - в 1986 г. а в холодный 1988 г численность эвфаузиид составляла всего 30223 экз./м. В последние годы наблюдается увеличение биомассы и численности эвфаузиид, до 46689 экз./м - в 2004 г. и 64560 экз./м в 2005 г.
Среди представителей меропланктона в весенне-летний период в зоопланктонном сообществе Тауйской губы доминируют личинки Decapoda и Cirripedia, составляя, соответственно, в разные годы от 1 до 50% и от 3 до 17 % общей биомассы зоопланктона. На долю меропланктона приходится от 3 до 52 % в разные годы исследований. На долю Polychaeta и Bivalvia приходится менее 1 %.
3.5. Значение зоопланктона в питании промысловых
рыб в северной части Охотского моря
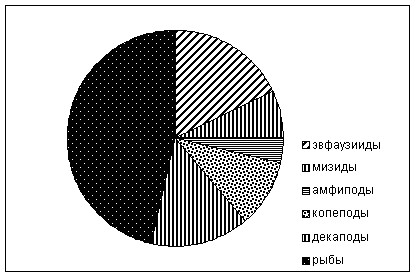
В период исследований чётко выраженной суточной ритмики питания минтая за исключением молоди не было выражено. Годовики минтая в течение суток интенсивно питались и в желудках постоянно присутствовала свежесъеденная пища (ИНЖ, 108о/ооо). Основу пищи составляли эвфаузииды (70%), в меньшей степени копеподы (25,6%). В рационе присутствовали амфиподы (4%) и мизиды (0,5%). Среди эвфаузиид преобладали T. rashii, из копепод - M. okhotensis. Максимум наполнения, за счёт интенсивного питания эвфаузиидами, приходился на ночные часы. Минтай размерной группы 17-30 см, в которую входят рыбы 2-3-х годовалого возраста, встречался в уловах реже годовиков. ИНЖ молоди минтая (17-30 см) была значительно ниже, чем у годовиков – 17,4о/ооо. Основу рациона составляли эвфаузииды (62,9%)и копеподы (28,5%)(рис. 10).
Основу уловов минтая размерной группы 30-40 см (мода 36-37 см) составляли 4-х годовалые неполовозрелые особи во II стадии развития гонад. ИНЖ довольно низкая, СПР - менее 1%. Рацион минтая данной размерной группы характеризуется довольно широким спектром, его основа – эвфаузииды (37,5%), молодь рыб (34,6) и копеподы (14,6%). В питании минтая этой размерной группы появляются декаподы (4,4%). ИНЖ была невысокой 25,2 о/ооо. По составу пищи минтай размерной группы 40-50 см отличался от предыдущей увеличением в питании доли рыб (44,5%) и декапод (14,2%). ИНЖ была равна – 14,3 о/ооо.
В питании минтая длиной более 50 см преобладали рыбы (63,9%), декаподы (21,8%) и эвфаузииды (13,2%). Из рыб доминировали минтай мойва и серебрянка. В целом ИНЖ крупноразмерного минтая, была, как и у предыдущих групп, относительно низкой – 41,9/ооо. Максимум наполнения желудков приходился на период с 21 до 00 ч.


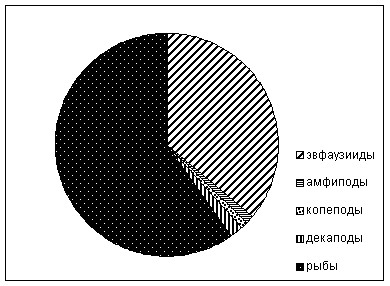

Рацион питания сельди в нерестовый период не отличался большим разнообразием. Основную долю, до 97,1% от массы пищевого комка, у исследованных рыб в среднем составлял вид T. raschii (Euphausiidae). Из копепод обнаружено три вида: M. okhotensis, C. glacialis, N. plumchrus. Было отмечено, что 56,7% желудков были пустые либо с незначительным наполнением до 20о/ооо, 26,6 % имели наполнение более 150 о/ооо, остальные, от 20 до 150о/ооо. ИНЖ был невысокий – 50,6о/ооо. Следует отметить, что в среднем самый высокий индекс наполнения желудков наблюдался в пробах, отобранных из утренних уловов.
Основу питания нагульной сельди в размерных группах 20-30 см, составляли копеподы – 85,7%. Наибольшее значение в питании нагульной сельди имеют M. okhotensis, их доля составляет 64,6%; вторым по значимости в питании является N. plumchrus – 20,2%, а третьим – N. cristatus (0,8%). На долю эвфаузиид приходится 9,3%, амфипод 5%.
4. Заключение
Зоопланктонное сообщество Тауйской губы является гетерогенным по своему составу и происхождению. Наряду с местной фауной (прибрежные и надшельфовые виды) в нем присутствуют элементы сообщества открытых вод привносимые с водами Ямского течения. Выявлено 30 видов в зоопланктоценозе Тауйской губы. К прибрежному сообществу относятся виды A. longiremis, E. nordmanni, C. abdominalis, P. leucartii, личинки Polychaeta, Bivalvia, Echinodermata, Gastropoda. Типичными видами надшельфового сообщества являются C. glacialis, T. raschii, M. okhotensis, N. plumchrus. Виды открытых сообществ, привносимые Ямским течением – E. bungii, T. pacifica, C. cristatus.
В результате многолетних наблюдений планктоценоза Тауйской губы была выявлена сезонность в развитии зоопланктона и прослежены межгодовые колебания его численности и биомассы. В течение всего периода исследований в планктоне по численности и биомассе преобладают две таксономические группы: Copepoda и Euphausiidae. Общая характерная особенность зоопланктона Тауйской губы является преобладание неритических видов над океаническими и умеренно – холодноводных видов над холодноводными, что обусловлено теплым режимом вод в годы исследований. Получены картины горизонтального распределения около 20 видов копепод, эвфаузиид и гипериид, которые формируют основу численности и биомассы зоопланктона Тауйской губы и прилегающего участка Охотского моря. Из числа исследованных видов выделены несколько индикаторных: в системе вод Ямского течения это прежде всего E. bungii и T. libellula.
В зоопланктонном сообществе Тауйской губы представлен один ярко выраженный всесезонный доминирующий вид – T. raschii, типичный фитофаг. Примечательно, что типичные хищные формы зоопланктона (хетогнаты, гиперииды) в данном сообществе практически отсутствуют.
Зоопланктонное сообщество Тауйской губы активно участвует в формировании зоны высокой биологической продуктивности в прилегающем к губе участке Охотского моря. Нерест массовых видов зоопланктонного сообщества Тауйской губы M. okhotensis, C. glacialis, N. plumchrus, многих видов декапод, эвфаузиид, проходит в прибрежной зоне, в мелководных и хорошо прогреваемых богатых биогенами заливах Тауйской губы. По мере взросления виды выносятся с западной выносящей ветвью Ямского течения в прилегающий к Тауйской губе участок Охотского моря. Затем здесь они участвуют в формировании зоны высокой биологической продуктивности Притауйского промыслового района.
В настоящее время акватория Тауйской губы мало затронута хозяйственной деятельностью человека. В связи с этим полученные данные по биоразнообразию зоопланктонного сообщества, закономерностям его распределения можно рассматривать как близкие к естественным, не трансформированным воздействием антропогенной деятельности. Это может явиться объективной основой для организации постоянного мониторинга экосистемы Тауйской губы, особенно в районах потенциального экологического риска. К таким относятся участки поверхностного месторождения бурого угля в бассейне реки Ола и на побережье залива Одян.
Выводы
1. Фитопланктон в Тауйской губе представлен холодноводными формами неритического комплекса: Coscinodicus, Thalassiozira, Rhizosolenia и Chaetoceros. Средняя биомасса фитопланктона в Тауйской губе в весенне-летний период составила 1087 мг/м. Квазистационарная зона высокой биомассы микроводорослей (от 500 до 1000 мг/м3) формируется между о. Завъялова и п-вом Старицкого, где осуществлялся приток более холодных субарктических вод в системе приносящей ветви Ямского течения.
2. Зоопланктонное сообщество Тауйской губы, представленное 30 видами, включает автохтонный комплекс прибрежных и надшельфовых видов и аллохтонные виды открытых вод, привносимые водами восточной ветви Ямского течения. Характерными видами прибрежного комплекса являются C. abdominalis, P. leucartii, E. nordmanni и личиночные стадии Euphausiacea, Decapoda, Echinodermata, Cirripedia, Polychaeta. К надшельфовым видам относятся T. raschii, C.glacialis, M. okhotensis. Привносимые виды - E. bungii, C. cristatus, T. libellula.
3. В сезонном аспекте в прибрежном зоопланктоценозе Тауйской губы резко выражена изменчивость соотношения голопланктонных и меропланктонных форм. В 2005-2006 гг. доля голопланктона по биомассе составляет 65% в июне, снижается до 53% в июле и возрастает до 90 % в октябре. При этом доля меропланктона возрастает от 24,4% в июне до 46,2% в июле и снижается осенью до 11%. Только в июне заметна доля науплий и циприсовидных личинок Cirripedia до 4% (43,3 мг/м3).
4. Заметные межгодовые изменения видового состава зоопланктона Тауйской губы отсутствуют, постоянные доминанты - T. rashii (Euphausiacea) и M. okhotensis (Copepoda). В годы позднего освобождения ото льда (2005 год) отмечается заметное снижение биомассы копепод до 203,76 мг/м, связанное с поздним нерестом С. glacialis и M. okhotensis. В это время в планктоне наблюдается большое количество их яиц и науплий. Характерны резкие изменения доли личинок Decapoda в зоопланктоне от 1% до 50%. При этом в целом можно констатировать постепенное увеличение их доли в зоопланктоне: 3% в 1989 г., 17,5% в 2004, 48% - в 2005 г., что может свидетельствовать о увеличении численности Decapoda в донных сообществах северной части Охотского моря.
5. Зоопланктонное сообщество Тауйской губы участвует в формировании зоны высокой биологической продуктивности Притауйского промыслового района Охотского моря. Зоопланктон выносится из губы в прилегающую акваторию западной ветвью Ямского течения. Об этом свидетельствует высокая концентрация биомассы отдельных видов (М. okhotensis, N. plumchrus, C. glacialis) в западной части на выходе из губы.
6. В период исследований чётко выраженной суточной ритмики питания минтая за исключением молоди выражено не было. Годовики минтая в течение суток интенсивно питались. Основу пищи составляли T. rashii (70%) и M. okhotensis (25,6%). Максимум наполнения, за счёт интенсивного питания эвфаузиидами, приходился на ночные часы. В рационе крупноразмерного минтая преобладает молодь рыб (44,5%) и декаподы (14,2%). В целом ИНЖ крупноразмерного минтая, была, относительно низкой – 41,9/ооо.
7. Основу рациона сельди в нерестовый период составляли представители двух таксономических групп – Euphausiaceae и Copepoda, в небольших количествах встречались представители Hyperiidae, Mysidae и личинки Decapoda. Значительную часть, до 97,1% от массы пищевого комка, составляла эвфаузиида T. raschii, а практически вся остальная доля приходилась на копепод M. okhotensis, C. glacialis, N. plumchrus.
Список работ, опубликованных
по теме диссертации
(Издания, рекомендованные ВАК, отмечены звездочкой)
- Вакатов А.В., Кащенко Е.В. Питание сельди Арманского побережья Тауйской губы в мае-июне 2005 года // Материалы VI научной конференции «Сохранение биоразнообразия Камчатки и прилегающих морей». Петропавловск-Камчатский, 2005. С. 233-236.
- Вакатов А.В. Экологическая характеристика зоопланктона залива Одян северной части Охотского моря в июле 2005 года // Тезисы X международной Пущинской школы-конференции «Биология – наука XXI века», Пущино, 2006. С. 241-242.
- *Вакатов А.В. Состояние планктонного сообщества Тауйской губы Охотского моря по результатам комплексной съемки в весенне-летний период 2005 года // Учен. зап. Казанск. гос. ун-та, 2007. - Т. 149, кн. 1. - С. 89-102.
- Вакатов А.В. Межгодовая динамика зоопланктонного сообщества Тауйской губы Охотского моря // Тезисы XI международной Пущинской школы-конференции «Биология – наука XXI века», Пущино, 2007. С. 202-203.
- Vakatov A.V. Zooplankton of Taui Bay of Sea of Okhotsk as indicator of ecosystem stability in places of a prospective coal mining // Abstracts of ECSA-42 simposium «Estuarine ecosystems: structure, function and management». Kaliningrad, 2007. P. 110-111.
- *Вакатов А.В. Фоновое состояние зоопланктона Тауйской губы северной части Охотского моря в весенне-летний период 2004 г. // Совр. пробл. науки и образования. М.: РАЕ, 2007. № 5. С. 82-93.
- *Вакатов А.В. Биомасса и распределение зоопланктона по результатам комплексной съемки в Тауйской губе северной части Охотского моря в июле 2006 года // Учен. зап. Казанск. гос. ун-та, 2007. Т. 149, кн. 3. С. 242-246.
- Вакатов А.В. Экологическая характеристика видового состава планктона северной части Охотского моря // Сборник научных трудов Всероссийской конференции «Эколого-биологические проблемы вод и биоресурсов; пути решения», Ульяновск, 2007. C. 225-230.
- Vakatov A.V. The seasonal dynamics of zooplankton of Taui Bay of Sea of Okhotsk // Abstracts of First Student Conference on Modern Biology «Bio-News», Kazan, 2008. P. 15.
- *Вакатов А.В., Смирнов А.А., Сабиров Р.М. Значение зоопланктона в питании промысловых рыб в северной части Охотского моря // Учен. зап. Казанск. гос. ун-та, 2008. Т. 150, кн. 3. 12 с. (в печати).