

Биоэкологическое обоснование защиты семенников капусты от основных вредителей в условиях юга россии
На правах рукописи
Мисриева Бичихан Усмановна
Биоэкологическое обоснование защиты семенников капусты от основных вредителей в условиях юга России
Специальность 06.01.11. - защита растений
Автореферат
диссертации на соискание учёной степени
доктора сельскохозяйственных наук
Москва - 2008
Работа выполнена в Российском государственном аграрном университете – МСХА имени К.А. Тимирязева (г. Москва) и на Дербентской селекционно-опытной станции виноградарства и овощеводства (Республика Дагестан)
Научный консультант: доктор биологических наук, профессор С.Я.Попов.
Официальные оппоненты:
Соколов Михаил Сергеевич, академик РАСХН, доктор биологических наук, профессор
Словцов Ростислав Иванович, доктор сельскохозяйственных наук, профессор
Радченко Евгений Евгеньевич, доктор биологических наук
Ведущая организация: Российский университет дружбы народов (г. Москва)
Защита диссертации состоится ……………….. в ……. час на заседании диссертационного совета Д.220.043.04 при Российском государственном аграрном университете – МСХА имени К.А.Тимирязева.
Адрес: 127550, Москва, ул. Тимирязевская, 49, учёный совет РГАУ – МСХА.
С диссертацией можно ознакомиться в библиотеке Российского государственного аграрного университета – МСХА имени К.А. Тимирязева.
Автореферат разослан ………………… г.
Размещён на сайте ВАК www vak. ed. gov.ru
Учёный секретарь
диссертационного совета, доцент А.Н. Смирнов
Общая характеристика работы
Актуальность исследований. Юг России является крупнейшим производителем белокочанной капусты. Стратегия овощеводства, предусматривающая обеспечение в полном объёме потребностей населения в овощах, в значительной степени направлена на создание и совершенствование семеноводческой базы. Однако в связи с нарушением хозяйственно-экономических связей между овощными семеноводческими сельскохозяйственными предприятиями юга России, производство семян овощных культур в последние годы значительно сократилось. По данным ВНИИО, за последние 10 лет в целом по стране производство сортовых семян снизилось со 125 до 9 тыс. тонн, около 5 тыс. тонн семян импортируется в Россию из-за рубежа (Р. А. Гиш, 2003).
В последние годы семеноводство гибридов белокочанной капусты концентрируется в регионах, позволяющих использовать беспересадочный способ, обеспечивающий значительное снижение затрат на производство семян. В России наиболее благоприятным местом для промышленного семеноводства белокочанной капусты является южный Дагестан. Несмотря на высокую экономическую эффективность используемой прогрессивной технологии получения семян, есть немало причин, вызывающих ежегодные потери урожая. Потенциальная урожайность семенников белокочанной капусты в условиях юга России может составлять 1,2 ц и более с 1 гектара. Однако средняя урожайность культуры в последние годы нередко не превышает 0,2-0,3 ц/га. Основной причиной недобора урожая семян белокочанной капусты является вредоносная деятельность фитофагов, способных в годы их массового распространения полностью уничтожить урожай. К числу наиболее опасных вредителей семенников капусты относятся брюквенный барид (Baris coerulescens Scop.), капустная тля (Brevicoryne brassicae L.), рапсовый цветоед (Meligethes aeneus Fab.), крестоцветные блошки (Phyllotreta) и капустная моль (Plutella maculipennis Curt.).
К началу наших исследований в отношении семенной капусты, выращиваемой по беспересадочной технологии, вопросы защиты от фитофагов не освещались. Информация о популяционной динамике и биоэкологических особенностях названных вредителей была скудной и недостаточной для формирования стратегии и тактики защиты семенников капусты от вредителей. Проводимые из года в год защитные мероприятия сводились к календарным обработкам, назначаемым без учета реальной фитосанитарной ситуации. Из-за полного отсутствия экономических порогов вредоносности основных видов вредителей семенников капусты, применение химических средств защиты растений не обосновывалось. Не учитывались также роль и степень воздействия их на энтомофагов, как значимого звена интегрированной защиты растений. Решению этих актуальных проблем и посвящается настоящая исследовательская работа.
Цель и основные задачи. Цель работы - разработка научно-обоснованной интегрированной защиты семенников капусты от вредителей, основанной на изучении видового состава, экологических свойств фитофагов, их вредоносности и экономических порогов вредоносности, природных энтомофагов, способных эффективно ограничивать численность популяций вредителей семенников капусты в условиях юга России.
Для реализации поставленной цели потребовалось решение следующих задач:
- Провести ревизию и уточнение видового состава вредителей семенников капусты в условиях юга России;
- Изучить сезонную и многолетнюю динамику численности и вредоносности основных вредителей семенников капусты, их фенологические особенности и сопряженность развития с кормовым растением;
- Изучить популяционную экологию вредителей семенников капусты с использованием К-факторных таблиц выживания;
- Выявить роль и значение энтомофагов в регулировании численности
популяций вредителей семенников капусты;
- Разработать систему фитосанитарного мониторинга семенников капусты в отношении основных видов вредителей;
- Оценить существующие и разработать новые (усовершенствованные) мероприятия по защите семенников капусты от вредителей;
- Разработать интегрированную систему защиты семенников капусты с учетом её экономической эффективности.
Научная новизна и практическая значимость результатов исследований
На основании обширного фактического материала, полученного в результате десятилетнего фитосанитарного мониторинга, впервые приводится список видов фитофагов и энтомофагов в агроценозе семенной белокочанной капусты, выращиваемой по беспересадочной технологии, изучены и уточнены особенности их биологии, экологии и вредоносности в условиях южной оконечности России (Южного Дагестана);
- впервые проведена видовая идентификация сообщества баридов в Дагестане, выявлен доминант - Baris coerulescens Scop., который является наиболее опасным вредителем семенников капусты;
- впервые в условиях южного Дагестана проанализирована популяционная динамика основных вредителей семенников капусты с использованием K-факторных таблиц выживания;
- на основе разработанной модели предложена система прогнозирования, позволяющая устанавливать периоды возникновения вспышек массового размножения и распространения брюквенного барида;
- впервые в условиях Дагестана установлена роль основных энтомофагов в ограничении численности вредителей семенников капусты. По итогам многолетних учетов отмечено влияние афидофага Diaeretiella rapae (M'lntosh) на динамику численности капустной тли. Из комплекса паразитоидов капустной моли наиболее активны наездники Diadegma collaris Grav., D. semillausum Hell. и D. fenestralis Holm., относящиеся к семейству Icheneumonidae, заражающие до 70% числа гусениц капустной моли;
- с учетом изученных факторов вредоносности разработаны и уточнены экономические пороги вредоносности брюквенного барида, рапсового цветоеда, капустной тли и крестоцветных блошек применительно к семенной культуре капусты;
- впервые для семенников белокочанной капусты предложена схема интегрированной системы защиты от основных видов вредителей.
Основные положения, выносимые на защиту:
Особенности региональной (в условиях Дагестана) популяционной динамики численности основных фитофагов семенной капусты являются теоретической и методологической основой для построения интегрированной системы защиты семенников капусты от вредителей;
Новые пороги вредоносности основных вредителей семенников капусты как фактор эколого-экономической целесообразности применения истребительных мероприятий;
Биологическое обоснование и построение системы интегрированной защиты семенников капусты от основных вредителей на основе глубокого изучения ключевых моментов их экологических свойств и вредоносности.
Апробация работы. Экспериментальные и теоретические исследования выполнялись в рамках государственной НИР по контракту МСХА им. К.А.Тимирязева с Министерством сельского хозяйства и продовольствия РФ от 5.10. 2001г., № 2074: «Проведение исследований по разработке новых методов семеноводства позднеспелой гибридной кочанной капусты, полученной на основе мужскостерильных линий», а также в рамках научно-исследовательской работы ФГНУ «Дербентская селекционно-опытная станция виноградарства и овощеводства». Материалы диссертации представлялись на 12 международных научно - практических конференциях в том числе: Международной научной конференции "Селекция и семеноводство овощных культур в XXI в." (Москва, 24-27.VI. 2000г.), международной научно-практической конференции «Проблемы научного обеспечения овощеводства юга России» (Краснодар, 4-7 IX. 2004г.), Всероссийской научно-практической конференции «Интегрированная защита сельскохозяйственных культур и фитосанитарный мониторинг в современном земледелии» (Ставрополь, 15-19.XI 2004г.), международной научной конференции «Актуальные проблемы земледелия, защиты растений, генетики, селекции и семеноводства овощных культур» (Москва XII. 2003 г.), международной научно-практической конференции «Современное состояние овощеводства» (Краснодар, КНИИОКХ, VIII, 2006г.), IX международной конференции «Биологическое разнообразие Кавказа» (Махачкала 5-6. XI 2007г.). Основные результаты исследований ежегодно докладывались на заседаниях и совещаниях ФГНУ ДСОСВ и О (Дербентской селекционно-опытной станции виноградарства и овощеводства), ФГУ «Территориальная станция защиты растений» по РД. Результаты исследований неоднократно докладывались также в РГАУ – МСХА имени К.А. Тимирязева.
Публикации. Основные результаты исследований опубликованы в 23 научных статьях, в том числе 10 в журналах, рекомендованных ВАК РФ.
Структура и объём диссертации. Диссертация изложена на 269 печатных страницах. Состоит из введения, 8 глав, заключения, выводов, предложений производству, приложений, 46 таблиц, 32 рисунков, в том числе 9 фотографий. Список использованной литературы включает 237 наименований, в том числе 48 иностранных авторов.
Участие в работе. Диссертационная работа выполнена непосредственно соискателем и является самостоятельным завершённым трудом. В выполнении экспериментальной части в различные годы нам оказывали помощь сотрудники Дагестанского Института прикладной экологии, Дербентской межрайонной станции защиты растений. Автор глубоко признателен своему научному консультанту профессору Попову С.Я. за ценные советы и содействие в выполнении направлений исследований, а также искреннюю благодарность директору Селекционной станции им. Н.Н. Тимофеева Г.Ф. Монахос, гл. специалисту ЗАО «Сортсемовощ» РД Т.Х. Касимову за помощь в постановке проблемы и сотрудничество.
Содержание работы
Введение. Во вводной части диссертации обоснована актуальность решения важной народно-хозяйственной проблемы – снижения потерь урожая и качества семян от особо опасных вредителей семенников капусты. Показана научно-практическая значимость исследуемой темы, сформулированы цели и задачи направлений исследований.
Глава 1. Состояние изученности вопроса и задачи исследований (обзор литературы)
В главе приведён аналитический обзор литературных источников по изучаемой проблеме, дан анализ адаптивных технологий возделывания семенной капусты, позволяющих эффективно сокращать плотность популяций основных вредителей. Проанализированы имеющиеся в источниках литературы концепции экономических порогов вредоносности (ЭПВ).
Анализ литературных источников свидетельствует о недостаточном информационном обеспечении системы защиты семенников капусты. Для разработки интегрированной защиты в соответствии с современными требованиями необходимы исследования, позволяющие дать им теоретическое обоснование. На основании анализа показано, что в отечественной и зарубежной литературе вопросы защиты семенников капусты мало освещены, имеющиеся публикации не затрагивают существо многих проблем. Всё это ориентирует на необходимость поиска конкретных путей решения задач по разработке теоретически обоснованной системы защиты семенников капусты в условиях юга России. В настоящей квалификационной работе автором сформулированы основные направления исследований.
Глава 2. Условия проведения, материалы и методы исследований. Климатическая характеристика вегетационных сезонов в годы исследований
В главе дана подробная характеристика климатических условий южно-плоскостной зоны Дагестана, где были проведены исследования. Агроклиматические условия южной зоны Дагестана относятся к зоне сухих субтропиков, являющихся весьма благоприятными для производства продовольственной и семенной белокочанной капусты.
Экспериментальная работа выполнялась в течение 10 лет – в период с 1998 по 2007 гг. Регулярные наблюдения велись на стационарных участках Дербентской селекционно-опытной станции виноградарства и овощеводства (ДСОСВ и О), Дагестанской опытной станции Всероссийского научно-исследовательского института растениеводства (ДОС ВНИИР), МУП агрофирма им. Г. Давыдовой, СПК «Колхоз им. Г. Казимова», агрофирма «Штул». Маршрутные обследования проводились в хозяйствах Дербентского района, культивирующих семенники капусты: МУП агрофирма «Низами», «Аглаби», «Джемикент», «Падар» и др. В указанных хозяйствах в разное время было заложено 26 стационарных опытных участка.
Методика проведения исследований. Мониторинг вредных и полезных видов вели в ходе регулярных обследований с использованием визуальных и инструментальных методов. Видовой состав и динамику численности наиболее распространенных видов устанавливали на основе проведения систематических учетов на стационарных участках в течение всего периода вегетации культуры. Наблюдения и эксперименты проводились с использованием лабораторных, вегетационных, полевых деляночных и производственных опытов по общепринятым методикам Г.Е. Осмоловского 1964, В.Ф. Палия (1966,1970), С.Р. Фасулати, 1971, А.С. Боголюбова, Е.А. Дунаева, (1996) и др. Изучение особенностей биологии и фенологии проводилось с использованием методик Б.В. Добровольского (1969). Для определения жизнеспособности популяций фитофагов использовали методики по составлению таблиц выживания, изложенные в работах Дж. К. Варли и др. (1978), а также С.Я. Попова (1981, 1983,1986).
Морфологические исследования насекомых сводились к микроскопии и сравнительному изучению таксономических признаков, с использованием стандартных стереомикроскопов. Видовой состав паразитоидов изучали методом индивидуального выведения из собранных в полевых условиях паразитированных хозяев по методикам С.В. Тряпицина, В.А. Шапиро, В.А. Щепетильникова (1969). Для учёта напочвенных беспозвоночных применялись ловушки Барбера (Barber, 1936). Идентификацию видов проводили по определителю О.Л. Крыжановского (1974). Определение видовой принадлежности насекомых подтверждено специалистами - систематиками ЗИН РАН: А.И. Халаим, А.Н. Рохлецовой, К.А. Надеин, М.Ш. Исмаиловой, И.А. Белоусовым, за что автор выражает признательность коллегам.
Объектами и материалами исследований были:
- наиболее распространенные и вредоносные фитофаги семенников белокочанной капусты: брюквенный барид (Baris coerulescens Scop.), капустная тля (Brevicoryne brassicae L.), рапсовый цветоед (Meligethes aeneus Fab.), капустная моль (Plutella maculipennis Curt.) и крестоцветные блошки (род Phyllotreta).
- энтомофаги, встречающиеся в агроценозе семенников капусты;
- инсектициды и биопрепараты, которые испытывались против вредителей семенников капусты.
Полевые мелкоделяночные и производственные опыты по определению биологической эффективности химических и биологических средств защиты проводили в соответствии с «Методическими указаниями по регистрационным испытаниям инсектицидов, акарицидов, моллюскоцидов и родентицидов в сельском хозяйстве» (ВИЗР, 2004). Статистическую обработку данных проводили по методикам, изложенным Б.А. Доспеховым (1985), а также с помощью пакета прикладных программ ПК.
Глава 3. Биофенологические особенности развития семенников белокочанной капусты в условиях южного Дагестана. На основе ежегодно проводимых фенологических наблюдений за семенниками белокочанной капусты получены среднемноголетние данные наступления и завершения каждой фенологической фазы растений. В условиях южного Дагестана начало сева культуры приходится на 10. VII ± 4.5 дней; массовый сев осуществляется 20. VII ± 3.5 дней; среднемноголетней датой высадки рассады является 16. IX ± 5.7 дней. Появление стебля и массовая бутонизация происходит следующей весной 14. IV ± 4.7 дней и 24. IV±4.1 дней соответственно. Начало цветения в условиях Дербентского района отмечается 24. IV ± 6.1 дней, массовое цветение - 12.V ± 5.1 дней. Образование стручков фиксируется 3.VI ± 9.8 дней. Массовая уборка осуществляется 14. VII ± 4.9 дней. Нами отмечена достаточно тесная взаимосвязь между пиками численности основных видов фитофагов и фазами развития семенников капусты, позволяющая планировать защитные мероприятия и добиваться их большей эффективности. Во время выращивания рассады, и вскоре после её высадки в грунт особенно опасны крестоцветные блошки. В фазе листовой мутовки и начала стрелкования культуры большой вред наносят капустная моль и капустная тля, в фазе бутонизации - рапсовый цветоед. В начале цветения семенников к ним присоединяются брюквенный барид и капустная тля.
Глава 4. Биоэкологические особенности развития и хозяйственное значение основных вредителей, повреждающих семенники белокочанной капусты. В главе дана краткая характеристика сезонных сроков развития крестоцветных блошек, капустной тли, капустной моли, брюквенного барида и рапсового цветоеда в условиях Южного Дагестана. Приводятся сведения о динамике заселения ими плантаций семенников капусты, показаны сезоны с максимальной их численностью. Данные по многолетней сезонной динамике развития основных вредителей семенников белокочанной капусты свидетельствуют о том, что в различные климатические сезоны плотность популяций фитофагов контрастна. Это связано, прежде всего, с биологическими особенностями развития фитофагов, их пищевой активностью в определенные фазы развития кормового растения. Полученные данные имеют как практический, так и теоретический интерес, поскольку могут служить основой для разработки прогнозов и оптимизации системы защитных мероприятий на семенниках капусты.
4.1. Биоэкологические особенности развития брюквенного барида (Baris coerulescens Scop.) на семенниках белокочанной капусты в условиях южного Дагестана
Брюквенный барид является для региона новым вредителем, встречающимся, в отличие от других фитофагов, только на семенниках капусты. Вредоносность брюквенного барида зависит от способа питания. Потребляя относительно небольшое количество пищи, личинки вредителя вызывают усыхание целого растения. К моменту уборки на массивах отмечаются значительные (до 70% и более) выпады растений.
Для рациональной организации мероприятий по борьбе с фитофагом необходимо знание его видовой принадлежности. В результате проведенных исследований нами было выявлено 22 вида, из которых 5 питается на растениях из семейства капустные, 2 - на маревых; по одному - на растениях из семейств мальвовых и злаковых, 2 вида - на Phlomis pungens и P. Tuberose; остальные виды были отмечены на следующих видах растений: Salcola dendroides Pall, Syrenia seliculosa Bieb, Comphorosma speliaca Z, Halocnemun strobilaceum Pall, Artemisia marshalliana, Pulicaria dysenterica, Sisymbrium и Reseda luteola (см. табл.1).
Таблица 1. Видовой состав баридов в Дагестане
| № п/п | Вид | Численность, % | Кормовое растение |
| 1 | Baris coerulescens Scop. | 71,60 | Капустные |
| 2 | B. timida Rossi | 6,10 | Мальвовые |
| 3 | Ulobaris loricata Boheman | 1,30 | Маревые |
| 4 | B. scolopacea Germar | 3,60 | Маревые |
| 5 | B. janthina Boheman | 3,10 | Капустные |
| 6 | B. melaena Boheman | 2,30 | Phlomis pungens |
| 7 | B. kirshi Desbrochers | 2,20 | Halocnemun strobilaceum Pall. |
| 8 | B. semistriata Boheman | 2,40 | Cardaria draba (L) Desv. |
| 9 | B. sulcata Bocheman | 1,20 | Comphorosma топ speliaca Z. |
| 10 | B. sibirica Faust | 1,04 | Poliantehes tuberose (L). |
| 11 | B. hochhuthi Faust | 0,90 | Syrenia seliculosa Bieb. |
| 12 | B. noaeae Becker | 0,90 | Salcola dendroides Pall. |
| 13 | B. carbonaria Boheman | 0,56 | Капустные |
| 14 | B. spitzyi Hochith | 0,40 | Artemisia marshalliana Spreng |
| 15 | B. analis Olivier | 0,40 | Artemisia marshalliana Spreng, сбоку чуть крупных сливающихся точках. Усиковые бороздки направлены косо к нижней стороне головотрубки и оканчиваются впереди глаз. Усики прикреплены в вершиной половине головотрубки. Рукоять усиков в основании слегка изогнутая, в два раза короче головотрубки. Первый членик жгутика усиков длиннее своей ширины, остальные поперечные. Булава усиков широкояйцевидной формы. Глаза невыпуклые и расположены по бокам основания головотрубки, овально- удлиненные. Края глаз не выдаются за контуры головной капсулы. Лоб чуть уже,чем основание головотрубки. Переднеспинка без заглазничьных лопастей, в основной половине более или менее параллельностороннняя, к вершине сильно сужена, основание двувыемчатое. Переднеспинка с тонко пунктированными участками, с гладкой срединной линией, у задних углов более тонко пунктированные участки. Щиток хорошо заметный, с выпуклостями по сторонам. Эпиплевры переднеспинки в грубых сливающихся в морщинки точках. |
| 16 | B. angusta Brulle | 0,40 | Sisymbrium officinale (L.) Scop |
| 17 | B. memnonia Boheman | 0.30 | Syrenia seliculosa Bieb. |
| 18 | Limnobaris dolorosa Gmail | 0,30 | Злаковые |
| 19 | B. picturata Menetrie | 0,30 | Капустные |
| 20 | B. lepidii Germar | 0,30 | Капустные |
| 21 | B. concinna Boheman | 0,20 | Reseda luteola L. (Weld) |
| 22 | B. limbata Brisout | 0,20 | Salcola dendroides Pall. |
Из приведенных данных видно, что среди баридов наиболее массовым является Baris coerulescens Scop. (71,6% к общей численности), В. timida Rossi занимает 6,1 % от всех видов, примерно в равных количествах B. scolopacea Germar, B. janthina Boheman (3,6 и 3,1%), B. kirshi Desbrochers, B. semistriata Boheman - 2,2 и 2,4% соответственно. Другие виды имели второстепенное значение. В разные годы заметных отклонений в структуре видового сообщества не наблюдалось.
Детальный агроклиматический анализ закономерностей развития брюквенного барида на семенниках капусты показал, что вегетационные сезоны 1998, 2003 и 2005гг. характеризовались относительно низкими показателями ГТК. Именно в эти сезоны отмечалась высокая вредоносность брюквенного барида. Проведённый анализ позволил утверждать, что интенсивное размножение вредителя происходило при ГТК 0,1-0,3. Максимальная плотность вредителя в эти периоды составляла от 32 до 37 экз. на 1 растение. В 1999, 2004 и 2006 гг. плотность вредителя была на уровне разработанного нами ЭПВ - т.е. не превышала 0,5 – 0,7 экземпляров вредителя на заселенное растение. Оптимальное развитие вредителя происходило при сумме эффективных температур, равной 1700°С (см. рис. 1).
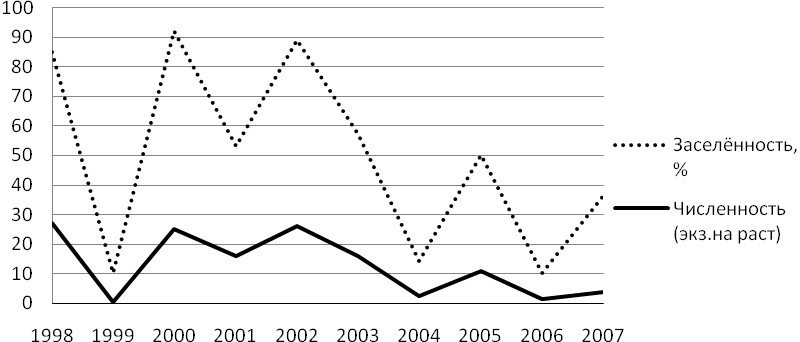
Рис. 1. Многолетняя сезонная динамика численности брюквенного барида на семенниках капусты. Дагестанская опытная станция ВНИИР.
Продолжительность откладки яиц у брюквенного барида иногда может продолжаться до 3 месяцев. В Южном Дагестане сроки отрождения личинок вредителя фенологически совпадают с окончанием цветения и началом образования стручков у семенников капусты - при достижении суммы эффективных температур 1113 °С.
B. coerulescens в условиях Южного Дагестана начинает откладывать яйца перед цветением (12.V – 15.V) и продолжает это вплоть до июня. Измерениями ширины головной капсулы показано, что личинка брюквенного барида в своём развитии претерпевает 4 возраста: 1-й возраст - 0,32-0,33 мм; 2-й возраст - 0,43-0,44 мм; 3-й возраст -0,61 мм; 4-й возраст-0,85-0,86 мм. Особи первых возрастов предпочитают питаться во внешней паренхиме кочерыги. Личинки 3-го возраста прогрызают отверстия перпендикулярно оси кочерыги и выводят из строя сосудистую систему корня. Личинки 4-го возраста не перегрызая эпидермиса, устраивают удлиненную колыбельку из огрызков ткани растения и экскрементов, где и окукливаются. В условиях южного Дагестана среднемноголетняя дата появления перезимовавших особей - 13.III ±9,8 дней; появление 50% перезимовавших жуков отмечается обычно 16 IV. ±7,7 дней. Начало откладки яиц приходится на 20. V ± 4, 1 дней; Средняя дата появления первых куколок - 9.VII ± 9,1 дней. Средняя дата появления массового количества куколок соответствует - 16. VII ±12 дней. Среднемноголетняя дата массового отрождения жуков нового поколения приходится на 27.VII ±10,5 дней. Средняя продолжительность стадии личинки при температуре воздуха 19-21°С колеблется от 35 до 45 дней. Оптимальные условия для развития куколки — температура выше 25-27°С, относительная влажность - 65%. Фаза куколки в этих условиях проходит в течение 10-13 дней. При понижении температуры продолжительность фазы куколки увеличивается до 23-27 дней. При температуре воздуха 12-13°С превращения куколок в имаго не происходит. Отрождение жуков новой генерации начинается с середины июля.
Значение энтомофагов. Наблюдениями, проведёнными в 2004-2005 гг., установлено, что одним из многочисленных компонентов агробиоценоза семенной капусты являются жужелицы. В отдельные годы они были способны ощутимо снизить численность фитофагов. Отмечено, что капустные агроценозы населяли в основном жужелицы 9 видов, из которых доминантным являлся Poecilus cupreus Linnaeus (54% особей всех видов), а субдоминантными — Brachinus crepitans Daft. (13,6 %) и Clivina spaliatus Linn. (13% общей численности). Количество Amara aenaea De Geer., составляло 9,5%. Остальные виды (Anchomenus dorsalis Pontop., Laemostenus caspius Fald., Harpalus distinquendus Daft., Brachinus explodens Daft.) - встречались значительно реже (1,9-1,1% от общей численности). Указанные виды присутствовали на полях семенников белокочанной капусты практически с ранней весны и до поздней осени, но численность их в течение вегетации культуры значительно колебалась. В 1-й декаде мая плотность насекомых составляла 1,5 экземпляра на ловушку в сутки, в 3-й — 2,2 экземпляра, во второй и третьей декадах июня — 4,2 и 5,3 экземпляра на ловушку в сутки соответственно. Увеличение плотности хищных жужелиц, было связано с увеличением габитуса растений, которые к моменту образования стручков формировали довольно густую крону. Повышенная влажность и умеренный микроклимат в приземном слое в этот период являются благоприятными для развития жужелиц. Значительная разница в плотности жужелиц отмечена на полях, где предшественником была озимая пшеница (1,7±0,8 экземпляра на ловушку в сутки). По пласту многолетних бобовых трав (люцерна 2-го года) плотность жужелиц была в 3 раза выше (5,3±0,4 экземпляра на ловушку в сутки). Экспериментально установлено, что численность личинок брюквенного барида в севообороте с озимой пшеницей была достоверно выше, чем после люцерны 2-го года: 13,1±0,9 экз. на растение и 3,3±0,5 экз. на растение соответственно. Таким образом, насыщение овощного севооборота люцерной позволит в последующем ограничивать численность брюквенного барида.
Оценка сортов и родительских линий F1 гибридов семенников белокочанной капусты на устойчивость к брюквенному бариду. Для разработки интегрированной защиты семенников белокочанной капусты от брюквенного барида большое значение имеют сведения об устойчивости сортов к фитофагу. Исследования по изучению фенологических показателей ряда сортов и родительских линий F1 гибридов белокочанной капусты, которые по нашему мнению могли быть диагностическими при определении устойчивости к вредителю, проводились в 2000 - 2002 гг. Объектами исследований служили родительские линии следующих гибридов: Трансфер, Казачок, Малахит, Соло, Экстра, Крюмон, Монарх и др. Наблюдения показали разную степень устойчивости родительских линий F1 гибридов к брюквенному бариду. Устойчивость некоторых гибридов тесно связана с периодом одревеснения корневой шейки (если этот процесс полностью заканчивался ко времени массового отрождения личинок, то гибрид слабо заселялся вредителем). В зависимости от продолжительности вегетационного периода семенников белокочанной капусты посев раннеспелых сортов и гибридов в южной зоне Дагестана проводят с 15 по 25 августа; среднеспелых с 25 июля по 5 августа и позднеспелых с 22 по 28 июля. Высадку в грунт ранних сортов производят с 25 сентября по 5 октября; среднеспелых - с 25 августа по 15 сентября, позднеспелых - с 20 августа по 5 сентября. Нами было отмечено, что общее количество поврежденных растений возрастало при более ранних сроках сева, т.е. у позднеспелых родительских линий F1 гибридов (Экстра, Крюмон, Монарх и др.). Это связано с растянутостью периода бутонизации и цветения, что благоприятно сказывалось на откладке яиц и питании личинок брюквенного барида. Потери урожая семян у этих сортов были ежегодно высоки и составляли 45,5 % и более. Сорта раннеспелых сортов и гибридов (Трансфер, Экспресс, Казачок и др.), повреждались личинками брюквенного барида слабее. Потери урожая семян обычно не превышали 10,8-15,0% (см. табл. 2).
Таблица 2. Повреждаемость родительских линий F1 гибридов семенников белокочанной капусты личинками брюквенного барида. Дербентский район, 2000-2002 гг.
| Сорт, гибрид | Срок сева | Агроценоз | 2000 | 2001 | 2002 | |||
| 1 | 2 | 1 | 2 | 1 | 2 | |||
| Соло | 15.VIII | ДСОСВиО | ------ | ------ | 35±0,7 | 13,5±0,4 | ------ | ------ |
| Казачок | 18.VIII | а/ф им. Г.Давыдовой | 17±0,3 | 9±0,5 | 12±0,4 | 7,6±0,8 | 13,8±0,5 | 4,6±0,5 |
| Трансфер | 20.VIII | СПК им. Г. Казимова | 11±0,6 | 7,4±0,4 | 15±0,12 | 9,4±0,9 | ------ | ----- |
| Малахит | 22.VIII | а/ф «Музаим» | 16±0,2 | 9,6±0,8 | 15±0,15 | 8,2±0,3 | 12±0,4 | 6,7±0,3 |
| Экспресс | 25.VIII | СПК им. Г.Казимова | 10±0,1 | 8±0,4 | 28±0,34 | 11,2±0,45 | 12±0,5 | 7±0,45 |
| Монарх | 22.VII | ДСОСВиО | 55±0,5 | 6,5±0,6 | 60±0,45 | 12,7±0,35 | 55±0,3 | 12,2±0,6 |
| Экстра | 24.VII | ДСОСВиО | 85±0,3 | 27±0,8 | 100±0,4 | 28,6±0,85 | 75±0,7 | 26,4±0,1 |
| Колобок | 25 VII | ДСОСВиО | 65±0,4 | 7,7±0,4 | 60±0,35 | 16,3±0,78 | 55±0,2 | 18±0,2 |
| Крюмон | 28.VII | а/ф «Падар» | 100±0,3 | 32,5±0,5 | 90±0,7 | 29,3±0,3 | 85±0,3 | 27,3±0,8 |
| НСР05 | 2,4 | 2,01 | 2,6 | 2,4 | 2,5 | 2,02 | ||
Примечание: графа 1 - % заселенных растений в поле; графа 2 - численность личинок вредителя на 1 растение (экз. на растение).
Таким образом, в результате проведенных исследований достоверно выявляется фенологический тип устойчивости у раннеспелых сортов и родительских линий F1 гибридов белокочанной капусты, которые на протяжении исследуемых лет повреждались от слабой до средней степени.
Вредоносность и экономический порог вредоносности брюквенного барида на семенниках капусты
В связи с особенностями технологии выращивания семенников капусты, рекомендованные для продовольственной капусты экономические пороги вредоносности малоприемлемы при назначении химических обработок. Для таких вредителей, как капустная тля, рапсовый цветоед и брюквенный барид они не разработаны. В этой связи предусматривалось решение задач по разработке порогов вредоносности основных видов вредителей семенников капусты в условиях южного Дагестана.
Для расчета экономического порога вредоносности брюквенного барида была использована методика определения ЭПВ на основе оценки истребительных мероприятий. Было установлено, что при слабом повреждении растений (2-3 личинки на поврежденное растение) потери массы семян с 1 растения составляли - 6±1,1г, при среднем (6-8 личинок на 1 растение) - 12,8±1,2г, при сильном повреждении (более 13 личинок) - 32,2±1,3 г с одного растения, когда урожай семян снижался почти вдвое. Для обоснованности ЭПВ нам потребовалось конкретизировать этот показатель применительно к производственным условиям с учетом цен на продукцию и пестициды, затрат на защитные мероприятия и уборку сохраненного урожая, а также накладных расходов. На основе опытных данных было установлено, что снижение урожая в пересчете на 1 особь вредителя составляло 0,0202%. Снижение урожая рассчитывали по формуле В.И. Танского (1988):
З х Н х Р Зу
ЭПВ = ------------------ х (1 + --------), (1)
У х Ц х П х К Ц
где ЭПВ – экономический порог вредоносности (число особей на растение); З – затраты на защиту растений, руб./га; Зу – затраты на уборку сохраненного урожая, руб./ц; Н – накладные расходы, (%); Р – рентабельность, (%); У - планируемый (фактический) урожай, ц/га; Ц – закупочная цена продукции, руб./ц; К – биологическая эффективность, %; П – снижение урожая в пересчете на 1 особь вредителя, %.
На основе опытных данных установили: 3 - 12500 руб./га; Зу - 455 руб./ц; Н - 25%; Р -115,6%; У – 6,27 ц/га; Ц - 560 руб./кг; К-65%; П - 0,0202%. Подставляя полученные значения в формулу расчета ЭПВ, получили - 58361,1; Полученную цифру делили на среднюю численность семенников капусты на 1 га и получили 2,16 экз. на 1 растение. При средней плодовитости самок брюквенного барида – 80 яиц и соотношении полов 1:1 ЭПВ вредителя будет соответственно 0,05 экз. жуков на 1 растение.
Концептуальная модель прогноза численности популяции брюквенного барида в условиях Дагестана
Важнейшим требованием к химическим обработкам против вредителей является их своевременность. Но, к сожалению, из-за недостатков прогнозирования оно выполняется далеко не всегда. Основой предлагаемой модели может служить фенологическая сопряженность развития брюквенного барида и растения-хозяина (семенной капусты). Для прогнозирования результата необходимо знать экспериментальные значения всех входящих в модель факторов. Наиболее существенными для построения модели были: ГТК (по Селянинову); сумма эффективных температур ( эфф. t°); сумма активных температур ( акт t°); смертность преимагинальных стадий вредителя (данные таблиц выживания). Каждый показатель условно делили на три уровня: годы с высокой плотностью вредителя - ГТК-1 (0,15-0,20); средней плотностью вредителя ГТК – 2 (0,3-0,6) и показатель ГТК <0,6, когда вредитель отмечается незначительно. Таблицы выживания: 10-15% -я; 30-40% - я и 70-80% - я выживаемость. Анализ корреляций между сопряжёнными факторами позволил нам выявить основной фактор модели (ГТК), который с одной стороны, показывает высокую отрицательную корреляцию между акт t°, эфф. t° в градации А (100% заселённость растений) (r = - 0,94) и тесную положительную связь между теми же показателями в градации Б (60% заселенность растений) (r = 1). Результативный признак численности популяции мы обозначили через h, а два других факторных признака соответственно через X1, и х2.
Однофакторная полиномиальная модель зависимости численности популяции от величины ГТК выражается функцией: h=a+в X1+СX12.
Таблица 3. Зависимость численности популяции от величины ГТК по годам исследований
| Год | А | ГТК |
| 1998 | 0,9 | 0,15 |
| 2000 | 0,86 | 0,31 |
| 2001 | 0,625 | 0,32 |
| 2003 | 0,625 | 0,3 |
| 2005 | 0,687 | 0,3 |
Используя данные таблицы, решив систему уравнений, получаем модель
h=0,031x12-0,252x1+l, 158 0,15 <=х1<=0,3,h >0,6
Для оценки влияния фактора X1=ГTK на численность популяции вычисляется коэффициент эластичности, который характеризует относительное изменение результата при изменении ГТК на 1%. 3 - (B+2cх1) (х1/h), где х1, h - средние значения. В нашем случае Э = - 0,08766. Это означает, что при увеличении ГТК на 1%, численность популяции уменьшается на 8%. Двухфакторная модель зависимости численности популяции от ГТК (х1) и Y(х2) имеет вид: h=0,029 х12-0,2 х1+l,l6+0,01х2, совокупный коэффициент детерминации R=0,89. На основании выводов модели, а также изучения корреляционных зависимостей численности популяции брюквенного барида, нами для практических целей создана логическая модель, позволяющая прогнозировать плотность популяции вредителя на следующий год.
Таблица 4. Логическая модель прогноза развития брюквенного барида
| Градации численности популяции | Минимально - достаточные данные для прогноза |
| h1 (численность популяции при ГТК1) | ГТК1, (от 0,15 до 0,3);Y, или Y2 (от 10% до 40%). |
| h2 (численность популяции при ГТК2) | ГТК2 (от 0,5 до 0,6); Y1 (от 10% до 15%); X - проведенная защита (1 или 2- кратно) |
Таким образом, для прогноза распространения вредителя достаточными являются гидротермический коэффициент, градации выживания популяции за осенне-зимний период (альтернативно - смертность) и проведенные в предыдущем году химические обработки. Последний фактор учитывается в том случае, если не соблюден севооборот и семенная капуста высаживается на прежних участках.
4.2. Биоэкологические особенности развития капустной тли (Brevicoryne brassicae L.) на семенниках белокочанной капусты
Особенности расселения капустной тли и закономерности её пространственного распределения на полях позволяют предвидеть массовое размножение вредителя, что в свою очередь даёт возможность сделать своевременный и грамотный прогноз. Для правильного учета численности фитофага и установления оптимальных сроков борьбы с ним необходимы сведения о «миграционных пиках». Мы изучали динамику численности популяции капустной тли, регистрируя возрастную структуру и миграционные процессы. Исследованиями установлено, что интенсивность лёта насекомых значительно менялась в течение сезона вегетации культуры. В весенний период количество отлавливаемых в ловушки насекомых незначительно (в среднем 27,6 экз. на ловушку в сутки). В это время в популяции капустной тли преобладают личинки младших возрастов (до 70%); во второй половине вегетации семенников капусты пропорция возрастных групп была примерно одинаковой. В колониях капустной тли помимо девственных самок, развиваются крылатые самки - расселительницы, активно перелетающие на соседние агроценозы. В июне средняя численность отловленных в ловушках насекомых составила 56 экз., в июле – 129,7. Из-за отсутствия факторов, сдерживающих численность фитофага, наблюдается значительное увеличение размеров популяции. Наиболее высокая численность отмечали в сентябре, когда за сутки отлавливалось до 230 особей тли на 1 ловушку в сутки. Осенью в колониях вредителя появляются самки - полоноски, отрождающие личинок, из которых развиваются и самки и самцы. Мигрируя, они также способствуют концентрации вредителя в новых очагах. Популяция мигрирующих особей сохранялась относительно высокой вплоть до декабря.
Результаты исследований свидетельствуют о выраженной тенденции к росту популяции в течение сезона и больших потенциальных возможностях вредителя.
Изменение плотности популяции капустной тли на семенниках капусты в течение года и уровень паразитизма её афидофагами. Агроценоз семенников капусты обладает достаточно высокими резервными возможностями саморегуляции, обеспечивающими предотвращение возникновения вспышек размножения капустной тли. Капустная тля на плантациях семенников белокочанной капусты ассоциирована с 17 видами многоядных и специализированных энтомофагов. Наиболее активную регулирующую роль в сокращении популяции капустной тли проявляют златоглазки Chrysopa carnea Stephens и C. formosa Brauer, а также наездник-афидофаг Diaeretiella rapae (M'Intosh) сем. Aphidiidае. В отличие от хризопид для имаго D. rapae характерна способность быстро находить и откладывать яйца на растения, заселенные единичными тлями. Нами в течение ряда лет прослеживалось влияние D. rapae на динамику численности капустной тли. По итогам многолетнего мониторинга мы пришли к выводу, что в отдельные годы энтомофаг способен эффективно регулировать численность капустной тли. Появление мумифицированных особей тли на семенных растениях капусты отмечается сразу после образования небольших колоний вредителя в начале марта, но в силу незначительной его численности в этот период афидофаг не способен оказывать ограничивающего воздействия на популяцию вредителя. Это воздействие начинает проявляться только со второй половины июля, когда соотношение хищник – жертва достигает 1:40. Именно при таком соотношении комплекс природных афидофагов, в котором доминирует D. rapae, способен самостоятельно сдерживать численность тлей. Таким образом, природные механизмы, ограничивающие размножение капустной тли, включаются в действие, как правило, во второй половине лета. К этому времени вредитель успевает достичь высокой численности и причинить значительный ущерб урожаю капусты. Процент паразитированных особей в этот период бывает незначительным (22,3 - 24,7%). Максимум численности D. rapae наблюдался с июля по сентябрь (70-80 % паразитированных особей). В 2005-2007гг. гидротермические условия были благоприятны для развития вредителя. Экспериментально установлено, что D. rapae и в полевых, и в контролируемых лабораторных условиях отдавала предпочтение личинкам второго (32,8%) и третьего (43,2 %) возрастов капустной тли, чем личинкам первого возраста и взрослым особям. Среднее количество личинок на одну паразитированную самку составило 8,6, а на непаразитированную - до 32 экземпляров. Таким образом, полученные данные свидетельствуют о том, что в отдельные годы соотношение хищник жертва и синхронизация их развития в значительной степени могут влиять на вредоносность капустной тли.
Экономический порог вредоносности капустной тли. ЭПВ капустной тли определяли по методике В.И. Танского (1988). Для получения дифференцированной степени заселения семенников капусты вредителем, проводили варьированное опрыскивание растений инсектицидами. Варианты были сгруппированы следующим образом: 1 балл - отдельные колонии на 15% поверхности листьев; 2 балла - 25% поверхности листьев и побегов; 3 балла- до 50% поверхности растений; 4 - контроль (без тлей). Урожай семян собирали в фазе восковой спелости отдельно по вариантам. В результате было установлено, что при повреждении растений по 1- му баллу, урожай семян почти не снижался. При повреждении растений по 2 – му баллу урожайность семян капусты снижалась на 18%, по 3 - му баллу - почти вдвое, т.е. средняя масса семян с неповрежденных растений составляла в среднем 55 г; с поврежденных по 1-му баллу - 54 г, по 2-му — 33 г, по 3 - му- 28 г семян с 1 растения. Подставляя полученные значения в расчётную формулу:
Х = Р2 х Б2+ Рз х Бз /М, (2)
(где X - общие потери (%); Р2, Рз - число растений с повреждениями 2 и 3 балла во всех учетах; Б2, Бз - потеря урожая (%), при повреждении с 2-мя и 3-мя баллами; М- общее число обследуемых растений), мы получили: 15,7% общих потерь. Таким образом, ЭПВ 5% капустной тли соответствовал - 25% заселенных растений семенной капусты (по 2-му баллу). Данный критерий достаточно легко использовать при полевой оценке численности вредителя.
4.3. Биоэкологические особенности развития рапсового цветоеда (Meligethes aeneus Fab). на семенниках белокочанной капусты
Результаты фитосанитарного мониторинга, проведённого на семенниках капусты за ряд последовательных лет, показали, что в 1998 и 2003 гг. отмечался подъём численности вредителя. Заселенность растений рапсовым цветоедом в эти годы составляла 50-75%, при средней численности 7,9 экз. вредителя на растение. Менее благоприятными для развития фитофага были 2006-2007 годы. Плотность популяции вредителя в этот период была ниже ЭПВ и составляла 0,2-0,6 экз. на растение при 5 - 7% -ой заселённости (см. рис.4).

Рис. 4. Многолетняя сезонная динамика численности рапсового цветоеда на семенниках капусты, ДСОСВ и О, Дербентский район Дагестана.
Мы связываем это с низкими температурами в период выхода жуков с мест зимовки. Так, среднесуточная температура воздуха в апреле 2007 г. составляла 9,7° С (нижний порог развития вредителя – 9,4°С). В 2006 году этот показатель составлял 12,2 °С. Для планирования защитных мероприятий необходимо знать сроки начала выхода популяции вредителя с мест зимовки. В условиях Дербентского района рапсовый цветоед появляется на полях семенной капусты при среднедекадной температуре воздуха -10,8°С. В эти дни максимальные дневные температуры обычно достигали 12-17°С. Массовое появление жуков рапсового цветоеда мы регистрировали в середине 2-й декады апреля, при достижении эфф. t° - 24°С ±10,1°С. Как правило, откладка яиц происходит через 3-5 дней после выхода жуков. Массовая откладка яиц осуществлялась в 3-й декаде апреля - при эфф. t° - 47,9°С. Среднемноголетняя дата начала отрождения личинок рапсового цветоеда - 26 IV. ± 5,8 дней, что обычно наблюдается при достижении эфф. t° - 67,5°С. Куколки рапсового цветоеда развивались в основном во 2-й и 3-й декадах мая. Средняя дата появления первых куколок по наблюдениям в исследуемые сезоны соответствовала 4.V ± 5,3 суток. Средняя дата появления массы куколок соответствовала – 15.V ± 5,4 суток. Приведенные нами многолетние показатели сезонного развития рапсового цветоеда могут служить ориентиром для практических целей, т.к. с их помощью можно прогнозировать наступление отдельных фенологических фаз развития вредителя, а также планировать сроки проведения обработок.
Вредоносность и экономический порог вредоносности рапсового цветоеда на семенниках капусты
Для определения критериев ЭПВ рапсового цветоеда нами были заложены эксперименты на базе агрофирмы им. Г. Давыдовой Дербентского района в 2004-2005 гг. Порог вредоносности рапсового цветоеда также определяли по методике Танского В.И. (1988). В зависимости от плотности популяции вредителя в первую очередь снижались такие показатели, как число ветвей первого порядка, число стручков на растении, число семян в стручке и масса семян с растения. Последовательное увеличение количества жуков с 3 до 20 приводило к снижению массы семян с одного растения по сравнению с контролем на 2,4; 4,0; 8,3; 15,4 и 19,8 г соответственно. По нашим данным, потери урожая семян семенной капусты от рапсового цветоеда довольно значительны. Максимальная разница в урожае семян составила 37,6%. Используя данные опыта мы получили: 3 - 435 руб./га, Зу -115 руб. 80 коп, Н - 25% = 1,0 (коэффициент), Р -124,53% = 2,27, У - 5,5 ц/га, Ц - 5600 руб./ц, К- 95,8% = 0,99, П - 0,0402 %.
Таким образом, двукратное опрыскивание семенников капусты Арриво, КЭ (250 г/л) с биологической эффективностью 95,8%, целесообразно при достижении рапсовым цветоедом плотности 2 экземпляра на растение в фазу бутонизации.
- Биоэкологические особенности развития крестоцветных блошек (род Phyllotreta) на семенниках белокочанной капусты
Сопряженность критических фенологических фаз семенников капусты и крестоцветных блошек обусловливает различный характер их вредоносности. В условиях Дербентского района развитие вредителя характеризуются двумя пиками. Первый соответствует фазе молочно-восковой спелости семян (массовое формирование стручков), второй приурочен к наиболее уязвимой фазе всходов — две пары настоящих листьев. Наибольшая вредоносность крестоцветных блошек отмечается с 20. VII по 10.VIII.
Данные многолетнего фитосанитарного мониторинга семенников капусты за ряд последовательных лет в агроценозе агрофирмы «Штул» свидетельствуют о периодичности вспышек распространения вредителя, происходящих раз в 2-3 года (см. рис. 5). Вредитель имеет довольно широкий круг трофических связей в ареале распространения. Высокую плотность крестоцветных блошек в годы исследований отмечали на Barbarea vulgaris, Sinapis arvensis, Raphanus raphanistrum, Capsela bursa pastoris и др. в начале марта, при среднесуточной температуре воздуха 6-7°С. Численность блошек в этот период достигала 150 экз. на кв. м. Наибольшее количество крестоцветных блошек в годы исследований мы находили на Barbarea vulgaris (сурепка обыкновенная) и Raphanus raphanistrum (редька дикая) – до 86,2% всех отмеченных поврежденных растений.

Рис. 5. Многолетняя сезонная динамика численности крестоцветных блошек на семенниках капусты, агрофирма «Штул», Дербентский район Дагестана.
4.5. Биоэкологические особенности развития капустной моли (Plutella maculipennis Curt.) на семенниках белокочанной капусты
Капустная моль в условиях Дербентского района имеет до 5 поколений в год. Бабочки 1-го поколения появляются, при достижении среднесуточных температур 15-17°С - 9.IV ±6,1 дней. Спустя 3-4 дня самки начинают откладывать яйца. Средняя дата откладки массы яиц бабочками первого поколения приходится на 25.IV±3.4 дней. Потенциальная плодовитость одной самки составляет 80-100 яиц, период яйцекладки обычно длится 20-25 дней. Экспериментально установлено, что эфф. t° для развития яиц составляет 64,4 -77,7°С, для развития гусениц - 108,0 - 117,7°С, для формирования куколок - 124-144°С. эфф. t°, необходимых для развития одного поколения составляет 369,1-395,9°С. Продолжительность эмбрионального развития по нашим наблюдениям составляла в среднем 3-5 дней. Среднемноголетняя дата отрождения гусениц в 1- м поколении - 21. IV ±7.6 дней; во 2-м - 4.VI ±5,2 дней; в 3-м – 26.VII ± 8,6 дней. Среднемноголетняя дата начала окукливания в 1- м поколении – 21. V ±12 дней, массового – 27.V ±8,7 дней; во 2-м и 3-м соответственно -19.VI ±11,6 дней; 26.VI ±4,8 дней и 3.VIII ±10,7 дней; 23 VIII ±7,7 дней.
Лёт бабочек в 4-м поколении обычно отмечается с 5. IX по 12.IX; массовое отрождение гусениц с 17.IX по 22.IX; начало окукливания 25.IX; массовое окукливание происходит с 10. X по 25.X.
Динамика развития популяций капустной моли в южной зоне Дагестана характеризуется определенным постоянством: наличием вспышек, достигающих эпизоотического масштаба с периодичностью 1-2 раза в 10 лет. Последняя из них отмечалась в 2001-2002 гг., когда на каждое заселенное растение приходилось более 30 разновозрастных личинок капустной моли, при 70-85% заселенности растений (см. рис. 6).

Рис. 6. Многолетняя сезонная динамика численности капустной моли на семенниках капусты, ДСОСВ и О (Ряд 1- среднее количество экз. вредителя на заселённое растение, ряд 2- заселенность растений, %).
Многолетними наблюдениями за динамикой развития капустной моли установлено, что в регулировании размножения вредителя существенную роль играют местные виды энтомофагов, в частности: Diadegma collaris Grav, D. semillausum Hell, D. fenestralis Holm, относящиеся к семейству ихневмонид (Ichneumonidae). Данные виды были идентифицированы систематиком А.И. Халаим (ЗИН РАН). Указанные виды паразитоидов заражают более 60% особей вредителя, как в фазе личинки, так и в фазе куколки, благодаря чему заметно сдерживают размножение фитофага. В отдельные годы, при невысокой численности фитофага даже 25-30%-й уровень паразитирования оказывался достаточным для полного подавления популяции капустной моли. Нами отмечено, что цикл развития паразитоида точно сопряжён с циклом развития хозяина. Средняя продолжительность полного цикла развития капустной моли и паразитоида диадегмы составляет 25-28 дней. На основе мониторинга и экспертно-диагностической оценки плантаций семенников белокочанной капусты на заселенность капустной молью, ЭПВ и прогноза вредоносности, в 2007 году были отменены химические обработки на площади 12 га в агрофирме «Штул» Дербентского района, что позволило хозяйству сэкономить 40764 руб. Акты об отмене химических обработок на семенниках капусты приведены в приложениях диссертации.
Глава 5. Популяционная экология вредителей семенников белокочанной капусты в Дагестане
Для построения тактики и стратегии защиты семенников белокочанной капусты от вредителей важное значение имеют сведения об изменении численности их популяций. Для изучения баланса популяций мы воспользовались таблицами выживания насекомых, которые дают возможность оценить сравнительное значение каждого фактора смертности в отдельности и выделить критические периоды в течение генерации, определяющие дальнейший ход динамики численности. Результат высокой смертности брюквенного барида вытекает из нескольких причин: гибели при отрождении; гибели от хищников; энтомофторозов; теплового шока (причиняемого растениям при сборе урожая, тем, что отсекается крона семенного растения), гибели во время перезимовки и др. В фазе яйца гибель насекомых происходит вследствие стерильности (средний показатель смертности за три года 2,5% погибших особей). Смертность личинок может быть вызвана рядом причин, среди которых наибольшее значение имеет гибель от хищников и гибель при отрождении (остаточная смертность - 0,01 и 0,02 соответственно) (табл. 5).
Таблица 5. Таблица выживания популяции брюквенного барида на семенниках капусты. Дербентский район, ДСОСВиО, 2005г.
| Стадия развития | Факторы смертности | Процент погибших особей | Количество погибших особей | Количество живых особей | lg числа живых | Остаточная смертность (уровень k) |
| Яйцо | Реальное число | 665 | 2,82 | |||
| В т.ч.стерильных | 2,52 | 16,7 | 648,3 | 2,81 | 0,01 | |
| (K яиц =0,01) | ||||||
| Личинка | Гибель при отрождении отрождении | 4,1 | 26,6 | 621,7 | 2,79 | 0,02 |
| Хищники | 4,7 | 29,2 | 592,5 | 2,77 | 0,02 | |
| Болезни | 3,7 | 21,9 | 570,6 | 2,74 | 0,03 | |
| Тепловой шок | 3,6 | 20,5 | 550,1 | 2,74 | - | |
| (K лич. = 0,07) | ||||||
| Куколка | Болезни Хищники | 2,8 | 15,4 29,9 | 534,7 | 2,73 | 0,01 |
| Хищники | 5,6 | 504,8 | 2,70 | 0,03 | ||
| Тепловой шок | 9,45 | 47,7 | 457,1 | 2,66 | 0,04 | |
| (K кук. =0,08) | ||||||
| (K в период преимагинального развития = 0,16) | ||||||
| Имаго | Хищники | 8,2 | 37,5 | 419,6 | 2,62 | |
| Болезни | 6,5 | 27,3 | 392,3 | 2,59 | 0,03 | |
| Гибель во время перезимовки перезимовки | 12,3 | 48,2 | 344,1 | 2,54 | 0,05 | |
| Общая гибель и миграции | 63,45 |
Жертвами хищников обычно бывают личинки младших возрастов, пока те еще питаются внешней паренхимной тканью растения-хозяина. Определенная часть личинок гибла от болезней и вследствие стресса, который мы условно назвали тепловым шоком, вызванным при срезании верхней кроны растения при уборке. Оставленные после уборки кочерыги пересыхают, вызывая массовую гибель неокуклившейся части личинок. Гибель в результате теплового шока отмечена у 3,6% особей (рис. 7).
Рис. 7. Факторы смертности популяций брюквенного барида на плантациях семенников капусты ДСОСВ и О, 2005-2007гг. (1- cтерильность, 2- гибель при отрождении, 3- хищники, 4- болезни, 5- тепловой шок, 6- гибель во время перезимовки)
Более высокий процент гибели от этих факторов был отмечен в фазе куколки. Наиболее высок процент гибели куколок от усыхания вследствие теплового шока – 9,45% (уровень k =0,04). В 5,6 % смертность вредителя была вызвана деятельностью хищных жужелиц. Гибель вредителя в имагинальной стадии также происходила от этих факторов. В 8,2% случаев гибель происходила от хищников, однако основная гибель фиксировалась в период перезимовки.
Таким образом, проведённый К-факторный анализ популяций брюквенного барида свидетельствует об их высокой выживаемости, в среднем за три года равной 46,55%. Наиболее уязвимой возрастной стадией развития являлась фаза куколки. Высокая смертность в этой возрастной стадии происходила в основном от действия двух наиболее значимых факторов: гибели в результате теплового шока и от хищников.
Анализ таблиц выживания рапсового цветоеда по итогам наблюдений за 2005-2007гг. показал, что основным фактором смертности вредителя в стадии яйца является стерильность. Смертность личинок вызвана в основном действием хищников и паразитоидов. В фазе куколки смертность особей в основном происходила от болезней (6,4%) и паразитоидов (4,3%). Гибель во время перезимовки, а также исчезновение в период миграционных перемещений, зарегистрированные весною 2006 г., составили 42%. Общая гибель популяции рапсового цветоеда в период преимагинального развития составила 32,9% (остаточная смертность 0,169). Результаты наблюдений в 2005-06 гг. приведены в таблице 6.
Таблица 6. Таблица выживания популяции рапсового цветоеда на семенниках капусты. Дербентский район, ДСОСВ и О, 2005-06 гг.
| Стадия развития | Факторы смертности | Процент погибших особей | Число погибших особей | Число живых особей | lg числа живых особей | Остаточная смертность (уровень k) |
| Яйцо | Реальное число на 20 погонных м ряда | 560 | 2,748 | |||
| Стерильность | 2,0 | 11,2 | 548,8 | 2,739 | 0,009 | |
| Личинка | Гибель при отрождении | 2,2 | 12,32 | 536,5 | 2,73 | 0,009 |
| Каннибализм и хищники | 9,5 | 53,2 | 483,3 | 2,68 | 0,05 | |
| Болезни | 1,8 | 10,1 | 473,2 | 2,67 | 0,01 | |
| Паразитоды | 4,6 | 25,7 | 447,5 | 2,65 | 0,02 | |
| Неизвестные факторы | 2,1 | 11,7 | 435,8 | 2,64 | 0,01 | |
| Куколка | Болезни | 6,4 | 35,8 | 400 | 2,60 | 0,04 |
| Паразитоиды | 4,3 | 24,1 | 375,9 | 2,57 | 0,03 | |
| Общая гибель в период преимагинального развития | Все факторы | 32,9 | (К в период преимагинального развития = 0,169) | |||
| Имаго: перезимовавшие жуки (число на 20 пог. м. ряда) | Гибель во время перезимовки и миграционное перераспределение | 42 | 157,8 | 218,1 | 2,3 | 0,27 |
Лимитирующим фактором в фазе личинки в 2006 г. были каннибализм и хищники (остаточная смертность 0,04). Примерно одинаковой была гибель от болезней и паразитоидов (2,7 и 2,4%, остаточная смертность 0,02). В 2,6% случаев причину гибели личинок определить не удалось (смертность от неизвестных факторов 0,01). В фазе куколки в 2006 г. Лимитирующим фактором были болезни (остаточная смертность 0,02). В 2,1% случаев смертность была вызвана деятельностью паразитоидов (k =0,01). Гибель во время перезимовки, а также исчезновение в период миграционных перемещений, зарегистрированные весною 2007 г., составили 37%. Общая выживаемость популяции рапсового цветоеда за генерацию составила 43,3 % (116,9 выживших жуков по отношению к начальному числу яиц, равному 270) (см. табл.7).
Таблица 7. Таблица выживания популяции рапсового цветоеда на семенниках капусты в СПК им. Г.Казимова, Дербентский район, 2006-2007 гг.
| Стадия развития | Факторы смертности | Процент погибших особей | Число погибших особей | Число живых особей | lg числа живых особей | Остаточная смертность (уровень k) |
| Яйцо | Первоначальное число на 20 пог. м ряда | 270 | 2,43 | |||
| Стерильность | 2,2 | 5,9 | 264,1 | 2,42 | 0,01 | |
| Личинка | Гибель при отрождении | 3,7 | 10,0 | 254,1 | 2,40 | 0,02 |
| Каннибализм и хищники | 8,2 | 22,14 | 231,9 | 2,36 | 0,04 | |
| Болезни | 2,7 | 7,3 | 224,6 | 2,35 | 0,02 | |
| Паразитоиды | 2,4 | 6,5 | 218,1 | 2,33 | 0,02 | |
| Неизвестные факторы | 2,6 | 7,02 | 211,1 | 2,32 | 0,01 | |
| Куколка | Болезни | 6 | 16,2 | 195 | 2,29 | 0,03 |
| Паразитоиды | 2,1 | 5,7 | 189,3 | 2,28 | 0,01 | |
| Неизвестные факторы | 1,4 | 3,8 | 185,5 | 2,27 | 0,01 | |
| Общая гибель в период преимаг. развития | Все факторы | 31,3 | (К в период преимагинального развития =0,17) | |||
| Имаго: перезимовавшие жуки (число на 20 пог. м. ряда) | Гибель во время перезимовки и миграционное перераспределение | 37 | 68,6 | 116,9 | 2,06 | 0,21 |
К - факторный анализ выживаемости популяции капустной моли на семенниках капусты в условиях южной зоны Дагестана показал, что наиболее уязвимой возрастной стадией развития вредителя по результатам трёхлетних наблюдений является фаза куколки. Высокая смертность в этой фазе происходит в основном от действия паразитоидов: D. collaris, D. semillausum, D. fenestralis. Значение таких факторов, как хищники, болезни и др. существенной роли в выживаемости популяции капустной моли не играли. Гибель от паразитоидов была достаточно высокой и в последующие годы (см. рис.9).
Рис. 9. Факторы смертности популяций капустной моли на плантациях семенников капусты. ДСОСВ и О, 2005-2007гг. (1-стерильность; 2 – гибель при отрождении; 3 – хищники; 4 – паразитоиды; 5 – болезни)
Общая гибель особей преимагинальных стадий капустной моли в 2005 - м году составила 75,7%. В последующие годы смертность вредителя была немного ниже и составляла 70,2% в 2006 г. и 61,3% - в 2007г. Таким образом, эффективность паразитоидов из рода диадегма является достаточной чтобы сдерживать популяцию вредителя на хозяйственно-неощутимом уровне.
Результаты К-факторного анализа выживаемости популяции капустной тли показали, что в отдельные годы основным фактором, лимитирующим рост популяции вредителя, являются энтомофторозы. Значительная гибель от энотомофторозов отмечалась в 2006 году в возрастной стадии «личинки младших возрастов». Гибель от паразитоидов составляла 21,8% (остаточная смертность k = 0,11). Высокий процент паразитизма был отмечен также в возрастной стадии «бескрылые самки» (18,2%, остаточная смертность k= 0,09). Гибель 6% особей бескрылых самок была вызвана хищниками (Chrysopa formosa Brauer и C. carnea (Stephens)., а также личинки мух сирфид A. obliqua, A. venusa) (остаточная смертность от действия хищников k=0,02). Общая ежегодная смертность преимагинальных стадий вредителя по итогам трёх лет наблюдений составила 84,6% (см. рис. 10).
Рис. 10. Факторы смертности популяций капустной тли на семенниках капусты. ДСОСВ и О, 2004, 2006, 2007 гг. (1- гибель от неизвестных факторов; 2 – болезни; 3 – паразитоиды; 4 – хищники)
Оценивая уровни влияния регулирующих факторов на динамику развития популяций вредителей, можно констатировать, что выживаемость популяций брюквенного барида и рапсового цветоеда достаточно высока (36 - 58 %). Основным фактором смертности брюквенного барида являлась перезимовка. Высокая смертность на фазе куколки происходила от действия двух наиболее значимых факторов: гибели в результате теплового шока и фактора хищничества. У рапсового цветоеда основная гибель также происходила в период перезимовки. На втором месте стоят такие факторы, как каннибализм личинок и хищники. Относительно низкая смертность популяции в период преимагинального развития обусловлена, прежде всего, защищенностью особей от внешних факторов среды, т.к. они проходят основные стадии развития внутри тканей растения-хозяина.
Анализ таблиц выживания капустной моли и капустной тли показал, что энтомофаги Diadegma collaris Grav, D. semillausum Hell., D. fenestralis Holm., Diaeretiella rapae (M'Intosh), Chrysopa formosa Brauer и C. carnea (Stephens) и др. в отдельные годы играли ключевую роль в регуляции численности фитофагов и были способны самостоятельно регулировать плотность популяций последних. Знание их сезонной динамики и закономерностей пространственного распределения на семенниках капусты позволили в перспективе оптимизировать существующие меры борьбы с ними и сделать достоверный прогноз распространения в следующем сезоне.
Глава 6. Разработка системы мониторинга фитосанитарного состояния семенников капусты в отношении их вредителей
Успех интегрированной защиты во многом зависит от принятой системы наблюдений за динамикой численности вредителей и их естественных врагов. В практической защите растений оперативные работы обычно организуются по указателям средних величин развития вредных организмов на заселенной площади, основными критериями которой являются количество поврежденных растений и площадь, заселенная вредным организмом (К.М. Степанов, А.Е. Чумаков (1972), И.Я. Поляков и др., (1984,1988). Однако численность вредителей на полях в разные годы значительно колеблется, поэтому использовать одни и те же объемы выборок недопустимо. В связи с этим, нами разработаны методы фитосанитарного мониторинга, позволяющие объективно оценить развитие и распространение вредителей семенников капусты. Они позволяют наиболее точно определять сроки проведения защитных мероприятий, осуществлять выборочные обработки инсектицидами, а при средней численности вредителей ограничиться биозащитой.
В 2004-2005гт. на полях трех хозяйств (МУП агрофирма им. Г. Давыдовой, а/ф «Штул» и ДОСВНИИР) мы изучали закономерности пространственного распределения капустной моли, рапсового цветоеда и брюквенного барида. Маршрут обследования выбирали с таким расчётом, чтобы ошибка в оценке значения средней численности вредителя не выходила за пределы 20-30%. Предварительно на поле просматривали 100 растений (проб) и определяли среднее число вредителей на 1 растение.
По результатам исследований были составлены графики, позволяющие определить объем выборки с заданной точностью. Графики строили по формулам отрицательного биноминального распределения, которому подчиняется распространение видов по полю. Они выражали зависимость объема выборки от численности вредителя при заданных уровнях точности, равных 10% (0,1), 20% (0,2) и 30% (0,3).
Для вредителей с повышенной численностью по краям полей (рапсовый цветоед, капустная тля) обследования проводили по Z - образному маршруту - одну половину вдоль двух краев поля, а вторую по одной из диагоналей. Многолетними исследованиями установлено, что заселённость фитофагами по краям полей в среднем в 3,4 - 4 раза выше, а максимально в 7,5 раз выше, чем в центре поля.
Для расчета поврежденности крестоцветными блошками мы воспользовались методикой Попова Ю.В. (ВНИИЗР) (1992г). Вначале установили площадь одного повреждения (0,25 мм, 0,5 мм, 1 мм и т.д.), затем приближенно подсчитали среднее число повреждений в малом квадрате, перемножили их число на площадь 1 повреждения (пятна), делили на 6,3 и выразили в процентах.
Рп = SnR/ 6,3 х 100 (3)
где Sn — площадь повреждения; R — среднее количество повреждений в малом квадрате. При отказе от химических обработок или снижении их объема маршрутными обследованиями было охвачено до 90-100% общей площади семенников капусты, в то время как при обследовании посевов, где систематически применялись пестициды - не более 10-15%. Более частые маршрутные обследования позволяют проводить мониторинг полезной и вредной биоты с высокой степенью точности.
Глава 7. Экологически обоснованная технология применения инсектицидов в программах защиты семенников капусты от основных вредителей
Несмотря на широкое применение испытуемых нами препаратов против комплекса вредителей различных сельскохозяйственных культур, на семенниках капусты они не зарегистрированы. Государственные каталоги пестицидов и агрохимикатов, разрешённых к прмименению на территории Российской Федерации, опубликованные в последние 10 лет, содержали лишь несколько наименований пестицидов, рекомендованных для семенных посевов овощных культур, в том числе капусты (Би- 58 Новый, КЭ (400г/л), Данадим, КЭ (400г/л), Фосфамид, КЭ (400 г/л), Арриво КЭ (250 г/л), Циткор КЭ (250 г/л). Кроме того, биологические особенности таких вредителей, как брюквенный барид, капустная тля и др. демонстрируют высокую сопряженность развития с фенологической фазой цветения у кормового растения. В этой связи нами проводилось широкое испытание новых для региона инсектицидов, а также инсектицидов, уже известных и широко применяемых в системах защиты других культур. При подборе средств защиты мы ориентировались на такие значимые критерии, как селективность, низкая норма расхода, препаративная форма, а также класс опасности. Опрыскивание семенников проводили в различные фазы вегетации растений в зависимости от сроков появления изучаемых вредителей. В результате испытаний были выделены наиболее эффективные препараты как селективного, так и широкого спектра действия.
Испытание инсектицидов против брюквенного барида и капустной тли. Результаты фенологического мониторинга показали, что капустная тля и брюквенный барид наиболее вредоносны в период цветения и начала образования стручков. В практической защите против указанных вредителей широко использовали препараты из группы пиретроидов и ФОС. К указанной группе инсектицидов в последние годы отмечалась групповая резистентность насекомых. В этой связи, нами проводилось широкое испытание альтернативных по химическому составу препаратов из группы никотиноидов (Регент, КЭ (25 г/л), Моспилан, РП (200 г/кг) и неоникотиноидов (Актара, ВДГ (250 г/кг), Конфидор, ВРК (200г/л). Благодаря своим трансламинарным свойствам, эти препараты быстро поглощаются и перемещаются через эпидермис листа. Эталоном служил Би - 58 Новый, КЭ (400 г/л) в норме 0,6, 1 и 1,5 л/га. Проведённые исследования показали, что наиболее эффективными в борьбе с брюквенным баридом и капустной тлёй оказались Конфидор, ВРК (200г/л) и Моспилан, РП (200 г/кг). При обработке Моспиланом, РП (200 г/кг) значительное количество особей тли погибало уже через сутки, полная гибель наступала через 3 суток. Конфидор, ВРК (200г/л) проявил выраженное системное действие. Уже через 3 суток гибель насекомых составила 98,7%. Полная их гибель отмечалась на 5-е сутки. Хорошую эффективность показал также инсектицид Регент, ВДГ (250 г/кг). Этот препарат на 5-е сутки оказывал такое же действие как Моспиланом, РП (200 г/кг). Что касается эталонного препарата БИ - 58 Новый, КЭ (400г/л), то следует отметить, что при рекомендованных нормах расхода (0,6 л/га) его эффективность на 3-и сутки не превышала 84,5%. Однако, при норме расхода препарата 1л/га гибель насекомых составляла 98,7%, а при 1,5 л/га – 99,5%. Полная гибель капустной тли наступала на 5-е сутки. Таким образом, для получения 95-100%-й эффективности необходимы, соответственно, более высокие нормы расхода этого препарата.
Испытания ряда биопрепаратов показали, что популяции капустной тли оказались менее чувствительными к действию биопрепаратов по сравнению с приведёнными инсектицидами. Смертность вредителя при обработке растений Битоксибациллином, П (БА – 1500 ЕА/мг) на 3 сутки составила всего 33,3%, на 5-е – 55%. Лепидоцид СК (БА-2000 ЕА/мг) практически не оказал токсичного действия на капустную тлю. Сравнительно высокую эффективность проявил Фитоверм, КЭ (2г/л) – 90%.
Растянутость периода выхода жуков брюквенного барида, скрытное питание, плотный листовой покров, защищающий насекомых от попадания рабочего раствора пестицида, резко снижают эффективность применяемых препаратов. С 2001 по 2007 гг. нами было проведено широкое испытание инсектицидов против вредителя: Базудина, Г (50г/кг), Базудина, ВЭ 60%, БИ-58 Новый, КЭ (400г/л), Карате Зеон, МКС (50г/л) и др. (рис.11).

Рис. 11. Биологическая эффективность препаратов против брюквенного барида. (Ряд 1- снижение численности вредителя после обработки (%) на 7 сутки; ряд 2 – то же на 14-е сутки).
В семеноводческих хозяйствах Дербентского района от брюквенного барида проводят 1-2 обработки до массового цветения семенников. По среднемноголетним фенологическим данным, начало отрождения личинок брюквенного барида приходится на 28. V±5,2. В связи с этим, мы существенно сдвинули сроки обработок с 27.IV на 25.V– 28.V (когда 75% растений капусты уже отцвело). Результаты испытаний показали, что двукратное опрыскивание посевов семенников капусты (первое - против перезимовавших жуков, второе - при отрождении личинок) обеспечило высокую смертность вредителя (80,5%) и превосходило однократную обработку по биологической эффективности. Отметим, что препараты Карате Зеон, МКС (50г/л) и Би-58 Новый, КЭ (400г/л) слабо подавляли вредителя. Их эффективность составляла 53,6% и 68,8% соответственно.
Таким образом, в результате испытаний выявлены наиболее эффективные инсектициды для защиты семенников капусты от наиболее вредоносных фитофагов.
Глава 8. Новая интегрированная система защиты семенников капусты
В основу оптимизированной системы защиты семенников капусты от фитофагов легли полученные нами данные исследований. Разработанная система базируется на результатах многолетнего фитосанитарного мониторинга и биоценотических принципах, позволяющая дифференцированно применять средства защиты в каждом конкретном случае. Суммируя результаты исследований 1998-2007 гг. мы предлагаем систему наблюдений и учётов за основными вредителями семенников капусты и разработанную интегрированную систему защиты в виде следующих схематичных таблиц.
Таблица 5. Интегрированная система защиты семенников капусты от основных вредителей в условиях юга России с учётом мониторинга фитосанитарного состояния
| Вредитель, фенологическая фаза развития растений | Методы обследования | Факторы повышающие вредоносность, экономический порог | Сигнализация | Защитные меры борьбы (профилактические, истребительные), сроки проведения защитных мероприятий |
| 1 | 2 | 3 | 4 | 5 |
| Крестоцветные блошки (за 1-2 дня до посева) | Сбор насекомых эксгаустером (30 и более жуков) для определения видового состава | Появление на сорняках:Barbarea vulgaris, Sinapis arvensis, Raphanus raphanistrum, Capsela bursa pastoris и др. | Перед посевом - 22 по 28.VI для позднеспелых сортов и гибридов; с 25. VII по 5. VIII среднеспелых сортов и гибридов и 15. VIII по 25. VIII - раннеспелых сортов и гибридов | Предпосевное протравливание семян (профилактически) Круйзер, КС (350 г/л) |
| Всходы (через 12-14 дней после появления) | 15-20 наложений ящика Петлюка на растения в открытых рассадниках | При повреждённости всходов выше ЭПВ (5-7% поврежденность растений) | Обработка рассады позднеспелых сортов и гибридов -5.VII±3; среднеспелых сортов и гибридов 17.VIII±4,2; раннеспелых сортов и гибридов - 14 IХ± 5,4 | Пролив почвы Актарой, ВДГ (250 г/кг) в 0,1% концентрации |
| 1 | 2 | 3 | 4 | 5 |
| Крестоцветные блошки, капустная моль, капустная тля Листовая мутовка (розетка) | Согласно методикам | При численности выше ЭПВ капустной тли (25% заселенных растений, (по 2-му баллу)). | За 2-3 дня до высадки рассады в грунт – на промышленную плантацию (2 – 3-я декады сентября) | Опрыскивание растений одним из перечисленных инсектицидов: Фастак, КЭ (100 г/л); Каратэ КЭ (50 г/л) и Децис Экстра, КЭ (125г/л); |
| Капустная тля, капустная моль Скручивание листьев и накопление листовой массы; завязывание кочана(2-3 декада октября) | Учет численности по методикам | При превышении численности ЭПВ капустной тли (25% заселенных растений, (по 2-му баллу)). | 1-2-я декады октября | Опрыскивание растений инсектицидами, применяемыми при низких положительных температурах: (одним из перечисленных) Децис Экстра, КЭ (125г/л), Фастак, КЭ (100 г/л), Золон, КЭ (350 г/л) |
| Фаза бутонизации | Определение численности вредителей по Z - образному маршруту: одну половину вдоль двух краев поля, а вторую по одной из диагоналей. При этом, пробы отбирают на расстоянии 15-20 м от края поля. | Проведение краевых обработок в начале заселения растений рапсовым цветоедом - 13. IV ±3,3. При численности мигрирующих особей капустной тли на клейких ловушках 15-20 особей на 1 ловушку в сутки | Опрыскивание растений в краевой полосе, шириной 5-10 м, 16-18.IV | Проведение барьерной обработки Золоном, КЭ (350 г/л) |
| 1 | 2 | 3 | 4 | 5 |
| Брюквенный барид (имаго), рапсовый цветоед, капустная тля В начале цветения семенников | Осматривается 100 растений Осмотр прикорневой части растений (бариды) | В период массового заселения вредителями | При численности вредителей выше ЭПВ: Брюквенного барида 0,5 экз. жуков на заселённое растение, рапсового цветоеда более 2 экз. на растение, капустной тли при численности крылатых особей в ловушках – более 24-25 экз. на ловушку в сутки. Наличие крестоцветных сорняков, отсутствие или низкая численность хищных жужелиц; отсутствие заражённых паразитами гусениц капустной моли, при заселении вредителем 20% растений. | Проведение сплошной обработки одним из перечисле6нных препаратов: Конфидор, ВДК (200 г/л) в норме расхода 0,2 л/га; Би-58 Новый (400 г/л) – 1,5 л/га; Моспилан, РП (200г/кг) |
| Брюквенный барид (личинка), капустная моль, капустная тля Фенофаза -конец цветения (70% растений - в фазе образования стручков) | Осмотр 100 растений. Подсчитывается среднее количество гусениц на растение (путём взятия проб с кочерыги) с последующей экспертизой в лаборатории | Сухая жаркая погода, при ГТК - 0,15-0,30 | В период начала отрождения личинок брюквенного барида, гусениц капустной моли (25-28.V). | Проведение комбинированной обработки против комплекса сосущих и грызущих вредителей Конфидором, ВРК (200 г/л) в норме расхода 0,5 л/га |
8.1. Экономическая эффективность интегрированной системы защиты семенников капусты
При традиционной комплексной зональной системе, защита семенников капусты сводилась к проведению плановых, сплошных календарных обработок против комплекса вредителей. Она не учитывала экономических порогов вредоносности фитофагов, роли естественных регуляторных механизмов, а также метеорологических особенностей каждого конкретного периода. Ниже приведены расчеты экономической эффективности при применении зональной системы защиты семенников капусты и разработанной нами интегрированной системы защиты в двух базовых хозяйствах МУП а/ф «Штул» и ДОС ВНИИР Дербентского района.
Таблица 6. Экономическая эффективность интегрированной системы защиты семенников капусты. МУП а/ф «Штул» (сорт Подарок), ДОСВНИИР (F1гибрид Экстра), 2007г.*
| Показатели | Единицы измерения | Зональная система защиты, принятая в Дербентском районе | Интегрированная система защиты семенников капусты | ||||||
| МУП а/ф «Штул» | ДОС ВНИИР | МУП а/ф «Штул» | ДОС ВНИИР | ||||||
| Урожайность | ц/га | 10,83 | 7,83 | 12,66 | 9,73 | ||||
| Прибавка урожая | ц/га | ----- | ---- | 1,83 | 1,9 | ||||
| Стоимость дополнительного урожая | руб/га | ---- | ----- | 31110 | 114000 | ||||
| З а т р а т ы: (стоимость инсектицидов с НДС, оплата труда механизатора, ГСМ, амортизация, затраты на уборку дополнительного урожая, прочие расходы на 1 обработку). | |||||||||
| Всего затрат | руб/га | 1475,9 | 1475,9 | 5332,8 | 5340,2 | ||||
| Окупаемость затрат | руб/га | ----- | --- | 8,1 | 29,5 | ||||
* Интегрированная система защиты семенников капусты в а/ф «Штул» применена на площади 2 га, зональная система – на 5 га. В ДОСВНИИР интегрированная система защиты применена на площади 1,5 га, зональная система – на 6 га.
Сравнительный анализ различных систем защиты семенников капусты в агрофирме «Штул» Дербентского района показал, что затраты на применение интегрированной защиты семенников капусты в расчёте на 1 га оказались на 3856,9 рублей больше, чем при применении комплексной зональной системы защиты. Стоимость полученного дополнительного урожая (при среднереализационной цене сортовых семян белокочанной капусты 170 руб. за 1 кг) при применении ИСЗ составила 31110 рублей с гектара. На каждый рубль затрат на защиту урожая получено 8,1 рубля дополнительного дохода.
Применённая в ДОС ВНИИР ИСЗ семенников капусты позволила собрать 190 кг дополнительного урожая гибридных семян с каждого гектара. Сумма реализации дополнительного урожая (при среднереализационной цене гибридных семян – 600 руб./кг) составила 114000 рублей. Окупаемость затрат в расчёте на рубль - 29,5 руб.
Урожай, полученный в результате применения интегрированной системы защиты семенников капусты, помимо количественных показателей отличался и качественными характеристиками. Результаты проведённого мониторинга посевных качеств семян (данные испытательной лаборатории ГСИ по РД) приведены в табл.7.
Таблица 7. Сравнительный анализ посевных качеств семян белокочанной капусты сорта Подарок и F1 гибрида Экстра в зависимости от принятой системы защиты. Испытательная лаборатория ГСИ по РД, 2007г.
| Показатели | Подарок (А/ф «Штул») | F1 Экстра, (ДОСВНИИР) | ||
| Зональная система защиты | Интегрированная система защиты | Зональная система защиты | Интегрированная система защиты | |
| Чистота, % | 99,5 | 99,7 | 99,7 | 99,8 |
| Всхожесть,% | 88 | 95 | 90 | 96 |
| Масса1000 семян, г | 5,41 | 5,75 | 3,83 | 4,68 |
Как показывают данные таблицы, в зависимости от принятой системы защиты семенников капусты от вредителей, всхожесть семян варьирует в довольно широких пределах. Разница во всхожести была на 5-6% выше при применении ИСЗ. Выполненность семян также зависела от системы защиты. Так, масса 1000 семян при применении ИСЗ семенников капусты у F гибрида Экстра составила 4,68 г, при проведении календарной схемы обработок - 3,83 г.
Качественные характеристики семян являются ориентиром для установления посевной годности и нормы высева. При всхожести семян 96% и чистоте 99,8% посевная годность составляет 95,8%. Сопоставляя качество полученных семян при различных системах защиты, нами были уточнены нормы высева семян. При установленной норме высева гибридных семян 250 г на 1 гектар со 100% посевной годностью, расчётная норма высева семян будет составлять 261 г/га; При всхожести семян 90%, чистоте 99,7% посевная годность составит 89,7%. Расчётная норма высева семян соответственно составит 278,7 г/га.
Выводы
1. По итогам многолетних биоэкологических исследований впервые удалось научно обосновать и разработать интегрированную систему фитосанитарных мероприятий по защите семенников белокочанной капусты от основных вредителей в условиях юга России. Система включает мониторинг видового состава вредителей, популяционной динамики численности вредителей, биологических особенностей, устойчивости сортов капусты к вредителям и прогноз вредоносности. Внедрение системы обеспечит существенное увеличение урожайности семенников белокочанной капусты и рентабельности семеноводства, снизит пестицидную нагрузку на агроценоз.
2. Впервые дан обзор энтомофауны агроценоза семенников капусты в условиях юга России. Выявлено до 70 видов насекомых, относящихся к 4 отрядам и 17 семействам, в том числе 14 видов фитофагов. Среди них наибольшую опасность для семенников капусты представляют брюквенный барид (Baris coerulescens Scop.), капустная тля (Brevicoryne brassicae L.), рапсовый цветоед (Meligethes aeneus Fab.), крестоцветные блошки (род Phyllotreta) и капустная моль (Plutella maculipennis Curt.).
3. Проведена видовая идентификация сообщества баридов в Дагестане. Выявлено 22 вида, среди которых доминирует – B. coerulescens Scop. (71,6% к общей численности). В. timida Rossi занимает 6,1 % от общей численности, B. scolopacea Germar – 3,6%, B. janthina Boheman – 3,1%, B. kirshi Desbrochers – 2,2%, B. semistriata Boheman – 2,4%. Уточнены морфологические признаки B. coerulescens, В. timida и др. видов баридов.
4. Впервые на основе многолетних учётов изучена биология, фенология, сезонная и многолетняя динамика численности основных вредителей семенников капусты. В условиях южного Дагестана появление 50% перезимовавших жуков брюквенного барида отмечается обычно 16. IV±7,7 дней. Начало откладки яиц приходится на 20. V ±4,1 дней. Средняя дата появления массового количества куколок соответствует 16. VII ±12 дней, массовое отрождение жуков нового поколения приходится на 27. VII ±10,5 дней. У рапсового цветоеда массовое появление жуков отмечается в середине апреля, при достижении эфф.t° - 24°С ±10,1°С. Массовая откладка яиц отмечается при эфф.t° - 47,9°С (26 IV ± 5,8 дней). Отрождение личинок рапсового цветоеда обычно наблюдается при достижении эфф.t° - 67,5°С. Численность крестоцветных блошек максимальна в 3-й декаде июля, первой декаде августа. Гусеницы капустной моли наиболее вредоносны в первой декаде октября. Для обоснованного проведения защитных мероприятий уточнены наиболее уязвимые фазы развития основных вредителей.
5. Анализ 12 К-факторных таблиц выживания основных вредителей семенников капусты позволил определить наиболее критические периоды в динамике численности, влияющие на дальнейший ход динамики численности фитофагов. Основными факторами смертности брюквенного барида является гибель во время перезимовки, а также тепловой шок (уровень k=0,04-0,06), рапсового цветоеда – каннибализм и хищники (уровень k=0,05), капустной моли – паразитоиды (уровень k = 0,14-0,20), капустной тли – паразитоиды (уровень k=0,15) и хищники (уровень k=0,02).
6. Численность и вредоносность капустной тли эффективно контролируется энтомофагами Diaeretiella rapae M'Intosh, Chrysopa formosa Brauer и C. carnea Stephens, капустной моли наездниками: Diadegma collaris Grav. D. semillausum Hell., D. fenestralis Holm. Установлен процент паразитирования капустной тли и капустной моли указанными энтомофагами.
7. В регулировании численности брюквенного барида в отдельные годы основную роль играют жужелицы Poecilus cupreus Linn. и Brachinus crepitans Daft. Установлено, что их численность существенно отличается в зависимости от предшественника семенников капусты, наибольшее количество хищников отмечено в люцерновом севообороте. Численность афидофага D. rapae в течение вегетации культуры неодинакова. Ограничивающее воздействие энтомофагов на популяцию капустной тли начинает проявляться только со второй половины июля, когда соотношение хищник жертва достигает минимум 1:40.
8. Установлены закономерности пространственного распределения популяции капустной тли, позволяющие определить сроки и количество миграций вредителя в целях своевременного и достоверного прогноза его численности. Было отмечено, что в ранневесенний период миграции насекомых незначительны (в среднем 24,4 экз. на ловушку в сутки), что связано с возрастными особенностями популяции. В 3-й декаде мая (3-4-е поколение) численность мигранток составляла 38,4 экз. на ловушку в сутки, в дальнейшем отмечается быстрый рост популяции капустной тли -129,7 экз./ловушку в сутки в июле, 224,3 экз./ловушку в сутки в сентябре.
9. С учетом изученных фактов вредоносности фитофагов на семенниках капусты были разработаны и уточнены их пороги вредоносности. ЭПВ брюквенного барида составил 0,05 экз. жуков на 1 растение; рапсового цветоеда -2 экз. на растение в фазу бутонизации; капустной тли - 25% заселенных растений семенной капусты (по 2-му баллу); крестоцветных блошек - 5 - 7% -я поврежденность всходов.
10. Ограничение вредоносности брюквенного барида возможно на основе использования разработанной нами прогностической модели, суть которой заключается в относительном изменении численности вредителя при изменении ГТК: при увеличении ГТК на 1% численность популяции вредителя уменьшается на 8%. Двухфакторная модель зависимости численности популяции брюквенного барида от ГТК: h=0,029 х12-0,2 х1+l,l6+0,01х2, совокупный коэффициент детерминации R=0,89.
11. Предложена экологически обоснованная система применения инсектицидов против основных вредителей семенников капусты в семеноводческих хозяйствах Дагестана, сохраняющая энтомофагов; она включает оптимизацию выбора инсектицидов, их дифференцированное применение, совмещение в одном технологическом процессе химической борьбы против комплекса основных вредителей семенников капусты.
Рекомендации производству
Для фитосанитарной стабилизации семеноводческих посевов белокочанной капусты юга России рекомендуется применение разработанной нами интегрированной системы защиты от основных вредителей.
Основными слагаемыми предлагаемой системы защиты являются:
- Постоянный мониторинг динамики численности популяций основных вредителей семенников капусты при одновременном обеспечении сохранения природных регуляторных механизмов (энтомофагов);
- Использование экспериментально установленных порогов вредоносности основных фитофагов семенников капусты: при численности вредителей, незначительно превышающих ЭПВ использование препаратов на биологической основе - Битоксибациллина, П (БА-1500 ЕА/мг) и Фитоверма, КЭ (2г/л);
- Оптимизация применения инсектицидов. Включение в систему защиты современного ассортимента инсектицидов с преимущественным использованием неоникотиноидных препаратов, применяемых в малых нормах расхода и высокоэффективных против комплекса основных вредителей семенников капусты;
- Использование приёмов поддержания высокой численности энтомофагов за счёт применения высокоселективных препаратов в безопасные для энтомофагов сроки, применения барьерных обработок, насыщения овощного севооборота многолетними бобовыми травами, позволяющее поддерживать высокую численность местных видов хищных жужелиц.
Список опубликованных работ по теме диссертации
- Мисриева Б.У. Фенология развития вредителей на семенниках белокочанной капусты при беспересадочном способе семеноводства /Г.Ф. Монахос, Д.В. Пацурия, Б.У. Мисриева, Т.Х. Касимов //Материалы междунар. науч.-практ. конф. "Селекция и семеноводство овощных культур в XXI в." - М., - 2000.- Т.2, - С. 89-90.
- Мисриева Б.У. Технология беспересадочного семеноводства гибридов поздней капусты, созданных на базе линий с мужской стерильностью (учеб.-метод. пособие) /Г.Ф. Монахос, А.В.Крючков, Д.В. Пацурия, Н.Н. Воробьева Лежкина А.А., Харламов Д.М., Судденко В.Г., Б.У. Мисриева, Т.Х. Касимов Т.Х. под ред. Монахос Г.Ф. - М.-2002.-12с.
- Мисриева Б.У. Гранулированный базудин против брюквенного барида /Б.У. Мисриева //Защита и карантин растений. – 2003.-№6.- С. 28-29.
- Мисриева Б.У. Эффективность базудина в борьбе с брюквенным баридом на семенниках капусты /Б.У. Мисриева //Докл. ТСХА /Московская с.-х. акад. им. Тимирязева. Москва.- 2004.- Вып. 276. - С. 507-509.
- Мисриева Б.У. Население жужелиц капустных агроценозов в южной зоне Дагестана /Б.У. Мисриева //Материалы Всероссийской научно-практической конференции «Интегрированная защита сельскохозяйственных культур и фитосанитарный мониторинг в современном земледелии» - Ставрополь.-2004г.
- Мисриева Б.У. Вредоносность брюквенного барида на семенниках капусты и меры борьбы с ним / Б.У. Мисриева // Материалы международной научно-практической конференции. – Краснодар. Куб ГАУ.- 2004.- С. 229-232.
- Мисриева Б.У. Биоресурсы капустного поля /Б.У. Мисриева //Защита и карантин растений.- 2005.- № 7. С. 27-28.
- Мисриева Б.У. Брюквенный барид на семенниках капусты /Б.У.
Мисриева //Защита и карантин растений.- 2005.- №12 - С. 18-19.
- Мисриева Б.У. Рапсовый цветоед на семенниках капусты в Дагестане /Б.У.Мисриева //Овощеводство.- 2006.- №8.-С.59-61.
- Мисриева Б.У. Капустная тля в Дагестане /Б.У.Мисриева //Защита и карантин растений.- 2006.- №11 - С. 36-37.
- Мисриева Б.У. Крестоцветные блошки на семенниках капусты /Б.У. Мисриева //Сб. научн.тр. Краснодар.- 2006.- С.253-261.
- Мисриева Б.У. Вредоносность и экономический порог вредоносности брюквенного барида на семенниках белокочанной капусты /Б.У. Мисриева //Гавриш.- 2006.- № 5. - С. 10-12.
- Мисриева Б.У. Как рассчитать экономический порог вредоносности рапсового цветоеда на семенниках капусты /Б.У. Мисриева// Картофель и овощи.- 2006.- № 6. - С. 29-30.
- Мисриева Б.У. Экономический порог вредоносности крестоцветных блошек на семенниках капусты /Б.У. Мисриева, А.М. Мисриев //Картофель и овощи.- 2007.- № 3. - С. 30 – 31.
- Мисриева Б.У. Оценка сортов и гибридов семенной белокочанной капусты на устойчивость к брюквенному бариду (Baris coerulescens Scop.) в Дагестане /Б.У. Мисриева //Овощеводство.-2007.-№3.-С. 62-64.
- Мисриева Б.У. Видовой состав баридов в Дагестане /Б.У. Мисриева //Докл. РАСХН.- 2007.- №4. - С. 29-30.
- Мисриева Б.У. Динамика численности капустной тли и уровень ее паразитирования афидофагами /Б.У. Мисриева //Защита и карантин растений. – 2007.- № 12.
- Мисриева Б.У. Биологическая эффективность неоникотиноидов на семенниках капусты /Б.У. Мисриева //Овощеводство.- 2007. - № 5.- C. 52-54.
- Мисриева Б.У. Таблицы выживания популяций рапсового цветоеда Meligetes aeneus Fab. на семенниках белокочанной капусты в условиях Южного Дагестана /С.Я. Попов, Б.У. Мисриева //Известия ТСХА. -2007г. - Вып. 4.- С. 93-100.
- Мисриева Б.У. Динамика численности популяций брюквенного барида на семенниках капусты в Дагестане /Б.У. Мисриева //Доклады РАСХН.- 2008.- №1.- С.29-30.
- Мисриева Б.У. Биологическая эффективность биопрепаратов в борьбе с вредителями семенников капусты в южной зоне Дагестана /Б.У. Мисриева, Е.Г. Гаджимусатапаева //Овощеводство. – 2008.-№5.-С. 64-66.
- Мисриева Б.У. Беспересадочное семеноводство капусты в Дагестане / Б.У. Мисриева // Университетская экология [международный сборник научных трудов] Махачкала. -2007.- С. 214-215.
- Мисриева Б.У. Сопряженность динамики развития фитофагов и фенологии семенных растений капусты /Б.У. Мисриева, А.М. Мисриев // Университетская экология [международный сборник научных трудов] Махачкала.- 2007.- С. 215-217.