

Агроэкологическое обоснование использования энтомофагов в защите злаковых культур от тлей в лесостепи западной сибири
На правах рукописи
БОКИНА
Ирина Геннадьевна
АГРОЭКОЛОГИЧЕСКОЕ ОБОСНОВАНИЕ ИСПОЛЬЗОВАНИЯ
ЭНТОМОФАГОВ В ЗАЩИТЕ ЗЛАКОВЫХ КУЛЬТУР ОТ ТЛЕЙ
В ЛЕСОСТЕПИ ЗАПАДНОЙ СИБИРИ
06.01.11 – защита растений
А в т о р е ф е р а т
на соискание ученой степени доктора
биологических наук
Кинель – 2009
Работа выполнена в Государственном научном учреждении Сибирский научно-исследовательский институт земледелия и химизации сельского хозяйства (ГНУ СибНИИЗХим) СО Россельхозакадемии
Научный консультант: доктор биологических наук, профессор,
заслуженный деятель науки РФ
Власенко Наталия Григорьевна
Официальные оппоненты: доктор биологических наук, профессор,
заслуженный деятель науки РФ
Каплин Владимир Григорьевич
доктор сельскохозяйственных наук, профессор
Еськов Иван Дмитриевич
доктор биологических наук,
Легалов Андрей Александрович
Ведущая организация: Новосибирский государственный аграрный
университет
Защита диссертации состоится « 20 » октября 2009 года в 1000 часов на заседании диссертационного совета ДМ.220.058.01 при ФГОУ ВПО «Самарская государственная сельскохозяйственная академия».
Адрес: 446442, Самарская область, г. Кинель-4, п. Усть-Кинельский, ул. Учебная, 1, диссертационный совет.
С диссертацией можно ознакомиться в библиотеке ФГОУ ВПО «Самарская государственная сельскохозяйственная академия».
Автореферат разослан « » ___________ 2009 г.
Размещен на сайте ВАК www.vak.ed.gov.ru
Ученый секретарь
диссертационного совета,
кандидат биологических наук,
профессор Г.К. Марковская
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы. Среди 11 территориально-экономических зон Российской Федерации агропромышленный комплекс Западной Сибири ежегодно производит около 10 млн. т зерна или 10,4% общероссийского (Храмцов, 2004). Факторами, лимитирующими урожайность злаковых культур, наряду с болезнями и сорняками, являются вредные насекомые. По оценкам различных авторов, потери урожая зерновых от таких специализированных вредителей, как хлебные блошки, злаковые мухи, сосущие насекомые в Западной Сибири на рядовых посевах достигают 10-23% (Горбунов и др., 1984; Гарбар, 1987), а на высокоурожайных могут быть еще выше (Коробов, 1996). В различных регионах нашей страны и за ее пределами одними из основных вредителей зерновых культур являются злаковые тли (Танский, 1972; Хохлов и др., 2000; Берим, 2002; Сагитов, 2004; Vickermann, Wratten, 1979; Wiktelius, 1982, 1984 и др.). В Западной Сибири численность тлей по годам сильно колеблется, вспышки их размножения наблюдаются в отдельные годы (Иванов и др., 1965).
Ориентация растениеводства на экологическую безопасность, ресурсо-энергосбережение и рентабельность предполагает снижение пестицидной нагрузки в агробиогеоценозах, в первую очередь за счет биологизации и экологизации продукционных и средообразующих функций агроэкосистем и агроландшафтов. Среди факторов биологизации важное место занимают биологические методы и средства защиты растений (Жученко, 1993, 1997). Одним из направлений биометода является использование энтомофагов, которые наряду с другими энтомопатогенными организмами, осуществляют регуляцию численности фитофагов, поддерживая биоценотическое равновесие в агроэкосистемах. С этой точки зрения тли вызывают большой интерес не только как вредители, но и как источник питания обширного ряда хищных и паразитических насекомых, составляющих основу видового разнообразия энтомофауны агроценозов (Воронин, Воронина, 1996). Каждая природно-хозяйственная зона характеризуется своеобразными экологическими условиями для вредных и полезных организмов, поэтому их численность, видовое соотношение, степень влияния энтомофагов на популяции вредителей в этих зонах неодинаков.
Для реализации биологического потенциала агроэкосистем необходимо знание реакции энтомофагов на проведение тех или иных агротехнических мероприятий. Создавая благоприятные условия для развития природных энтомофагов и других полезных видов, агротехнические приемы способны усилить биологический метод, обеспечить оптимизацию биоценотической структуры агробиоценозов и охрану окружающей среды (Павлов, 1981; Сусидко, 1997).
В связи с вышеизложенным вопросы изучения злаковых тлей и трофически связанных с ними хищных и паразитических насекомых, а также влияния на них различных агротехнических приемов возделывания зерновых культур приобретают особую актуальность.
Цель исследований – изучение энтомофагов злаковых тлей в агроценозах зерновых культур в условиях лесостепи Западной Сибири, влияния на них основных элементов агрокомплекса возделывания яровой пшеницы с целью сохранения биоценотических механизмов регуляции, стабильной экологической ситуации в агроэкосистемах.
Основные задачи:
уточнить видовой состав, эколого-биологические характеристики, многолетнюю и сезонную динамики численности доминирующих видов энтомофагов злаковых тлей в агроценозах озимой ржи, яровой пшеницы и овса;
уточнить роль энтомофагов в снижении численности злаковых тлей;
установить роль лесополос, дикорастущей растительности и многолетних трав в повышении численности хищных и паразитических энтомофагов;
выявить влияние основных элементов агрокомплекса (предшественников, системы основной обработки почвы и средств химизации) на численность злаковых тлей и их энтомофагов.
Научная новизна. В условиях лесостепи Западной Сибири уточнен видовой состав хищных и паразитических насекомых яруса травостоя озимой ржи, яровой пшеницы и овса, трофически связанных со злаковыми тлями, насчитывающий к настоящему времени 134 вида (89 энтомофагов и 45 паразитов энтомофагов). Отмечена идентичность видового состава в различных агроландшафтных районах Новосибирской области. Получены новые сведения о биологических особенностях многих видов энтомофагов, многолетней и сезонной динамике их численности в агроценозах зерновых культур, стациальному распределению. Показана эффективность энтомофагов в снижении плотности популяций злаковых тлей. Впервые установлена роль лесополос, дикорастущей растительности, многолетних и двулетних трав в повышении численности хищников и паразитов. Впервые изучено влияние системы основной обработки почвы, средств химизации, технологий возделывания, предшественников на численность злаковых тлей, хищных и паразитических энтомофагов в травостое яровой пшеницы.
Основные положения, выносимые на защиту:
формирование экологически стабильных агроэкосистем и сохранение в них биоценотических механизмов регуляции должны основываться на изучении видового состава, трофических связей, биологических особенностей хищных и паразитических насекомых в агроценозах сельскохозяйственных культур;
значительную роль в регулировании численности злаковых тлей играет комплекс энтомофагов яруса травостоя;
применение удобрений и химических средств защиты растений, регламентируемых технологиями возделывания, оказывая влияние на отдельные группы вредных и полезных членистоногих, не приводит к нарушению естественной регуляции в агроценозах зерновых, не нарушает их целостности;
мероприятия, направленные на создание оптимизированных по видовому составу сетей лесополос, охрана цветущего дикорастущего разнотравья, возделывание многолетних трав способствуют повышению численности хищных и паразитических насекомых, стабильности экологической ситуации в агроэкосистемах.
Практическая и теоретическая значимость. На основе изучения многолетней и сезонной динамики численности энтомофагов, их биологии, синхронности с развитием тлей, прожорливости хищников, зараженности тлей паразитами определены доминирующие и наиболее эффективные виды афидофагов. Установлены уровни численности хищников и жертвы, при которых в агроценозах озимой ржи, яровой пшеницы, овса поддерживается биоценотическое равновесие, то есть численность вредителей сдерживается на хозяйственно-неощутимом уровне, либо при достижении порога экономической вредоносности снижается до такого уровня в течение нескольких дней, при этом проведение химических обработок не требуется. Знание влияния на злаковых тлей и их энтомофагов основных элементов агрокомплекса дает возможность поддерживать численность энтомофагов на определенном уровне и сохранять стабильность экологической ситуации в агроценозах сельскохозяйственных культур. Изучение распределения энтомофагов на окружающих зерновые культуры стациях позволяет проводить мероприятия по повышению их численности и эффективности. Результаты исследований в дальнейшем планируется использоваться при составлении методических рекомендаций, учебных пособий для студентов и аспирантов вузов.
Апробация работы. По теме исследований опубликовано 25 работ, в том числе 13 работ в изданиях, рекомендованных ВАК РФ, а также монография «Злаковые тли и их энтомофаги в лесостепи Западной Сибири». Материалы диссертации докладывались на VI региональной конференции молодых ученых Сибири и Дальнего Востока «Роль научно-технического прогресса в реализации продовольственной программы Сибири и Дальнего Востока», Новосибирск (1987 г.); на Всесоюзной конференции молодых ученых «Актуальные вопросы интенсификации сельского хозяйства», Новосибирск (1989 г.); VII совещании энтомологов Сибири, Новосибирск (1989 г.); X съезде Всесоюзного энтомологического общества, Ленинград (1989 г.); конференции молодых ученых «Экологические проблемы защиты растений», Ленинград (1990 г.); XIII съезде Российского энтомологического общества, Краснодар (2007 г.); III Международной научно-практической конференции «Аграрная наука – сельскому хозяйству», Барнаул (2008 г.); Международной научно-практической конференции «Современные средства, методы и технологии защиты растений», Новосибирск (2008 г.), XII Международной научно-практической конференции «Аграрная наука – сельскохозяйственному производству Казахстана, Сибири и Монголии», Шымкент (2009 г.).
Личный вклад автора. Выбор направления научного поиска, сбор, обработка и анализ исходной информации. Работа является обобщением результатов 15-ти летних исследований, выполненных лично автором в соответствии с тематическим планом выполнения НИР ГНУ СибНИИЗХим СО Россельхозакадемии, согласно государственных программ Россельхозакадемии: по заданию 03.H «Разработать научные принципы построения севооборотов, обеспечивающих стабильное функционирование равнинных ландшафтов северной лесостепи Западной Сибири на выщелоченных черноземах с помощью сбалансированного применения биологических, химических и агротехнических средств регулирования продукционного процесса в агроценозах», по заданию ГКНТ 03.01.H1. «Изучить основные параметры фитосанитарного состояния посевов зерновых в севооборотах и подобрать ассортимент эффективных экологически безопасных средств защиты зерновых, рапса, сои и плодово-ягодных культур от вредных организмов», по заданию ГКНТ-2-256 «Разработать модели экологически безопасной интегрированной защиты основных сельскохозяйственных культур от вредителей, болезней и сорняков и дать эколого-экономическую оценку в различных почвенно-климатических зонах Сибири», по заданию (02.05.01)05.Н1 «Разработать экологически безопасные элементы технологий комплексного применения удобрений, химических средств защиты растений, регуляторов роста и биопрепаратов в адаптивно-ландшафтном земледелии лесостепи Приобъя с целью увеличения продуктивности культур и повышения окупаемости удобрений».
Структура и объем диссертации. Диссертационная работа изложена на 326 страницах машинописного текста и состоит из введения, 11 глав, выводов, предложений производству, списка использованной литературы, включающего 678 наименований, в том числе 161 – на иностранных языках, 2 приложений. Работа иллюстрирована 36 таблицами и 42 рисунками.
СОДЕРЖАНИЕ РАБОТЫ
1. Злаковые тли и их энтомофаги: история изучения
Первая глава посвящена аналитическому обзору отечественной и зарубежной литературы по изучаемой проблеме. Приведены сведения по эколого- фаунистической характеристике и вредоносности злаковых тлей, численности, видовому составу и роли хищных и паразитических энтомофагов яруса травостоя в снижении плотности популяций вредителей, влиянию на них элементов агрокомплекса возделывания зерновых. Проанализированная информация позволяет сделать вывод, что некоторые из этих вопросов в лесостепи Западной Сибири изучены слабо, исследования по большей части их в условиях региона отсутствуют.
2. Объекты, условия и методы проведения исследований
2.1. Объекты исследований
В связи с целью и задачами исследований в качестве объектов исследований были взяты злаковые тли и их энтомофаги.
До 95% суммарной численности злаковых тлей, обитающих в травостое зерновых, составляют большая злаковая Sitobion avenae F. и черемухово-злаковая тли Rhopalosiphum padi L..
Большая злаковая тля – однодомный вид. Зимует в фазе яйца на дикорастущих злаках и всходах озимой ржи, с внутренней стороны нижней части листьев, в пазухах листьев по 1-2 на растении. Личинки отрождаются во второй декаде мая, приблизительно через 20 дней после начала возобновления вегетации растений ржи. Максимальная численность большой злаковой тли в агроценозе озимой ржи наблюдается в первой – начале второй декады июля в конце цветения растений – начале формирования зерновок и достигает 780 особей/м. В это время тля заселяет до 49% растений, на один стебель в среднем приходится до 1,1 особи. На яровых культурах максимум плотности популяции тли (до 1930 особей/м) наблюдается в третьей декаде июля – начале августа в фазы формирования зерновки – молочной спелости зерна, тля заселяет до 58% растений, на один стебель в среднем приходится до 4,5 особей.
Черемухово-злаковая тля – вид двудомный. В Западной Сибири зимует в фазе яйца с нижней стороны оснований почек, в трещинах коры ветвей и ствола черемухи. Отрождение личинок из зимующих на черемухе яиц наблюдается в конце апреля – начале мая в период набухания почек. С конца мая – второй декады июня начинается миграция крылатых особей тли на посевы зерновых. К концу июня вредитель полностью перелетает с черемухи на злаковые растения. На посевах озимой ржи численность тли достигает пика (до 1480 особей/м) в первой – второй декаде июля в фазы цветения – формирования зерновки, тля заселяет до 9,1% растений, на один стебель в среднем приходится до 1,9 особей. На яровых культурах максимальная плотность популяции тли наблюдается в третьей декаде июля – начале августа, редко в более ранние сроки, в фазы колошения пшеницы, выметывания метелок овса – формирования зерновки – молочной спелости зерна и составляет до 7290 особей/м, тля заселяет до 73,6% растений, на один стебель приходится в среднем до 14,3 особей.
Снижение численности злаковых тлей на полях наблюдается через 1-2 недели после ее пика, чему способствуют усиление активности афидофагов, уменьшение содержания питательных веществ к фазе восковой спелости зерна.
В качестве объектов исследования были взяты также массовые виды энтомофагов, трофически связанных со злаковыми тлями.
2.2. Методы проведения исследований
Исследования проводили в 1985-1993, 1998-2000 и 2005-2007 гг. на производственных посевах зерновых культур и опытных полях Сибирского НИИ земледелия и химизации в ОПХ «Элитное» Новосибирского района (центрально-лесостепной Приобский агроландшафтный район), 5 хозяйствах Краснозерского района (южно-лесостепной Барабинский) и в ОПХ «Кочковское» Кочковского района (центрально-лесостепной Верхнекарасукский) Новосибирской области, расположенной в лесостепи Западной Сибири.
Видовой состав, сезонную динамику численности злаковых тлей и их энтомофагов, их биологические особенности, структуру популяций, зараженность тлей паразитическими наездниками, эффективность хищных энтомофагов изучали в агроценозах озимой ржи (сорт Короткостебельная 69), яровой пшеницы (Новосибирская 81), овса (Сир 4, в 1985 г. – Белозерный). Основные методы учета – подсчет и наблюдение за насекомыми во всех фазах их развития на 250 модельных растениях (Зубков, 1980; Танский и др., 1998), кошения стандартным энтомологическим сачком (Кожанчиков, 1961; Палий, 1970; Фасулати, 1971), ручной сбор и содержание в лабораторных условиях живых и мумифицированных тлей, энтомофагов. Для изучения стациального распределения энтомофагов, оценки роли лесополос и окружающей растительности в повышении численности хищных и паразитических членистоногих еженедельно кошения делали в лесополосах около зерновых культур, на полянах дикорастущего разнотравья, а также на многолетних и двулетних травах, возделываемых в хозяйстве: люцерне, доннике, эспарцете, клевере, костреце безостом. Помимо посевов зерновых еженедельно практически в течение всей вегетации наблюдали за развитием черемухово-злаковой тли и ее энтомофагов на черемухе.
Весь собранный и выведенный энтомологический материал этикетировали и хранили в пробирках либо в смонтированном виде. При сборе, выведении, хранении насекомых учитывали требования основной методической литературы (Тряпицын, Шапиро, Щепетильникова, 1982; Кирияк, 1984).
Оценку влияния на злаковых тлей и их энтомофагов элементов агрокомплекса проводили в 3 стационарных многофакторных опытах. В 1992-1993 гг. изучали влияние системы основной обработки почвы, средств химизации, размещения яровой пшеницы в севообороте. Численности тлей и их энтомофагов учитывали на 1-й и 4-й культурах после пара в пятипольном зернопаровом севообороте (пар – пшеница – пшеница – овес – пшеница). Основные обработки почвы – вспашка (на глубину 20-22см, на 3-4 культурах после пара – 25-27см), глубокое рыхление (на глубину 20-22см, на 3-4 культурах после пара – 25-27см), мелкая плоскорезная (минимальная) на глубину 12-14 см. Учеты проводили на трех уровнях химизации – без применения удобрений и пестицидов, с применением удобрений (фосфор в дозе 120 кг д.в./га в пару на ротацию севооборота, азот в дозе 60 кг д.в./га – под вторую и третью культуру, азот в дозе 80 кг д.в./га – под четвертую культуру), с внесением полного комплекса средств химизации (удобрения – по аналогичной схеме, пестициды – в соответствии с фитосанитарной ситуацией, складывающейся в агроценозе). В 1992 г. пшеницу в фазе 2-3 настоящих листьев обрабатывали Пума-супер 7.5, ЭМВ (1 л/га), в фазу кущения – 2,4-ДА, 40% ВК (2 л/га) + Це Це Це 460, ВК (2л/га), в начале фазы колошения – Тилтом, КЭ (0,5 л/га), в 1993 г. – в фазу кущения – Пума-супер 7.5, ЭМВ (1 л/га) + Гранстар, СТС (25 г/га) + Це Це Це 460, ВК (2 л/га), в фазу флагового листа – начала колошения – баковой смесью Байлетона, СП (1кг/га) и Фастака, КЭ (0,1 л/га). Площадь делянок 130 м, повторность опыта 3-х кратная.
В 1998-2000 гг. изучали влияние на злаковых тлей и их энтомофагов предшественников пшеницы. Учеты численности насекомых проводили на пшенице по пару, озимой ржи, второй пшенице после пара в двух зернопаровых севооборотах (пар черный – пшеница – пшеница – ячмень; пар черный – озимая рожь – пшеница – ячмень), на пшенице после овса, вико-овса, рапса в трех зерновых севооборотах (вико-овес – пшеница – пшеница – ячмень; рапс – пшеница – пшеница – ячмень; пшеница – овес – пшеница – ячмень), на бессменной пшенице и люцерне. Численность тлей, хищных и паразитических насекомых изучали на самих предшествующих культурах, а также на ячмене и ячмене с донником в зернотравяном севообороте (донник – озимая рожь – пшеница – ячмень). Поля севооборотов размером 475 м размещаются рендомизированно по блокам в 6 ярусов в 3-х кратной повторности.
В третьем опыте в 2005-2007 гг. оценивали влияние на злаковых тлей, хищных и паразитических насекомых технологий возделывания яровой пшеницы (сорт – Новосибирская 29). Экстенсивная технология (контроль) не предусматривала применение агрохимических средств. При нормальной технологии в дополнение к мероприятиям экстенсивного земледелия для снятия первого лимитирующего фактора урожайности (пищевого режима) вносили фосфор в дозе 20 кг д.в./га в рядки при посеве, а также гербициды против двудольных сорняков (2006 г. – Диален Супер, ВР, 0,6 л/га, 2005, 2007 гг. – Элант, КЭ, 0,7 л/га). При интенсивной технологии в дополнение к нормальной применяли протравливание семян (Раксил, КС, 1,5 л/т), азотные удобрения на планируемую урожайность по почвенной диагностике перед посевом, гербицид против однодольных сорняков (Пума супер 100, КЭ, 0,5 л/га), фунгицид (2005-2006 гг. – Фалькон, КЭ, 0,6 л/га, 2007 г. – Тилт, КЭ, 0,5 л/га), регулятор роста (2007 г. – Це ЦЕ Це 460, ВК, 1,5 л/га), при необходимости – инсектицид (2006 г. – Децис, КЭ, 0,25 л/га, 2007 г. – Децис Экстра, КЭ, 0,05л/га). Площадь делянок 2070 м (9Х230 м), расположение вариантов опыта систематическое, повторность опыта 3-х кратная.
Для учета вредной и полезной энтомофауны в течение вегетации с периодичностью в 7-10 дней делали кошения стандартным энтомологическим сачком, по 10-30 взмахов, в зависимости от размера делянок, на всех вариантах в трех-четырехкратной повторности. В дальнейшем для сравнимости данные пересчитывали на 10 и 50 взмахов.
Статистическую обработку полученных результатов проводили с помощью корреляционного, однофакторного и многофакторного дисперсионных анализов (Сорокин, 2004).
Период проведения исследований охватил все многообразие гидротермических условий. По увлажнению 1987, 1990, 1992, 1995, 1998, 2002, 2004 и 2007 гг. можно охарактеризовать как умеренно дефицитные; 1988, 1991, 1994, 1997 и 2005 гг. – дефицитные; 1989, 1999 2003 гг. – остродефицитные; 1986, 1993, 1996, 2000, 2001, 2006 и 2007 гг. – умеренно увлажненные.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
3. Видовой состав хищных и паразитических насекомых яруса
травостоя, трофически связанных со злаковыми тлями
С начала 90-х годов комплекс энтомофагов, трофически связанных со злаковыми тлями, не претерпел существенных изменений и к настоящему времени насчитывает 134 вида (рис. 1). Из них 89 видов (66,4%) – энтомофаги злаковых тлей, 45 (33,6%) – паразиты этих энтомофагов. Из 89 видов энтомофагов злаковых тлей 75 (84,3%) являются хищниками, 14 (15,7%) – паразитами. Из 45 видов паразитов энтомофагов злаковых тлей 27 (60,0%) – паразиты хищников, 18 (40,0%) – сверхпаразиты или вторичные паразиты злаковых тлей.
Вновь отмеченными видами являются паразиты златоглазок Helorus anomalipes (Panzer) и Baryscapus principiae Damenichini. B. principiae впервые зарегистрирован в фауне Западной Сибири.
Рис. 1. Структура комплекса энтомофагов, трофически связанных со злаковыми тлями
Развитие 76 видов (48 энтомофагов злаковых тлей и 28 паразитов энтомофагов) отмечено в травостое зерновых, 19 видов (12 энтомофагов злаковых тлей и 7 паразитов энтомофагов) – на черемухе, 39 видов (29 энтомофагов и 10 паразитов энтомофагов) развиваются как на травянистой, так и древесно-кустарниковой растительности, причем 27 из них (17 энтомофагов и 10 паразитов энтомофагов) встречаются в данных местообитаниях одинаково часто, 3 энтомофага предпочитают для своего развития зерновые, 9 (энтомофаги) – черемуху.
Все виды, являющиеся обычными для агроценозов зерновых культур, встречаются во всех обследуемых нами агроландшафтных районах Новосибирской области: в центрально-лесостепном Приобском, центрально-лесостепном Верхнекарасукском и южно-лесостепном Барабинском. Различия наблюдаются только в количественном соотношении видов, которое меняется во времени и пространстве.
4. Видовое соотношение, биоэкологические особенности и
сезонная динамика численности основных видов хищных энтомофагов злаковых тлей в агроценозах зерновых культур
В главе приводятся особенности фенологии, биоэкологии, сезонной и многолетней динамике численности видов сем. Nabidae, Anthocoridae, Coccinellidae, Chrysopidae, Hemerobiidae, Raphidiidae, Syrphidae, Chamaemyiidae. Наиболее многочисленными хищниками являются набиды, кокцинеллиды, сирфиды, хризопиды и антокориды.
Выявлено 6 видов клопов семейства Nabidae, трофически связанных со злаковыми тлями. Доминирующий вид – Nabis ferus L. Средняя суммарная численность его в течение вегетации составляет в травостое озимой ржи 41,2-92,9% общего обилия набисов, в травостое яровой пшеницы – 40,7-95,0%, овса – 51,9-96,1%.
Максимальная численность личинок наблюдается на ржи в третьей декаде июля, на яровых культурах – в конце первой – начале третьей декады августа и достигает в среднем, на ржи – 9-81, пшенице – 6-17,5, овсе – 2,5-9 особей/100 взмахов сачком. Количество взрослых набисов на посевах увеличивается после выхода нового поколения. Больше всего их встречается в агроценозе ржи в конце июля – второй декаде августа, пшеницы и овса – в третьей декаде августа. В этот период на 100 взмахов сачком на ржи приходится в среднем от 9,5 до 87,5 имаго клопов, на пшенице – от 7 до 29,5, на овсе – от 6 до 15 клопов.
Из 17 видов кокцинеллид доминирующими в агроценозах зерновых культур на протяжении 1987-1990 гг. были Coccinella septempunctata L., Propylaea quatuordecimpunctata L., Hippodamia tredecimpunctata L. Численность их составляла в травостое ржи, соответственно, 8,9-32,6, 17,2-24,3 и 14,0-66,0% суммарного количества собранных за вегетацию имаго кокцинеллид, пшеницы – 8,9-28,3, 28,9-43,4, 30,0-35,2%, овса – 11,3-34,8, 29,8-30,7, 33,3-55,7%. После 1990 г. на посевах озимой ржи по обилию стала преобладать Adonia variegatа Goeze (43,4-53,3%), пшеницы и овса – P. quatuorcimpunctata (31,3-76,7%). Кроме этих двух видов в агроценозах зерновых обильными в эти годы были также H. tredecimpunctata и A. amoena.
Максимум численности личинок коровок наблюдается во второй – третьей декаде июля. В это время на 100 взмахов сачком приходится в среднем от 2,5 (при низкой плотности популяций злаковых тлей) до 100-105 особей/100 взмахов сачком. Наибольшее количество жуков нового поколения встречается в первой – начале третьей декады августа и составляет в травостое озимой ржи в среднем 14-25, пшеницы – 48-69,5, овса – 30-85 особей на 100 взмахов.
Из 12 видов сем. Chrysopidae по обилию преобладают Chrysopa carnea Steph. и Ch. phyllochroma Wesm. Максимум численности личинок златоглазок наблюдается в первой – третьей декадах августа, в это время на 100 взмахов сачком приходится от 1,5 до 51 особей. Имаго нового поколения выходят с третьей декады июля – начала августа. Массовый выход – в конце августа – начале сентября.
Выявлено 29 видов сирфид, из них у 19 видов в колониях злаковых тлей найдены личинки, у 10 зафиксированы имаго. Доминирующими видами являются Episyrphus balteatus Deg., Sphaerophoria scripta L., Scaeva pyrastri L., Melanostoma mellinum L. и мухи рода Platycheirus Lep. and Serv.
Пик численности личинок сирфид в травостое ржи приходится на первую – начало второй декады июля, яровых культур – третью декаду июля – середину августа, в это время насчитывается до 20-40, редко – до 100 особей /100 взмахов сачком.
Из сем. Anthocoridae наиболее массовыми являются виды рода Orius Wolff., из них Оrius niger Wolff., составляющий от 40,2 до 100% общей численности ориусов. Максимальное количество личинок клопов (3-20 экз./100 взмахов) приходится на третью декаду июля – первую декаду августа. Обилие взрослых ориусов на озимой ржи увеличивается к началу – середине августа, на яровой пшенице и овсе – к началу августа – сентябрю. В это время численность их достигает 4-45 особей на 100 взмахов сачком.
Трофически наиболее сильно связаны со злаковыми тлями личинки сирфид, являющиеся афидофагами, имаго и личинки кокцинеллид, златоглазок (кроме имаго златоглазок, являющихся палинофагами), у которых, несмотря на возможность питания различными членистоногими, тли являются предпочитаемым видом корма (Савойская, 1983; Дорохова, 1987). У клопов набисов и ориусов злаковые тли составляют значительную, но часто не главную, часть рациона их питания (Пучков, 1961, 1976; Кержнер, 1965; Сливкин, 1985).
5. Видовое соотношение, зараженность паразитическими
перепончатокрылыми злаковых тлей
Кроме хищных энтомофагов важную роль в снижении плотности популяций злаковых тлей играют паразиты. Отмечено 14 видов афидиид и афелинид, из которых 2 вида паразитируют на большой злаковой тле, 6 – на черемухово-злаковой тле, 6 – заражают оба вида тлей (рис. 2). По заражению черемухово-злаковой тли на черемухе доминирует Praon volucre Hal., численность его составляет 36,4-83,9% суммарного количества выведенных из тлей паразитов. В отдельные годы значительно повышается обилие Ephedrus persicae Frogatt. (до 50,6%) и Aphelinus chaonia Walker (до 39,5%). В агроценозах зерновых культур преобладает Aphidius rhopalosiphi De Stefani, составляющий на озимой ржи 50,0-76,4%, на пшенице – 7,4-87,8%, на овсе – 56,4-76,8% численности паразитов. Значительную роль в сложении сообщества паразитических наездников на зерновых культурах занимает P. volucre (до 45,3%), на пшенице в отдельные годы доминирует A. transversus Thomson (до 91,2%).

Рис. 2. Трофические связи паразитов злаковых тлей
Большинство перепончатокрылых паразитов широкие олигофаги, паразитируют на тлях от 2 до 37 родов (Тобиас, Кирияк, 1986; Elliott et аl., 1994). Значительно более узкий круг хозяев имеют Parapraon necans, заражающий Rhopalosiphum insertum Walk. и Rh. nymphaeae L. (Тобиас, Кирияк, 1986), Aphelinus transversus, зарегистрированный ранее лишь на Hayhurstia atriplicis L. (Курдюмов, 1913 б; Яснош, 1964). Паразиты предпочитают заражать личинок и бескрылых самок тлей.
В агроценозах зерновых в наибольшем количестве мумифицированные тли встречаются в период максимума плотности популяций вредителей (до 50 шт./м на озимой ржи, до 70 шт./м – на пшенице, до 130 – на овсе). Однако число их в это время не превышает 4,6% общего обилия тлей. В большем количестве по отношению к численности тлей мумии отмечены на озимой ржи в начале размножения вредителей на полях (5,9-8,3%) и через 2 недели после пика их численности (8,8-14,6%). На яровых культурах при резком снижении плотности популяций вредителей, когда на полях остаются практически одни мумии, число их достигает 91-100%, при более медленном снижении плотности популяциий вредителей – 22-33% общей численности тлей. При незначительной заселенности зерновых культур злаковыми тлями зараженные паразитами особи вредителей встречаются единично. Необходимо отметить, что количество мумий в травостое зерновых не отражает в полной мере степень паразитизма вредителей наездниками, так как часть тлей уничтожается хищниками, часть – покидает посевы, не успев мумифицироваться.
Осенью, при поздней миграции большой злаковой тли на всходы озимой ржи и невысокой ее численности, паразиты заражают небольшую часть вредителей (до 14%). В противоположном случае наездники начинают заражать тлю раньше, к наступлению холодов незараженными остаются 13,5-18% вредителей. Из паразитов, выведенных из мумий тлей, собранных осенью на озимой ржи, 41,7-100% приходится на А. rhopalosiphi, до 58,3% – на P. voluсre.
6. Видовое соотношение и роль паразитов в снижении численности
энтомофагов злаковых тлей
Численность личинок и куколок кокцинеллид снижают 3 вида паразитов (Phalacrotophora fasciata Fall., Oomyzus scaposus Thomson, Homalotylus flaminius Dalm.), личинок сирфид – 18 (доминируют Promethes sulcator Grav., Diplazon laetatorius F., Syrphoctonus signatus Grav., Woldstedtius biguttatus Grav., Pachyneuron groenlandicum Holmgr.), мух-серебрянок – 1 (Melanips opacus Hartig). Из коконов златоглазок выведено 3 вида паразитов (Baryscapus principiae Damenichini, Helorus anomalipes (Panzer), Isodromus vinulus Dalm.). На афидиидах и афелинидах паразитируют 20 видов (по заражению афидиид доминируют Dendrocerus carpenteri Curtis, Asaphes suspensus Nees., Phaenoglyphis villosa Hartig, Alloxysta sp. 1, афелинид – Alloxysta mullensis Cameron). В целом на посевах зерновых культур паразиты снижают численность кокцинеллид на 8,6-55,3 %, личинок сирфид – на 3,2-57,1%, афидиид и афелинид – на 30,6-85,6%; на черемухе паразиты заражают до 40% сирфид, 17,3-88,8% – афидиид и афелинид. В годы с низкой численностью энтомофагов паразитирования на них наездников не происходит либо бывает очень слабым.
7. Эффективность хищных и паразитических энтомофагов
в ограничении численности злаковых тлей
Отдельные виды афидофагов не могут в одиночку ограничить плотность популяции злаковых тлей до принятого уровня экономического порога вредоносности. В биоценотическом комплексе афидофаги действуют по принципу дополнительности, когда энтомофагия одного вида усиливается действием других. В итоге на динамику численности тлей оказывается мощный биоценотический пресс, позволяющий при определенном соотношении афидофаг:тля регламентировать инсектицидные обработки (Воронин, Воронина, 1996; Воронин и др., 2000).
Выявленный нами в лесостепи Западной Сибири комплекс энтомофагов злаковых тлей динамичен. В течение сезона и по годам наблюдаются колебания численности хищников и паразитических перепончатокрылых, обусловленные их видовыми особенностями, погодными условиями, зараженностью энтомофагов паразитами, для некоторых перепончатокрылых – видовым составом тлей и т.д. Существенное влияние на динамику численности полезных видов оказывает плотность популяций злаковых тлей. Установлена тесная прямая связь между суммарной численностью тлей и энтомофагов за период вегетации (в агроценозе озимой ржи: r = +0,88, яровой пшеницы и овса: r = +0,80), а также во время максимальной плотности популяций вредителей (на озимой ржи: r = +0,72, на яровых культурах: r = +0,86).
В лесостепи Западной Сибири наибольшее значение в снижении численности тлей играют хищники, а из них – кокцинеллиды (рис. 3, 4). Сдерживание кокцинеллидами численности тлей начинается весной на черемухе. На яровых культурах они появляются одними из первых и остаются в преобладающем количестве практически в течение всей вегетации, составляя на пшенице и овсе, соответственно, от 32,6 до 100 и от 40,0 до 100% обилия хищных энтомофагов.

а

б
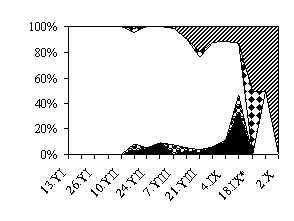
Рис. 3. Сезонная динамика соотношения энтомофагов в агроценозе
яровой пшеницы, %
а – 1987 г.; б – 1988 г. * – зерновые на полях убраны
Пик численности личинок кокцинеллид совпадает с пиком численности тлей либо отклоняется от него в среднем на неделю в ту или другую сторону. Во время максимальной плотности популяций вредителей в агроценозах пшеницы и овса кокцинеллиды составляют, соответственно, 28,6-70 и 33-90,9% суммы хищных энтомофагов (рис. 5). За сезон на долю кокцинеллид на первой культуре приходится 28,2-64,2%, на второй – 51,9-82,3% общего количества хищников (рис. 6). Исключением являются 1998 и 1999 гг., когда кокцинеллиды были малочисленны. В результате наших исследований в агроценозах яровых зерновых выявлена тесная прямая связь суммарной численности злаковых тлей и хищных кокцинеллид за период вегетации (r = +0,89), а также численности злаковых тлей и кокцинеллид во время максимальной плотности популяций вредителей (r = +0,93).
а

б
Рис. 4. Сезонная динамика соотношения энтомофагов в агроценозе овса, %
а – 1986 г.; б – 1987 г.; * – зерновые на полях убраны
На озимой ржи кокцинеллиды преобладают до середины июля. Численность их в это время составляет 47,6-82,4% суммарного количества хищников, в дальнейшем снижается до 1-20% (рис. 7). Поэтому связь между обилием злаковых тлей и хищных кокцинеллид в агроценозе ржи выражена слабее (r = +0,59). Во время пика численности тлей на долю кокцинеллид приходится 12-68%, за сезон – 11,2-69,8% суммы хищных энтомофагов (см. рис. 5, 6). Также как на яровых хлебах минимальное количество кокцинеллид на озимой ржи отмечено в 1998 и 1999 гг.
Со второй половины лета в травостое ржи преобладают клопы рода Nаbis. Значительную роль клопов в агроценозе озимой ржи подтверждает тесная связь между их количеством и численностью злаковых тлей как за период вегетации, так и во время максимума плотности популяций вредителей (r = +0,88). На яровых зерновых основная роль набисов часто сводится не к предотвращению нарастания, а к ускорению спада численности тлей. Отсутствие связи между обилием злаковых тлей и хищных набисов за период вегетации в агроценозах яровой пшеницы и овса подтверждает это (r = +0,35).
а


Рис. 5. Соотношение хищных энтомофагов во время пика численности
злаковых тлей, %
а – озимая рожь; б – пшеница; в – овес
Роль личинок сирфид и видов сем. Chrysopidae в комплексе энтомофагов злаковых тлей увеличивается в годы с высокой плотностью их популяций в агроценозах. Клопы ориусы, вследствие невысокой численности, небольшой массы тела, а отсюда и потребности в пище, по эффективности значительно уступают другим энтомофагам. Положительным их свойством является возможность уничтожать тлей за колосковыми чешуйками, в пазухах листьев и других местах, труднодоступных для остальных хищников.


Рис. 6. Соотношение хищных энтомофагов злаковых тлей в агроценозах зерновых культур в целом за вегетацию, %
а – озимая рожь; б – пшеница; в – овес
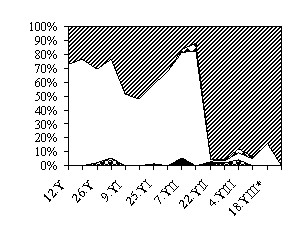

а
б
Рис. 7. Сезонная динамика соотношения энтомофагов в агроценозе
озимой ржи, %
а – 1987 г.; б – 1988 г. * – зерновые на полях убраны
Об эффективности энтомофагов говорит сдерживание ими темпов размножения тлей, снижение пика численности вредителей, укорочение периода его спада.
В естественных условиях полную ликвидацию вредителя-хозяина не способен обеспечить ни один из наиболее эффективных энтомофагов вследствие неполной приуроченности энтомофагов к хозяевам, меньшей их экологической пластичностью по сравнению с последними. В свою очередь сохранение некоторой части вредителей позволяет паразитам и хищникам выжить (Щепетильникова, 1954, 1957).
Одним из показателей эффективности хищных энтомофагов является их прожорливость. Приводятся сведения по прожорливости личинок сирфид, имаго и личинок 5 видов кокцинеллид, 2 златоглазок, хищных набисов и ориусов.
Наибольшее число тлей приходится на одного хищника во время максимальной плотности популяций вредителей. На яровых зерновых увеличение соотношения хищник:жертва может наблюдаться также в начале вегетационного периода, когда численность энтомофагов еще не высока, однако по мере их миграции на посевы происходит снижение этого показателя. В агроценозе озимой ржи вспышки размножения тлей нами не отмечены, соотношение хищник:жертва в большинстве изучаемых лет не превышает 1:30. Наибольшим оно бывает во время максимальной плотности популяций тлей, а также за неделю и через неделю после ее наступления (рис. 8).
а
б
в
Рис. 8. Соотношение хищник:жертва во время пика
численности тлей в агроценозах зерновых культур
а – озимая рожь; б – яровая пшеница; в – овес
В агроценозах яровых культур соотношение хищник:жертва во время максимальной численности тлей держится на уровне до 1:40, лишь в отдельные годы увеличивается до 1:55. В эти годы численность тлей превышает ЭПВ. Однако и при таком соотношении энтомофаги в течение недели практически полностью очищают посевы от тлей.
Таким образом, по уточненным нами данным, при соотношении хищник:жертва на озимой ржи 1:30, на пшенице и овсе 1:40 численность злаковых тлей не поднимается до порога экономической вредоносности. В связи с этим указанные соотношения рекомендуется принять за критерий эффективности энтомофагов в условиях лесостепи Западной Сибири.
8. Роль лесополос и окружающей растительности в повышении
численности хищных и паразитических энтомофагов
На основе изучения видового состава, фенологии, сезонной динамики численности наиболее массовых видов хищных и паразитических энтомофагов в лесополосах, прилегающих к посевам зерновых культур, на многолетних и двулетних травах: люцерне, доннике, клевере, эспарцете, костреце безостом, полянах дикорастущего разнотравья сделан вывод о том, что изучаемые стации являются не только местом зимовки энтомофагов, но и постоянным местом их обитания и размножения. В лесополосах наиболее часто встречаются кокцинеллиды C. septempunctata, H. tredecimpunctata, P. quatuordecimpunctata, златоглазки Ch. carnea и Ch. perla. В колониях тлей на бодяке щетинистом и борщевике сибирском, произрастающих в лесополосах, наблюдается развитие данных видов от яйца до имаго. Постоянными обитателями лесополос являются клопы набисы, причем некоторые виды (N. limbatus) более распространены в лесополосах, чем в травостое зерновых. В прилегающих к зерновым лесополосах отмечается размножение клопов ориусов. Наиболее интенсивно оно происходит в годы, когда плотность популяций тлей в агроценозах зерновых невысока. В течение всей вегетации на цветущей растительности по краям лесополос питаются имаго сирфид, паразитические перепончатокрылые.
Установлено, что из многолетних и двулетних трав предпочитаемым местом обитания и размножения кокцинеллид и златоглазок являются люцерна, донник, клевер, на костреце личинки кокцинеллид не найдены, личинки златоглазок – единичны. Клопы рода Orius обитают на всех изучаемых многолетних и двулетних травах, единично встречаются на костреце и дикорастущем разнотравье. Для Nabis ferus, N. punctatus предпочитаемым местообитанием являются кострец, люцерна, клевер, для N. flavomarginatus – кострец и дикорастущее разнотравье, N. brevis – дикорастущее разнотравье. В течение всей вегетации цветущую растительность посещают сирфиды. Однако размножение сирфид на многолетних травах и дикорастущем разнотравье либо не происходит, либо бывает незначительным. Возможно микроклимат, создающийся в загущенных посевах травянистой растительности для них неблагоприятен.
Многолетние травы являются резерватом многих видов перепончатокрылых наездников. Особую роль травянистой растительности в динамике численности паразитических перепончатокрылых подчеркивал P. Stary (1978). Нами отмечено, что наименее интенсивно лёт имаго паразитов родов Aphidius и Praon происходит на клевере, костреце, рода Aphelinus – на клевере и доннике. Больше всего паразитов зарегистрировано на люцерне, смеси многолетних трав с сорной растительностью, дикорастущем разнотравье.
Численность хищников и имаго паразитов на многолетних травах превышает количество энтомофагов, встречающихся в травостое пшеницы. Так, в среднем за 1998-2000 гг. кокцинеллид на люцерне было больше, чем на посевах пшеницы в 5,1 раз, златоглазок – в 2,6, набисов – в 2,8, имаго первичных паразитов – в 2,1 раза (табл. 1). Люцерна является местом массового размножения клопов ориусов. Обилие их имаго и личинок на люцерне в среднем за три года в 72,7 раз превышало количество этих насекомых на пшенице. Исключение составляют сирфиды, более интенсивно размножающиеся в травостое зерновых. Суммарная численность хищников на люцерне за сезон была в 15,3 раза больше количества энтомофагов в агроценозе пшеницы.
Таблица 1 – Суммарная численность энтомофагов в течение вегетации в
травостое люцерны и яровой пшеницы, экз./100 взмахов сачком
| Энтомофаги | Люцерна | Пшеница | ||||||
| 1998 | 1999 | 2000 | В среднем за три года | 1998 | 1999 | 2000 | В среднем за три года | |
| Кокцинеллиды | 353 | 477 | 190 | 340 | 43 | 27 | 130 | 67 |
| Сетчатокрылые | 287 | 350 | 60 | 232 | 110 | 127 | 30 | 89 |
| Личинки сирфид | 10 | 13 | 13 | 12 | 47 | 13 | 23 | 28 |
| Клопы набисы | 220 | 147 | 57 | 141 | 93 | 27 | 30 | 50 |
| Клопы ориусы | 4903 | 4880 | 1117 | 3633 | 37 | 97 | 17 | 50 |
| Суммарная численность | 5773 | 5867 | 1437 | 4358 | 330 | 291 | 230 | 284 |
| Первичные паразиты тлей (имаго) | 110 | 140 | 24 | 91 | 43 | 17 | 70 | 43 |
Лесополосы, многолетние травы и поляны дикорастущего разнотравья в течение вегетационного периода способствуют повышению численности энтомофагов за счет предоставления им возможности дополнительного питания нектаром, пыльцой цветущей травянистой растительности, развития полифагов на промежуточных хозяевах. Кроме того, энтомофаги находят здесь убежище во время обработок инсектицидами близлежащих полей. Зная доминирующие и наиболее эффективные виды энтомофагов на тех или иных культурах, и размещая рядом с посевами этих культур предпочитаемые энтомофагами многолетние травы, можно полосными укосами трав повышать численность хищников и паразитических перепончатокрылых на защищаемой культуре.
9. Влияние предшественников на численность злаковых тлей
и их энтомофагов
Найдено, что в неблагоприятных условиях для развития злаковых тлей (1999 г. – сухая жаркая погода, обработка инсектицидом, проведенная в фазу кущения пшеницы) на бессменной пшенице питается минимальное их количество (в среднем 15 особей/10 взмахов во время пика численности, 37 – за период вегетации), сравнима с ней численность вредителей на пшенице после ячменя, рапса, вико-овса. На пшенице по лучшим предшественникам – пару, пшенице после пара, озимой ржи, а также овсу плотность популяции тлей достоверно выше (в 6,6-9,3 раза), чем на бессменных посевах (степень влияния фактора 44,4%). При благоприятных условиях для развития злаковых тлей (1998, 2000 гг.), также как и в среднем за три года, достоверных различий в численности вредителей на бессменной пшенице и на пшенице по различным предшественникам не зарегистрировано (степень влияния фактора 0,0-4,3%) (табл. 2).
Таблица 2 – Влияние предшественников на численность злаковых тлей и их энтомофагов в агроценозе яровой пшеницы, экз./10 взмахов сачком
(средние за 1998-2000 гг.)
| Предшественник | Злаковые тли | Coccinellidae | Syrphidae | Nabidae | Chrysopidae | Род Orius | Паразиты тлей (имаго) | Сумма хищных энтомофагов |
| Пар | 310 | 6,7 | 2,8 | 5,0 | 8,9 | 5,0 | 4,3 | 28,4 |
| Пшеница после пара | 335 | 4,2 | 3,5 | 1,0 | 11,2 | 2,7 | 2,2 | 22,6 |
| Озимая рожь | 369 | 4,2 | 3,5 | 1,8 | 10,7 | 3,0 | 1,8 | 23,2 |
| Овес | 309 | 3,1 | 4,1 | 2,9 | 9,6 | 4,1 | 2,3 | 23,8 |
| Вико-овес | 282 | 4,0 | 3,5 | 2,9 | 5,9 | 3,9 | 2,0 | 20,2 |
| Рапс | 280 | 2,2 | 2,8 | 2,7 | 7,6 | 3,1 | 2,3 | 18,4 |
| Ячмень | 331 | 3,0 | 2,2 | 3,1 | 9,0 | 4,2 | 2,2 | 21,5 |
| Бессменная пшеница | 251 | 5,7 | 2,1 | 3,4 | 8,9 | 13,1 | 2,7 | 34,2 |
| НСР05 | 119 | 4,3 | 3,7 | 2,8 | 7,2 | 5,2 | 2,6 | 11,8 |
Влияние предшественников на обилие в течение вегетации в агроценозе яровой пшеницы кокцинеллид, златоглазок, сирфид отсутствует, либо не превышает среднего уровня. Отмечено явное предпочтение ориусами посевов бессменной пшеницы, численность их здесь ежегодно в 1,7-5,4 раза выше, чем на посевах пшеницы в севооборотах. В отдельные годы достоверно больше набисов и имаго паразитических наездников находили на пшенице по пару, хотя в среднем за три года это различие сглаживается.
Высокий уровень суммарной численности хищных энтомофагов яруса травостоя ежегодно поддерживается на посевах пшеницы после пара и на бессменной пшенице, соответственно, на уровне 23,0-33,0 и 25,0-47,7 особей на 10 взмахов сачком за вегетацию (в среднем за три года 28,4 и 34,2), степень влияния фактора 23,8-48,4%. Концентрации хищников и паразитических перепончатокрылых здесь способствует высокая численность злаковых тлей, на бессменной пшенице, по-видимому, также более благоприятные микроклиматические условия для некоторых видов энтомофагов и лучшие условия для сохранения их популяций в послеуборочный период в связи с близостью мест зимовки.
В зерновых, зернопаровых и зернотравяном севооборотах на самих предшествующих культурах, таких как озимая рожь, ячмень, ячмень с донником, рапс средняя суммарная численность тлей за вегетационный период ниже, соответственно, в 3,3, 3,4, 2,3, 5,3 раза по сравнению с пшеницей по пару (табл. 3). Паровой предшественник создает благоприятные условия для роста и развития растений пшеницы, улучшения их пищевой ценности, чем способствует привлечению на посевы злаковых тлей. На овсе и вико-овсе обилие злаковых тлей сравнимо с их численностью на пшенице по пару или достоверно превышает ее. Вследствие предпочтения заселения тлями тех или иных культур степень влияния последних на суммарное количество в их агроценозах вредителей в течение вегетации сильная и варьирует от 55,7 до 77,9%.
Таблица 3 – Численность энтомофагов на различных культурах в севооборотах, экз./10 взмахов сачком (средние за 1998-2000 гг.)
| Культуры | Злаковые тли | Coccinellidae | Syrphidae | Nabidae | Chrysopidae | Род Orius | Паразиты тлей (имаго) | Сумма хищных энтомофагов |
| Пшеница по пару | 310 | 6,7 | 2,8 | 5,0 | 8,9 | 5,0 | 4,3 | 28,4 |
| Озимая рожь по пару | 93 | 2,9 | 2,2 | 6,3 | 4,4 | 2,1 | 1,4 | 18,9 |
| Овес | 577 | 8,8 | 5,3 | 4,2 | 7,2 | 1,1 | 4,8 | 27,7 |
| Вико-овес | 430 | 7,0 | 4,0 | 3,2 | 4,8 | 10,8 | 7,8 | 29,8 |
| Рапс** | 58 | 2,0 | 1,3 | 2,8 | 3,8 | 5,9 | 1,2 | 15,8 |
| Ячмень | 92 | 4,5 | 2,3 | 1,8 | 1,8 | 0,6 | 1,4 | 11,0 |
| Ячмень+ донник | 132 | 5,2 | 2,5 | 5,0 | 2,1 | 0,1 | 4,3 | 14,9 |
| Люцерна* | - | 34,0 | 1,2 | 14,1 | 23,2 | 363,3 | 16,3 | 435,8 |
| НСР05 | 179 | 5,1 | 3,5 | 4,5 | 4,0 | 4,7 | 5,1 | 11,0 |
* при расчете НСР данные по люцерне не включались
** вид тлей не определялся
Численность энтомофагов за вегетацию также зависит от культуры, на которой они развиваются (степень влияния фактора 48,0-77,3%). На отдельные группы энтомофагов, при низкой плотности их популяций, культуры не оказывают влияния, при значительной – влияние от среднего до высокого (на кокцинеллид – до 43,6%, златоглазок – до 66,8, сирфид – до 46,4, ориусов – до 78,8, набисов – до 19,0, имаго первичных паразитов – до 66,8%). Достоверно больше златоглазок встречается на пшенице по пару, овсе, имаго перепончатокрылых паразитов, кроме этого на вико-овсе, ориусов – вико-овсе, у сирфид и кокцинеллид существует тенденция большего заселения этих культур. Набисы отмечены на посевах различных культур в одинаковой численности. Ежегодно обилием всех хищных энтомофагов выделяются пшеница после пара, овес и вико-овес, то есть посевы культур, наиболее заселяемые злаковыми тлями. Количество их здесь за вегетационный период составляет в среднем за три года, соответственно, 28,4, 27,7 и 29,8 особей на 10 взмахов сачком или в 1,6-2,7 раза больше, чем на других культурах.
Предпочитаемым местом питания и размножения хищных набисов, кокцинеллид, златоглазок, клопов ориусов, имаго афидиид и афелинид являются посевы бессменной люцерны. Суммарное количество имаго и личинок хищников на люцерне в 3,8-156 раз превышает их обилие в травостое пшеницы, озимой ржи, ячменя, овса, вико-овса, рапса.
10. Влияние системы основной обработки почвы, средств
химизации, размещения яровой пшеницы в севообороте на
злаковых тлей и их энтомофагов
Нами установлено, что системы основной обработки почвы как на 1-й, так и 4-й пшенице после пара существенно не сказываются на численности злаковых тлей, хищных и паразитических энтомофагов (степень влияния фактора 2,1- 5,9%).
Из основных элементов агрокомплекса возделывания яровой пшеницы наибольшее влияние на злаковых тлей и их энтомофагов оказывают внесение минеральных удобрений и пестицидов, место культуры в севообороте (табл. 4, 5). Так, на пшенице, выращиваемой с внесением сбалансированного минерального питания и пестицидов, численность тлей в среднем по опыту составляет 281 особь/10 взмахов, что в 1,9-3,5 раза превышает их количество на пшенице, возделываемой с внесением одних удобрений и в 1,7-5,9 раза – без внесения средств химизации (степень влияния фактора 65-83%).
Применение одних удобрений без средств защиты растений не влияет на рост плотности популяций тлей, либо обилие их увеличивается, но не в такой степени, как от полного комплекса средств химизации. Средняя численность злаковых тлей на пшенице по пару в 1,3 раза больше их количества на четвертой пшенице после пара (степень влияния фактора 3,2-24,5%).
Применение комплекса средств химизации наиболее сильно влияет на обилие видов сем. Coccinellidae (24,9-54,7%) и Syrphidae (64,9-70,6%), суммарную численность энтомофагов (41,7-49,9%), а также имаго перепончатокрылых паразитов (54,5-61,6%). Место культуры в севообороте – на обилие кокцинеллид (9,9-23,8%), клопов набисов (21,9-28,3%) и ориусов (25,1-39,1%), суммарную численность энтомофагов (18,2-19,8%). Влияние изучаемых факторов на обилие энтомофагов опосредуется их воздействием на численность злаковых тлей, реже – предпочтением определенного местообитания. Так, в среднем по опыту количество кокцинеллид на посевах с полным комплексом средств химизации составляет 16,1 особей/10 взмахов сачком, что в 2,2 раза превышает их обилие на посевах с внесением одних удобрений и в 1,8 раз – без внесения удобрений и пестицидов. На пшенице по паровому предшественнику средняя численность кокцинеллид за вегетацию равняется 13 особям/10 взмахов, что в 1,5 раз превышает их количество на четвертой пшенице после пара.
Таблица 4 – Влияние системы обработки почвы, применения средств
химизации и места культуры в севообороте на численность злаковых тлей и их энтомофагов, экз./10 взмахов сачком (средние за 1992-1993 гг.)
| Система обработки почвы | Уровень химизации | Злаковые тли | Coccinellidae | Syrphidae | Nabidae | ||||
| Пшеница после пара | |||||||||
| 1-я | 4-я | 1-я | 4-я | 1-я | 4-я | 1-я | 4-я | ||
| Вспашка | 1 | 94 | 100 | 7,8 | 7,9 | 1,7 | 2,7 | 3,1 | 1,5 |
| 2 | 115 | 105 | 9,1 | 8,2 | 3,8 | 2,5 | 1,2 | 0,8 | |
| 3 | 333 | 337 | 21,5 | 7,9 | 13,3 | 7,5 | 4,0 | 0,3 | |
| Глубокое рыхление | 1 | 78 | 107 | 8,5 | 7,9 | 1,3 | 1,5 | 1,7 | 0,9 |
| 2 | 129 | 77 | 7,6 | 6,5 | 2,2 | 2,1 | 0,8 | 0,6 | |
| 3 | 301 | 237 | 21,9 | 12,8 | 11,9 | 6,4 | 4,3 | 1,5 | |
| Плоскорезная обработка | 1 | 154 | 79 | 8,5 | 7,5 | 1,3 | 2,6 | 2,7 | 0,8 |
| 2 | 152 | 83 | 6,9 | 6,3 | 2,9 | 1,8 | 1,2 | 0,5 | |
| 3 | 339 | 229 | 18,5 | 8,3 | 11,3 | 6,7 | 3,0 | 1,1 | |
| Без обработки | 1 | 143 | 81 | 14,5 | 7,5 | 2,9 | 2,2 | 3,1 | 0,8 |
| 2 | 139 | 99 | 7,6 | 5,9 | 2,6 | 2,5 | 0,9 | 0,5 | |
| 3 | 300 | 174 | 23,4 | 14 | 11,1 | 6,5 | 4,5 | 0,9 | |
| В среднем по культурам | 190 | 142 | 13,0 | 8,4 | 5,5 | 3,8 | 2,5 | 0,9 | |
| Средние по фактору система обработки почвы | I II III IV | 181 155 173 156 | 10,4 10,9 9,3 12,1 | 5,2 4,3 4,5 4,7 | 2,6 1,3 1,5 1,7 | ||||
| Средние по фактору уровень химизации | 1 2 3 | 105 112 281 | 8,7 7,3 16,1 | 2,0 2,5 9,3 | 2,4 0,7 2,3 | ||||
| НСР0.5 по факторам: место культуры в севообороте 45,0 2,7 0,9 1,0 система обработки почвы 59,0 4,1 1,3 1,3 уровень химизации 47,0 2,6 1,1 1,2 частных средних 123,0 7,0 2,6 3,3 | |||||||||
Примечание: 1- без применения средств химизации, 2 – с внесением
удобрений, 3 – с внесением полного комплекса средств химизации
Представители семейства Syrphidae, являясь специализированными хищниками тлей, также повторяют динамику их численности и в большем количестве отмечены на посевах пшеницы с наиболее интенсивным развитием и размножением вредителей. Средняя численность личинок сирфид за период вегетации на пшенице, выращиваемой с полным комплексом средств химизации составляет 9,3 особей/10 взмахов, что в 3,7 раз превышает их обилие при возделывании пшеницы с внесением одних удобрений и в 4,7 раз – без удобрений и пестицидов. Количество сирфид в среднем за 2 года на пшенице по пару равняется 5,5 особям/10 взмахов, на четвертой пшенице после пара – 3,8, то есть в 1,4 раза меньше.
Таблица 5 – Влияние системы обработки почвы, уровня химизации и места культуры в севообороте на численность энтомофагов злаковых тлей,
экз./10 взмахов сачком (средние за 1992-1993 гг.)
| Система обработки почвы | Уровень химизации | Chrysopidae | Род Orius | Сумма хищных энтомофагов | Первичные паразиты тлей (имаго) | ||||
| Пшеница после пара | |||||||||
| 1-я | 4-я | 1-я | 4-я | 1-я | 4-я | 1-я | 4-я | ||
| Вспашка | 1 | 4,7 | 4,6 | 2,1 | 0,9 | 19,3 | 17,5 | 3,5 | 4,1 |
| 2 | 3,2 | 1,9 | 2,5 | 1,3 | 19,6 | 14,6 | 5,0 | 5,9 | |
| 3 | 3,7 | 2,9 | 1,2 | 1,5 | 43,7 | 19,9 | 12,1 | 14,0 | |
| Глубокое рыхление | 1 | 3,2 | 1,4 | 2,1 | 1,8 | 16,7 | 13,3 | 2,9 | 4,1 |
| 2 | 3,1 | 1,9 | 2,5 | 1,3 | 16,1 | 12,3 | 5,2 | 5,2 | |
| 3 | 3,2 | 3,3 | 2,0 | 0,6 | 41,9 | 24,3 | 10,9 | 9,6 | |
| Плоскорезная обработка | 1 | 4,5 | 3,6 | 2,1 | 1,1 | 19,2 | 15,7 | 3,8 | 4,8 |
| 2 | 3,2 | 2,7 | 3,8 | 1,1 | 17,9 | 12,2 | 5,7 | 6,9 | |
| 3 | 2,9 | 1,5 | 2,3 | 0,9 | 37,9 | 18,5 | 10,3 | 9,7 | |
| Без обработки | 1 | 3,6 | 2,9 | 2,5 | 1,5 | 26,5 | 14,8 | 5,1 | 5,6 |
| 2 | 4,0 | 3,1 | 3,8 | 0,9 | 18,8 | 12,9 | 5,2 | 8,1 | |
| 3 | 3,6 | 4,3 | 1,5 | 0,8 | 43,9 | 26,5 | 9,0 | 9,5 | |
| В среднем по культурам | 3,6 | 2,8 | 2,4 | 1,1 | 26,8 | 16,9 | 6,6 | 7,3 | |
| Средние по фактору система обработки почвы | I II III IV | 3,5 2,7 3,1 3,6 | 1,5 1,7 1,9 1,9 | 22,4 20,8 20,3 24,0 | 7,3 6,3 6,3 6,2 | ||||
| Средние по фактору уровень химизации | 1 2 3 | 3,5 2,9 3,2 | 1,8 2,1 1,3 | 17,9 15,5 32,3 | 4,1 5,6 9,8 | ||||
| НСР0.5 по факторам: место культуры в севообороте 1,1 0,5 4,5 1,9 система обработки почвы 1,1 0,7 5,1 2,5 уровень химизации 1,4 0,6 3,6 2,0 частных средних 2,8 1,5 12,6 4,9 | |||||||||
Примечание: 1- без применения средств химизации, 2 – с внесением
удобрений, 3 – с внесением полного комплекса средств химизации
Плотность популяций хищных клопов сем. Nabidae и рода Orius на пшенице не зависит от уровня химизации и достоверно изменяется в зависимости от места культуры в севообороте. Хищные клопы предпочитают заселять пшеницу по пару, отличающуюся более мощными и развитыми растениями, отсутствием сорняков и большей плотностью популяций вредителей. Средняя численность клопов набисов и ориусов за вегетацию на пшенице по паровому предшественнику составляет 2,4-2,5 особей/10 взмахов, что соответственно в 2,8 раз и 2,2 раза превышает их количество на четвертой пшенице после пара.
Четкой приуроченности видов сем. Chrysopidae к вариантам с большей численностью тлей или с тем или иным уровнем агротехники и местом культуры в севообороте не найдено.
Лет имаго первичных паразитов злаковых тлей происходит по всему массиву пшеницы. Достоверно чаще паразитические перепончатокрылые встречаются на посевах, выращиваемых с применением полного комплекса средств химизации. Средняя численность имаго паразитов за период вегетации составляет здесь 9,8 особей/10 взмахов, что в 1,8 раз превышает их количество на пшенице, выращиваемой с применением одних удобрений и в 2,4 раза – без пестицидов.
Количество хищных энтомофагов на пшенице, возделываемой с комплексным применением удобрений и пестицидов, составляет 32,3 особи/10 взмахов, что в 2,1 раз больше, чем на пшенице, выращиваемой с внесением одних удобрений и в 1,8 раз – без удобрений и пестицидов. Средняя суммарная численность хищников за вегетацию на четвертой пшенице после пара составляет 16,9 особей/10 взмахов, на первой пшенице после пара – 26,8 или в 1,6 раза больше.
Таким образом, в условиях лесостепи Западной Сибири средний уровень плотности популяций злаковых тлей и их энтомофагов не зависит от системы основной обработки почвы. Замена глубокой пахоты с оборотом пласта поверхностными обработками, в качестве приема, способствующего энерго-ресурсосбережению, не приводит к возрастанию угрозы со стороны вредных членистоногих, не изменяет общего характера трофической структуры сообщества пшеничного поля. Применение удобрений и пестицидов, а также выращивание пшеницы по паровому предшественнику создают благоприятные условия для роста и развития растений. Такие растения формируют большую вегетативную массу, более крупный и озерненный колос, более длительное время остаются зелеными и сочными, содержат, по данным Г.Ф. Дудник (1985), повышенное количество моносахаров, что способствует привлечению и нарастанию численности злаковых тлей. Рост плотности популяций тлей, в свою очередь, приводит к росту численности энтомофагов, эффективно снижающих плотность популяций вредителей. Особенно заметна синхронизация в развитии со злаковыми тлями у кокцинеллид, сирфид, перепончатокрылых паразитов. Экологическая стабильность агроценоза не нарушается.
11. Влияние технологии возделывания яровой пшеницы
на злаковых тлей и их энтомофагов
В результате проведенных нами исследований найдено, что численность злаковых тлей и хищных и паразитических энтомофагов на пшенице, возделываемой по экстенсивной и нормальной технологиям, различается недостоверно (табл. 6, 7).
Таблица 6 – Влияние технологии возделывания яровой пшеницы на
численность злаковых тлей, клопов pода Nabis и Orius (экз./50 взмахов сачком)
| Технология возделывания | Злаковые тли | Клопы р. Nabis | Клопы р. Orius | ||||||
| 2005 | 2006 | 2007 | 2005 | 2006 | 2007 | 2005 | 2006 | 2007 | |
| Предшественник – пар | |||||||||
| Экстенсивная | - | 357 | 410 | - | 7,2 | 2,2 | - | 16,2 | 16,7 |
| Нормальная | - | 472 | 402 | - | 7,8 | 2,8 | - | 13,8 | 25,0 |
| Интенсивная | - | 662 | 488 | - | 9,5 | 2,2 | - | 26,2 | 41,2 |
| НСР0.5 | - | 243 | 267 | - | 8,2 | 2,3 | - | 10,2 | 18,5 |
| Предшественник – пшеница после пара | |||||||||
| Экстенсивная | 647 | - | 587 | 10,5 | - | 6,7 | 22,2 | - | 31,2 |
| Нормальная | 722 | - | 385 | 12,2 | - | 3,8 | 23,8 | - | 7,2 |
| Интенсивная | 1088 | - | 337 | 12,8 | - | 3,8 | 45,5 | - | 27,2 |
| НСР0.5 | 245 | - | 327 | 4,7 | - | 6,8 | 13,7 | - | 20,8 |
Таблица 7 – Влияние технологии возделывания яровой пшеницы на
численность видов семейств Syrphidae, Coccinellidae, Chrysopidae
| Технология возделывания | Сем. Chrysopidae | Сем. Coccinellidae | Сем. Syrphidae | ||||||
| 2005 | 2006 | 2007 | 2005 | 2006 | 2007 | 2005 | 2006 | 2007 | |
| Предшественник – пар | |||||||||
| Экстенсивная | - | 17,2 | 13,3 | - | 15,5 | 13,3 | - | 4,5 | 0,5 |
| Нормальная | - | 11,2 | 7,2 | - | 17,2 | 11,7 | - | 2,2 | 1,7 |
| Интенсивная | - | 17,2 | 13,8 | - | 30,0 | 23,8 | - | 7,8 | 5,0 |
| НСР0.5 | - | 8,7 | 9,0 | - | 9,8 | 10,2 | - | 3,2 | 2,2 |
| Предшественник – пшеница после пара | |||||||||
| Экстенсивная | 21,2 | - | 15,5 | 16,2 | - | 6,2 | 8,8 | - | 3,3 |
| Нормальная | 17,2 | - | 7,2 | 24,5 | - | 20,0 | 8,3 | - | 1,2 |
| Интенсивная | 23,8 | - | 13,8 | 35,5 | - | 24,5 | 15,5 | - | 6,2 |
| НСР0.5 | 5,2 | - | 13,5 | 14,3 | - | 8,7 | 5,5 | - | 2,5 |
(экз./50 взмахов сачком)
Максимальное обеспечение растений необходимыми элементами питания, применение пестицидов, способствует активным ростовым процессам и удлиняет период вегетации культуры, чем благоприятствует развитию и размножению вредных организмов, в том числе злаковых тлей. Так, в 2005 и 2006 гг. суммарная численность тлей за вегетацию на полях пшеницы интенсивного возделывания, где растения отличаются мощной вегетативной массой и колосом, составляла в среднем 1088 и 662 особей/50 взмахов сачком, что в 1,7-1,9 раз больше, чем при экстенсивном возделывании, и в 1,4-1,5 раз больше, чем на посевах с нормальной технологией возделывания. В 2007 г. разница в численности вредителей на всех вариантах опыта была недостоверна.
Из хищных энтомофагов менее всего реагируют на технологии возделывания пшеницы клопы набисы и златоглазки. Отличий в плотности их популяции на опытных полях не найдено. Возделывание пшеницы по интенсивной технологии приводит к достоверному росту количества кокцинеллид, сирфид, первичных паразитов злаковых тлей. Суммарная численность кокцинеллид за период вегетации в этом случае достигает в среднем 23,8-35,5 особей/50 взмахов сачком, что превышает соответствующий показатель на вариантах с экстенсивной и нормальной технологиями в 1,2-4 раза. Количество личинок сирфид за вегетационный период на полях интенсивного типа составляет 5,0-15,5 особей/50 взмахов, что в 1,6-10 больше, чем при выращивании культуры по экстенсивной и нормальной технологиям. Численность имаго первичных паразитов на пшенице, возделываемой по интенсивной технологии, составляет 30,5-40,5 особей/50 взмахов и в 1,4-8 раз выше в сравнении с экстенсивной и нормальной технологиями (табл. 8). Обилие клопов ориусов в одни годы (2005, 2007 – пшеница по пару) на посевах с интенсивной технологией возделывания пшеницы больше, чем на посевах с экстенсивной технологией, в другие (2006, 2007 – пшеница по пшенице) – различается незначительно.
Таблица 8 – Влияние технологии возделывания яровой пшеницы на
численность первичных паразитов злаковых тлей, суммарное количество
хищных энтомофагов (экз./50 взмахов сачком)
| Технология возделывания | Первичные паразиты тлей (имаго) | Суммарная численность хищных энтомофагов | ||||
| 2005 | 2006 | 2007 | 2005 | 2006 | 2007 | |
| Предшественник – пар | ||||||
| Экстенсивная | - | 15,5 | 13,8 | - | 60,5 | 46,2 |
| Нормальная | - | 21,2 | 21,7 | - | 52,2 | 48,3 |
| Интенсивная | - | 30,5 | 38,8 | - | 90,5 | 86,2 |
| НСР0.5 | - | 11,3 | 16,3 | - | 26,0 | 36,3 |
| Предшественник – пшеница после пара | ||||||
| Экстенсивная | 21,2 | - | 4,5 | 78,8 | - | 62,8 |
| Нормальная | 22,8 | - | 8,8 | 87,2 | - | 39,5 |
| Интенсивная | 40,5 | - | 36,2 | 133,3 | - | 75,5 |
| НСР0.5 | 8,7 | - | 5,2 | 22,5 | - | 31,5 |
Суммарная численность всех хищных энтомофагов за вегетационный период на пшенице, выращиваемой по нормальной и экстенсивной технологиям, составляет 39,5-87,2 и 46,2-78,8 особей/50 взмахов сачком, что ниже, чем на полях интенсивного типа в среднем, соответственно, в 1,5-1,9 и 1,2-1,9 раз.
Таким образом, в условиях лесостепи Западной Сибири возделывание яровой пшеницы по интенсивной технологии не приводит к нарушению экологического равновесия в агроценозе. Если численность вредителей и увеличивается, то параллельно с ними возрастает численность энтомофагов, что стабилизирует энтомокомплекс и обеспечивает оптимизацию фитосанитарной обстановки.
Вышеизложенное подтверждает мнение Н.А. Вилковой, В.И. Танского и других ученых (Вилкова, Танский, 1994; Танский, 1997; Танский и др., 2001) о том, что агроценозы – системы достаточно устойчивые и способны в значительной мере смягчить влияние на них средств химизации, при условии рационального их применения. Агротехнические мероприятия, оказывая сильное влияние на отдельные компоненты агроэкосистем, не нарушают их целостности, не приводят к росту напряженности в них трофических взаимоотношений.
ВЫВОДЫ
1. В лесостепи Западной Сибири в агроценозах злаковых культур сохранены биоценотические механизмы регуляции плотности популяций злаковых тлей.
2. Уточнен состав комплекса энтомофагов яруса травостоя, снижающих численность тлей на зерновых культурах и черемухе – первичном хозяине черемухово-злаковой тли, насчитывающий к настоящему времени 134 вида насекомых. Из них 89 видов (66,4%) являются энтомофагами злаковых тлей, 45 (33,6%) – паразитами этих энтомофагов. Из энтомофагов 75 видов (84,3%) – хищники, 14 (15,7%) – паразиты. Вновь отмеченными видами являются паразиты златоглазок Helorus anomalipes (Panzer) и Baryscapus principiae Damenichini. B. principiae впервые зарегистрирован в фауне Западной Сибири.
3. Видовой состав энтомофагов злаковых тлей в центрально-лесостепном Верхнекарасукском и южно-лесостепном Барабинском агроландшафтных районах Новосибирской области идентичен видовому составу энтомофагов злаковых тлей в центрально-лесостепном Приобском агроландшафтном районе. Различия наблюдаются только в количественном соотношении видов, которое меняется во времени и пространстве.
4. Выявлены доминирующие виды хищных и паразитических энтомофагов. Среди имаго кокцинеллид до 1990 г. доминировали Coccinella septempunctata L., Propylaea quatuordecinapunctata L. и Hippodamia tredecimpunctata L., в дальнейшем в агроценозе озимой ржи по численности стала преобладать Adonia variegatа Goeze, пшеницы и овса – P. quatuordecimpunctata. Среди хищных набид доминирующим видом является N. ferus L., сирфид – Episyrphus balteatus Deg., Scaeva pyrastri L., Sphaerophoria scripta L, Melanostoma mellinum L., мухи рода Platycheirus Lep. and Serv.; златоглазок – Chrysopa carnea Steph., Ch. phyllochroma Wesm; паразитических перепончатокрылых – Aphidius rhopalosiphi De Stefani, Praon volucre Hal., Aphelinus transversus Thomson. Изучены биоэкологические особенности наиболее массовых видов энтомофагов. Определена роль паразитов в снижении численности полезных видов.
5. Сезонная и многолетняя динамика численности энтомофагов в агроценозах зерновых культур определяется их видовыми особенностями, погодными условиями, зараженностью энтомофагов паразитами, для некоторых перепончатокрылых – видовым составом тлей и т.д. Существенное влияние на динамику численности полезных видов оказывает плотность популяций злаковых тлей. Установлена тесная прямая связь между суммарной численностью злаковых тлей и энтомофагов за период вегетации (в агроценозе озимой ржи: r = +0,88, яровой пшеницы и овса: r = +0,80), а также во время максимальной плотности популяций вредителей (на озимой ржи: r = +0,72, на яровых культурах: r = +0,86). На посевах яровых зерновых наиболее сильно эта связь выражена у кокцинеллид и сирфид (в целом за период вегетации, соответственно, r = +0,89 и +0,50, во время максимальной плотности популяций вредителей: r = +0,93 и +0,85), на озимой ржи – у клопов набисов (за период вегетации и во время максимальной плотности популяций вредителей: r = +0,88). При низкой заселенности посевов зерновых злаковыми тлями численность многих видов энтомофагов здесь также остается низкой, размножение их незначительное либо вовсе не происходит.
6. Наибольшее значение в снижении плотности популяций злаковых тлей в агроценозах зерновых культур играют хищники, а из них – представители сем. Coccinellidae, в травостое озимой ржи, кроме этого, – хищные клопы рода Nаbis. Роль личинок сирфид и видов сем. Chrysopidae в комплексе энтомофагов злаковых тлей увеличивается в годы с высокой их численностью в агроценозах. Влияние паразитических наездников значительно осенью на озимой ржи, где ими может быть заражено до 87% большой злаковой тли, мигрирующей на всходы для откладки зимующих яиц. Уточнен критерий эффективности энтомофагов в лесостепи Западной Сибири.
7. Большое значение в повышении численности энтомофагов играют системы полезащитных насаждений, особенно если к ним прилегают полосы незапаханной земли, поляны дикорастущего разнотравья, многолетние и двулетние травы: люцерна, донник, эспарцет, клевер, кострец безостый. Они являются не только местом зимовки энтомофагов, но и постоянным местом обитания и размножения многих видов кокцинеллид, сетчатокрылых, хищных клопов набисов и ориусов, имаго сирфид и паразитических перепончатокрылых.
8. Предпочитаемым местом обитания и размножения кокцинеллид и златоглазок являются люцерна, донник, клевер, смесь многолетних трав с сорной растительностью, на костреце личинки кокцинеллид не найдены, личинки златоглазок – единичны. Клопы рода Orius обитают на всех изучаемых многолетних и двулетних травах, местом их массового размножения является люцерна, единично они встречаются на костреце и дикорастущем разнотравье. Для Nabis ferus, N. punctatus предпочитаемым местообитанием являются кострец, люцерна, клевер, для N. flavomarginatus – кострец и дикорастущее разнотравье, N. brevis – дикорастущее разнотравье. Больше всего имаго паразитических наездников отмечено на люцерне, смеси многолетних трав с сорной растительностью, дикорастущем разнотравье. В течение всей вегетации цветущую растительность посещают сирфиды. Однако размножение их на многолетних травах и дикорастущем разнотравье не происходит, либо бывает незначительным. Возможно микроклимат, создающийся в загущенных посевах травянистой растительности для них неблагоприятен.
9. Суммарное количество злаковых тлей и их энтомофагов за вегетацию зависит от вида культуры, на которой они развиваются (степень влияния фактора, соответственно, 56-78 и 48-77%). На отдельные группы энтомофагов, при низкой их численности, вид культуры не оказывает влияния, при значительной – влияние от среднего до высокого (на кокцинеллид – до 44%, златоглазок – до 67, сирфид – до 46, ориусов – до 79, набисов – до 19, имаго первичных паразитов – до 67%). Плотность популяций тлей на пшенице после пара, посевах овса и вико-овса в 1,9-12,9 раз больше, чем на озимой ржи по пару и по доннику, рапсу, ячменю и ячменю с донником. Обилие хищных энтомофагов яруса травостоя, снижающих численность вредителей, также ежегодно поддерживается на высоком уровне на посевах пшеницы после пара, овсе и вико-овсе.
10. Из основных элементов агрокомплекса возделывания яровой пшеницы наибольшее влияние на злаковых тлей и их энтомофагов оказывают внесение минеральных удобрений и пестицидов, место культуры в севообороте. На пшенице, выращиваемой с внесением сбалансированного минерального питания и пестицидов, численность тлей в 1,9-3,5 раза превышает их количество на пшенице, возделываемой с внесением одних удобрений, и в 1,7-5,9 раза – без внесения средств химизации (степень влияния фактора 65-83%). Средняя численность злаковых тлей на пшенице по пару в 1,3 раза больше их численности на четвертой пшенице после пара (степень влияния фактора 3-25%).
11. Применение комплекса средств химизации наиболее сильно влияет на обилие видов сем. Coccinellidae (25-55%) и Syrphidae (65-71%), суммарную численность энтомофагов (42-50%), а также имаго перепончатокрылых паразитов (55-62%). Место культуры в севообороте – на обилие кокцинеллид (10-24%), клопов набисов (22-28%) и ориусов (25-39%), суммарную численность энтомофагов (18-20%). Влияние изучаемых факторов на обилие энтомофагов опосредуется их воздействием на численность злаковых тлей, реже – предпочтением определенного местообитания. Количество хищных энтомофагов на пшенице, возделываемой с комплексным применением удобрений и пестицидов, в 1,9-2,5 раза больше, чем на пшенице, выращиваемой с внесением одних удобрений и в 1,4-2,8 раз – без удобрений и пестицидов. Средняя суммарная численность хищников за вегетацию на первой пшенице после пара в 1,4-2 раза больше, чем на четвертой пшенице после пара.
12. Система основной обработки почвы не влияет на численность злаковых тлей, хищных и паразитических энтомофагов, которая в среднем варьирует, соответственно, от 155 до 181 особи тли/10 взмахов за вегетацию (степень влияния фактора 3-4%), от 20,3 до 24,0 особей хищников/10 взмахов (степень влияния 2-3%) и от 6,2 до 7,3 особей имаго паразитов (степень влияния 4-6%).
13. В условиях сухой жаркой погоды и при обработке инсектицидом, проведенной в фазу кущения пшеницы, численность злаковых тлей на бессменной пшенице минимальная, также как на пшенице по рапсу, ячменю и вико-овсу, достоверно больше (в 6,6-9,3 раза) вредителей отмечается по лучшим для пшеницы предшественникам – пару, пшенице после пара, озимой ржи, а также овсу (степень влияния фактора 44,4%). При благоприятных условиях для развития злаковых тлей достоверных различий в их обилии на посевах пшеницы по различным предшественникам и на бессменной пшенице нет (степень влияния фактора 0,0-4,3%). Предшественники не оказывают влияние, либо оказывают среднее влияние на обилие в травостое пшеницы кокцинеллид, златоглазок, сирфид. Отмечено явное предпочтение ориусов посевов бессменной пшеницы. В отдельные годы достоверно больше набисов и имаго паразитических наездников отмечается на пшенице по пару. Высокий уровень суммарной численности всех хищных энтомофагов ежегодно поддерживается на посевах пшеницы после пара и на бессменной пшенице.
14. В лесостепи Западной Сибири возделывание пшеницы по интенсивной технологии не приводит к нарушению экологического равновесия в агроценозе. При увеличении численности вредителей возрастает обилие энтомофагов, что стабилизирует энтомокомплекс и обеспечивает оптимизацию фитосанитарной обстановки.
ПРАКТИЧЕСКИЕ РЕКОМЕНДАЦИИ
Для сохранения регуляторной способности ценозов и эффективной деятельность хищников и паразитических перепончатокрылых необходимо повышение биоразнообразия агроэкосистем, в том числе их флористического разнообразия, что возможно путем создания систем полезащитных насаждений, живых изгородей, оптимизированных по густоте и видовому составу, сети участков с повышенным биоразнообразием или агроландшафтных заповедных зон. В лесостепи Западной Сибири создание их возможно в многочисленных колках, являющихся резерватами многих видов энтомофагов.
Для повышения биоразнообразия и обогащения фауны агроценозов зерновых полезными видами необходима организация длинно-ротационных и разнообразных по набору культур севооборотов, введение в севооборот и возделывание рапса ярового, люцерны и других многолетних трав, посевов непрерывно цветущих фуражных и нектароносных растений.
При соотношении хищник : жертва на озимой ржи 1:30, яровых зерновых 1:40 не целесообразно проводить химические обработки посевов зерновых против злаковых тлей, так как их численность сдерживается энтомофагами и не превышает порога экономической вредоносности.
Химические обработки не рекомендуется проводить в период пика активности наиболее массовых и эффективных энтомофагов. Наименее опасно применение химических средств защиты на начальных этапах роста численности вредителей, до массового появления их природных врагов. Если возникает необходимость обработки в более позднее время, нужно выбирать такие сроки, когда основные виды энтомофагов находятся в устойчивой к действию инсектицидов фазе яйца, куколки, паразиты – внутри мумифицированных тлей. Растения обрабатывают в безветренную погоду после 17-18 ч, когда наблюдается спад летной активности насекомых. Посевы зерновых необходимо обрабатывать до укоса многолетних трав, являющихся резерватом полезной энтомофауны.
Необходимо обучение биологизации и экологизации защиты растений специалистов всех уровней.
СПИСОК ОСНОВНЫХ РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ
ДИССЕРТАЦИИ
Статьи в изданиях, рекомендованных ВАК РФ:
1. Кротова И.Г. Хищные левкописы (Diptera, Chamaemyiidae), обитающие на посевах зерновых в северной лесостепи Приобья / И.Г. Кротова // Сибирский вестник с.-х. науки. – 1993. – № 3. – С. 58-59.
2. Кротова И.Г. Паразиты злаковых тлей и их роль в снижении численности вредителей на посевах зерновых культур в северной лесостепи Приобья Западной Сибири / И.Г. Кротова // Зоологический журнал. – 1993. – Т. 72. – Вып. 12. – С. 51-57.
3. Кротова И.Г. Паразиты сирфид (Diptera, Syrphidae) – афидофагов на посевах зерновых культур в Западной Сибири / И.Г. Кротова // Зоологический журнал. – 1993. – Т. 72. – Вып. 12. – С. 58-62.
4. Кротова И.Г. К биологии черемуховой тли Rhopalosiphum padi (L.) (Homoptera, Aphididae) и ее паразита Aphelinus transversus Thomson (Hymenoptera, Aphelinidae) на посевах зерновых культур в северной лесостепи Приобья // Энтомологическое обозрение. – 1994. – Т. 73. – № 2. – С. 249-254.
5. Власенко Н.Г., Кротова И.Г., Власенко А.Н. Влияние средств химизации на полезную энтомофауну агроценоза яровой пшеницы / Н.Г. Власенко, И.Г. Кротова, А.Н. Власенко // Агрохимия. – 1996. – № 2. – С. 97-101.
6. Бокина И.Г. Вторичные паразиты злаковых тлей в северной лесостепи Приобья Западной Сибири / И.Г. Бокина // Зоологический журнал. – 1997. – Т. 76. – № 4. – С. 432-437.
7. Бокина И.Г. Энтомофаги злаковых тлей в Западной Сибири / И.Г. Бокина // Защита и карантин растений. – 1999. – № 7. – С. 13-14.
8. Каличкин В. Формирование агроэкологических условий для агроценозов яровой пшеницы / В. Каличкин, Г. Захаров, Т. Крупская, М. Бекасова, В. Шоба, И. Бокина // Сибирский вестник сельскохозяйственной науки. – 2003. – № 4. – С. 3-11.
9. Бокина И.Г. Севооборот и вредители яровой пшеницы // Защита и карантин растений. – 2005. – № 6. – С. 20-21.
10. Бокина И.Г. К биологии клопов-антокорид (Heteroptera, Anthocoridae) – энтомофагов злаковых тлей в лесостепи Западной Сибири / И.Г. Бокина // Энтомологическое обозрение. – 2008. – Т. 87. – № 4. – С. 736-740.
11. Бокина И.Г. От чего зависит размножение злаковых тлей и их энтомофагов / И.Г. Бокина // Защита и карантин растений. – 2008. – № 11. – С. 21-22.
12. Каличкин В.К. Фитосанитарное состояние посевов пшеницы в зависимости от технологий ее возделывания / В.К. Каличкин, И.Г. Бокина, С.А. Ким, И.Н. Минина, В.Н. Шоба // Достижения науки и техники АПК. – 2008. – № 4. – С. 27-29.
13. Бокина И.Г. Роль и характер растительности и численность энтомофагов злаковых тлей в лесостепи Западной Сибири / И.Г. Бокина // Зоологический журнал. – 2009. – Т. 88. – №. 8. – С.
Научные статьи, тезисы, монография:
1. Коробов В.А. Фитосанитарное состояние посевов яровой пшеницы по вредителям при интенсивной технологии возделывания / В.А. Коробов, И.Г. Кротова // Интенсификация возделывания зерновых культур в Западной Сибири / Сб. науч. тр. / СибНИИЗХим. – 1990. – С. 114-126.
2. Кротова И.Г. Паразиты злаковых тлей в северной лесостепи Приобья / И.Г. Кротова // Проблемы аграрной науки в условиях перехода к рынку: Тез. докл. науч.-практ. конф. проф.-преп. коллектива, науч. сотр., аспирантов Новосиб. гос. агр. ун-та и работников пр-ва (23-25 мая 1991). – Новосибирск. – 1991. – С. 134-136.
3. Кротова И.Г. Анализ взаимовлияния популяций злаковых тлей и их энтомофагов на посевах озимой ржи / И.Г. Кротова // Защита с.-х. культур от болезней и вредителей в Сибири: Сб. науч. тр. РАСХН. Сиб. Отделение. СибНИИЗХим. – Новосибирск, 1992. – С. 147-153.
4. Бокина И.Г. Влияние системы обработки почвы и средств химизации на злаковых тлей и их энтомофагов в агроценозе яровой пшеницы в Западной Сибири / И.Г. Бокина // Вестник защиты растений. – 2006. – № 2. – С. 25-33.
5. Бокина И.Г. Влияние предшественников на численность злаковых тлей и их энтомофагов в северной лесостепи Приобья / И.Г. Бокина // Вестник защиты растений. – 2007. – № 2. – С. 44-54.
6. Бокина И.Г. Кокцинеллиды (Coleoptera, Coccinellidae) в агроценозе зерновых культур в Западной Сибири / И.Г. Бокина // Достижения энтомологии на службе агропромышленного комплекса, лесного хозяйства и медицины: Тез. докл. XIII съезда РЭО (Краснодар, 9-15 сентября 2007 г.). – Краснодар, 2007. – С. 35.
7. Бокина И.Г. Влияние технологии возделывания на вредную и полезную фауну в агроценозе яровой пшеницы в лесостепи Западной Сибири / И.Г. Бокина // Аграрная наука – сельскому хозяйству: Сб. статей. III Международная науч.-практич. конф. – Барнаул: Изд. АГАУ, 2008. – С. 209-211.
8. Бокина И.Г. Экологическая оценка интенсивной технологии возделывания яровой пшеницы в лесостепи Западной Сибири / И.Г. Бокина // Современные средства, методы и технологии защиты растений: Материалы международной научно-практической конференции (Новосибирск, 10-11 июля 2008 г.). – Новосибирск, 2008. – С. 26-30.
9. Каличкин В.К. Влияние удобрений и средств защиты растений на урожайность яровой пшеницы / В.К. Каличкин, С.А. Ким, И.Г. Бокина, И.Н. Минина // Аграрная наука – сельскому хозяйству: Сб. статей. IV Международная науч.-практич. конф. – Барнаул: Изд. АГАУ, 2009. – Кн. 2. – С. 386-388.
10. Бокина И.Г. Эффективность энтомофагов в снижении численности злаковых тлей в Западной Сибири / И.Г. Бокина // Аграрная наука – сельскохозяйственному производству Казахстана, Сибири и Монголии: Тр. XII Международной науч.-практич. конф. (Шымкент, 16-17 апреля 2009 г.). – Алматы: Изд. «Бастау». – Т. 1. – С. 314-315.
11. Бокина И.Г. Злаковые тли и их энтомофаги в лесостепи Западной Сибири / И.Г. Бокина. – Новосибирск, 2009. – 182 с.
12. Бокина И.Г. Рациональное использование химических средств защиты растений как условие эффективной деятельности энтомофагов и сохранения экологического равновесия в агроэкосистемах / И.Г. Бокина // Проблемы экологии агроэкосистем: пути и методы их решения: Материалы Всероссийской научной конференции. – Новосибирск, 2009.