

Влияние гравитационной разгрузки на реакцию сердечного ритма у обезьян, вызванную стимуляцией отолитов
На правах рукописи
МИЛЛЕР
Наталья Всеволодовна
ВЛИЯНИЕ гравитационной РАЗГРУЗКИ
НА РЕАКЦИЮ СЕРДЕЧНОГО РИТМА У ОБЕЗЬЯН,
ВЫЗВАННУЮ СТИМУЛЯЦИЕЙ ОТОЛИТОВ
14.00.32 – Авиационная, космическая и морская медицина
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Москва – 2008
Работа выполнена в Государственном научном центре Российской Федерации – Институте медико-биологических проблем РАН
Научный руководитель: кандидат биологических наук
Бадаква Анатолий Михайлович
Официальные оппоненты: доктор биологических наук, профессор
Виноградова Ольга Леонидовна
доктор биологических наук, профессор
Шульговский Валерий Викторович
Ведущая организация: ГНИИИ военной медицины МО РФ
Защита диссертации состоится «___» __________ 2008 г. в ____ часов на заседании диссертационного совета Д002.111.01 в ГНЦ РФ – Института медико-биологических проблем РАН по адресу: 123007, Москва, Хорошевское шоссе, д. 76 А.
С диссертацией можно ознакомиться в библиотеке ГНЦ РФ – ИМБП РАН.
Автореферат разослан «____» _____________ 2008 г.
Ученый секретарь
диссертационного совета
доктор биологических наук Левинских М.А.
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность темы
Многочисленные исследования последних лет, проведенные как в экспериментах на животных, так и в исследованиях с участием человека, свидетельствуют об участии вестибулярной системы в регуляции кровообращения (Yates, Miller, 1998; Ray, Hume, 1998; Radtke et al, 2003). Показано существование прямого распространения возбуждения от вестибулярных рецепторов к эффекторам вегетативной нервной системы (обзоры: Yates, 1992; 1996 a; 1996 b).
Адаптивное торможение вестибулярных структур в условиях невесомости может вызывать уменьшение участия вестибулярного входа в компенсаторных реакциях кровообращения на постуральные возмущения в послеполетный период, в том числе быть одним из факторов, влияющих на послеполетную ортостатическую устойчивость.
Основной массив данных, связанных с изучением вестибуло-вызванных сердечно-сосудистых рефлексов, получен на наркотизированных кошках. Изучались главным образом реакции, связанные с воздействием вестибулярной стимуляции на активность симпатического отдела вегетативной нервной системы. Возможное влияние вестибулярного раздражения на сердце, опосредуемое парасимпатическими структурами, оценивалось как ничтожное (Yates, 1992; Yates, Kerman, 1998; Yates et al., 2000). Вместе с тем снижение парасимпатических влияний на сердце является наиболее срочной компенсаторной реакцией при переходе в ортостатическое положение (Осадчий, 1982). Поэтому исследование реакции сердечного ритма (СР) на кратковременную стимуляцию отолитов у бодрствующих обезьян, физиологические реакции которых близки к наблюдаемым у человека, в норме и в условиях гравитационной разгрузки является актуальной задачей.
Цель работы
Оценить влияние гравитационной разгрузки на реакцию СР, вызванную стимуляцией отолитов, у бодрствующих обезьян.
Задачи исследования
- Охарактеризовать изменения СР у обезьян в ответ на кратковременную стимуляцию отолитов.
- Определить влияние длительной антиортостатической гипокинезии (АНОГ) на реакцию СР у обезьян, вызванную стимуляцией отолитов.
- Оценить изменения реакции СР на стимуляцию отолитов у обезьян в космическом полете (КП) и после его завершения.
- Сопоставить изменения реакции СР в КП и после его завершения с динамикой ответа вестибулярных нейронов на стимуляцию отолитов у обезьян в КП.
Научная новизна
Впервые обнаружено существование коротколатентного рефлекторного учащения СР на стимуляцию отолитов, имеющего парасимпатическую природу.
Впервые показана линейная зависимость изменения величины интервала R-R ЭКГ (RR) в ответ на кратковременную стимуляцию отолитов от его начального уровня (RR0), следствием которой является уменьшение влияния кратковременной стимуляции отолитов на СР с ростом исходной ЧСС (ЧСС0).
Впервые показано, что длительная АНОГ у обезьян вызывает уменьшение влияния кратковременной стимуляции отолитов на СР.
Впервые охарактеризованы изменения реакций СР в ответ на стимуляцию отолитов у обезьян в течение КП, а также после его завершения, и показана связь этих изменений с динамикой возбудимости вестибулярных нейронов.
Научно-практическая значимость
Разработанные методические приемы тестирования реакции СР в ответ на кратковременную стимуляцию отолитов позволяют оценивать участие отолитовой системы в регуляции кровообращения.
Данные о влиянии гравитационной разгрузки на реакции СР в ответ на стимуляцию отолитов, полученные на обезьянах, могут быть использованы в качестве теоретической основы для проведения дальнейших исследований участия вестибулярной системы в быстрых хронотропных реакциях сердца и разработки методов прогностической оценки послеполетной ортостатической неустойчивости у человека.
Основные положения, выносимые на защиту
- Кратковременная стимуляция отолитов у обезьян вызывает учащение СР, которое имеет рефлекторный характер и определяется уменьшением парасимпатических влияний на сердце.
- Гравитационная разгрузка уменьшает участие отолитовой системы в быстрых хронотропных реакциях сердца.
Апробация работы
Основные результаты диссертационной работы доложены и обсуждены: на заключительном российско-американском симпозиуме «Медико-биологические исследования по программе «Наука-НАСА» (Хантсвилл, США, 1997); на международном симпозиуме по исследованию приматов в космосе в рамках ХI конференции «Космическая биология и авиакосмическая медицина» (Москва, 1998; 2 доклада); на 13 международном симпозиуме «Человек в космосе» (Санторини, Греция, 2000); на российской конференции «Организм и окружающая среда» (Москва, 2000; 3 доклада); на 23 международной конференции по гравитационной физиологии (Стокгольм, Швеция, 2002 г.).
Диссертация апробирована на заседании секции «Космическая физиология и биология» Ученого совета ГНЦ РФ - ИМБП РАН.
По теме диссертации опубликовано 8 печатных работ.
Структура и объем диссертации
Диссертация изложена на 132 страницах машинописного текста и состоит из введения, литературного обзора, описания методик, результатов исследования, обсуждения, выводов и списка цитируемой литературы. Список цитируемой литературы включает 186 источника, из которых 38 опубликованы в отечественных изданиях, 148 – в иностранных. Материал диссертации иллюстрирован 33 рисунками и 1 таблицей.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Работа проведена на 29 обезьянах-самцах macaca mulatta возрастом 3-5 лет, массой 4-6 кг в рамках комплексных программ: контракта «Россия-НАСА» (NAS-15-10110), исследования влияния длительной АНОГ на физиологические системы обезьян и исследований на биоспутнике №11.
Операционная подготовка животных включала: 1) подкожное вживление электродов для отведения ЭКГ и сигналов реопневмограммы (РПнГ) (Гуревич, 1982) всем животным; 2) установку на черепе (9 животных) армированного металлом акрилового кольца (Сирота и др., 1992) для фиксации головы, монтажа предусилителя сигналов нейрональной активности (НА) и крепления защитного колпака с датчиком скорости поворота головы; 3) установку на черепе металлических канюль (4 обезьяны) для введения микроэлектродов (Сирота и др., 1992).
Все хирургические процедуры и экспериментальные манипуляции с животными были одобрены Биоэтической комиссией ГНЦ РФ - ИМБП РАН.
Проведено три серии экспериментальных исследований.
В первой серии исследовали влияние кратковременной стимуляции отолитов на реакцию СР. В первой части, проведенной на 8 обезьянах с жестко фиксированной головой, для стимуляции отолитов по направлению интерауральной линии использовали затухающие колебания четырехштанговых качелей с начальным скачком ускорения 0,16 и 0,08 g. Вторая часть первой серии исследований проведена на 20 обезьянах. Животное фиксировали в ложементе кресла, не ограничивая движения головы и конечностей; кресло устанавливали в полетной капсуле. Для стимуляции отолитов использовали последовательность из 30 одиночных циклов вертикального перемещения кресла с начальным скачком ускорения 0,08 g. Регистрировали сигналы ЭКГ, РПнГ и позиции кресла.
Во второй серии, проведенной на 10 из 20 животных первой серии, исследовали влияние 30-суточной АНОГ (-6) на реакцию СР, вызванную стимуляцией отолитов. Обезьяны были протестированы через 1 и 7 сут после завершения АНОГ таким же образом, как и во второй части первой серии исследований.
Третья серия исследования проведена на 2 обезьянах в автоматическом режиме в КП на биоспутнике. В нечетные сутки (начиная с 3 сут) тестировались реакции СР на восемь предъявлений одного цикла колебания ложемента вдоль продольной оси тела животного. В четные сутки осуществлялось тестирование ответа вестибулярных нейронов на последовательность 40 синусоидальных колебаний кресла с обезьяной вдоль ее продольной оси с частотой 1,67 Гц. Через сутки после завершения КП тестирование реакции СР осуществляли на четырехштанговых качелях.
Регистрировали: сигналы ЭКГ и РПнГ - при тестировании реакций СР; нейрональную активность (НА) популяции нейронов и вертикальную скорость поворота головы - при тестировании ответа вестибулярных нейронов в КП; сигналы позиции и ускорения качелей с животным – во всех сериях исследований.
При обработке сигнала ЭКГ последовательность интервалов R-R преобразовывали путем линейной интерполяции в регулярную последовательность отсчетов с шагом времени 50 мс (огибающую интервалограммы) и усредняли реакции интервала R-R за несколько предъявлений стимула синхронно с его началом. Динамику СР представляли в виде изменения длительности интервала R-R или в виде изменения ЧСС.
При обработке сигнала РПнГ изменение электрического сопротивления грудной клетки, связанного с дыхательным циклом, относили к базовому сопротивлению грудной клетки (Бадаква, 1989).
Сигнал НА оцифровывали с частотой 40 кГц, выделяли спайки отдельных нейронов с помощью оконного дискриминатора и строили внутристимульные гистограммы (Волошин, 1987): период стимула разбивали на фиксированное количество отрезков постоянной длительности (бинов), определяли среднее за 10 циклов стимула количество спайков, попадающих в каждый бин, и представляли его в виде частоты импульсов в секунду.
При анализе модуляции активности нейронов отолитовым стимулом использовали аппроксимацию внутристимульных гистограмм гармоническими функциями с основной и кратными частотами стимула методом наименьших квадратов (Schor et al., 1984). Т.к. стабилизация движений глаз и головы во время линейных перемещений осуществляется при взаимодействии отолитовых и зрительных сигналов, и отолитовый вход, отвечающий на ускорение, преобладает в области высоких частот, а зрительный вход, отвечающий на скорость, преобладает в области частот ниже 0,75 Гц (Borel, Lacour, 1992), то для исключения участия зрительного входа в модуляции НА первая гармоника была разложена на составляющие, соответствующие ускорению (sint) и скорости (cost):
A1sin(t+) = А sin(t) + B cos(t), где A= A1sin и B= A1cos,
что позволило выделить модуляцию НА, соответствующую ускорению кресла, и характеризовать динамику возбудимости вестибулярных нейронов в КП изменением коэффициента A.
Полученные данные статистически обрабатывали, используя критерии Стьюдента и Фишера. В случае повторных измерений применяли поправку Бонферони. Критический уровень достоверности принимали равным 0,05.
РЕЗУЛЬТАТЫ ЭКСПЕРИМЕНТОВ И ИХ ОБСУЖДЕНИЕ
Влияние кратковременной стимуляции отолитов на реакцию сердечного ритма
Начальное движение качелей с ускорением 0,16 g вызывало у обезьян быстрое укорочение интервала R-R и последующее более медленное его возвращение к начальному уровню (рис. 1). В ряде случаев в динамике интервала R-R наблюдалась вторая волна его уменьшения. Укорочение интервала R-R при неизменном или даже углубленном дыхании сопровождалось уменьшением или исчезновением его дыхательной вариативности, что свидетельствует о снижении парасимпатических влияний на сердце в ответ на стимуляцию отолитов.
 | Рис.1. Пример реакции интервала R-R и дыхания у обезьяны в ответ на стимуляцию отолитов при движении качелей с начальным ускорением 0,16 g. Сверху вниз: интервалограмма, РПнГ, ускорение качелей и метка времени (1 с). Вертикальная линия показывает начало стимуляции. |
Однако движение с начальным ускорением 0,16 g, как правило, сопровождалось длительными периодами выраженной двигательной активности животного, что затрудняло получение достаточного для усреднения количества данных в течение даже нескольких экспериментальных дней. Движение качелей с начальным ускорением 0,08 g вызывало менее выраженные начальные изменения СР (рис. 2), но гораздо реже сопровождалось двигательными артефактами. При многократном повторном предъявлении стимула этой величины реакция СР не угасала и была хорошо воспроизводима.
Тестирование реакции СР в ответ на стимуляцию отолитов с начальным ускорением 0,08 g у 8 обезьян, представленной в виде

Рис. 2. Усредненные реакции СР, представленные динамикой ЧСС (M±m), у обезьяны при движении качелей с начальными значениями ускорения 0,16 g (А) и 0,08 g (Б). Вертикальные линии – начало стимуляции. * - P<0,05
динамики ЧСС (рис. 3), показало, что быстрое и достоверное начальное увеличение ЧСС, сменяющееся более медленным ее уменьшением, наблюдалось у всех животных. Более медленное повторное учащение СР, после которого ЧСС вновь постепенно возвращалась к престимульному уровню, наблюдалось только у 4 из 8 обезьян (левая панель) и требует отдельных исследований.
 | Рис. 3. Усредненные реакции ЧСС (M±m) у восьми обезьян при движении качелей с начальным значением ускорения 0,08 g. Внизу - сигнал ускорения качелей. Числа на графиках – номера обезьян. * - P<0,05. |
Начальное увеличение ЧСС у обезьян нельзя отнести к вегетативному компоненту ориентировочной реакции животного, так как устойчивая воспроизводимость реакции СР и отсутствие ее угашения при многократных предъявлениях стимула как в один, так и в разные экспериментальные дни, исключают такое предположение. Неспецифические реакции «испуга» на предъявление различных стимулов у животных (Casto et al., 1989) и человека (Bradley et al., 1993) угасают со временем, становясь меньшими при многократных предъявлениях стимула.
Данные, полученные на обезьянах, согласуются с результатами исследований отолитово-вызванных реакций СР у человека (Yates et al., 1999; Radtke et al., 2003; Yates, Bronstein, 2005; Jauregui-Renaud et al., 2006), показавшими отсутствие угасания реакции СР на повторные кратковременные движения всего тела или головы испытуемого.
Оценка латентного периода реакции СР (Radtke et al., 2003) у обезьян на стимуляцию отолитов, проведенная путем сопоставления значений трех интервалов R-R (интервала, во время которого начиналась стимуляция, предшествующего и последующего интервалов) показала, что латентный период начального изменения СР можно считать заметно меньшим, чем 600 мс. Так как симпатические нервные влияния на сердце осуществляются с заметно большей задержкой (Mokrane, Nadeau, 1998), очевидно, что укорочение интервала R-R являлось следствием торможения парасимпатических влияний на сердце.
Изменения СР, вызванные увеличением проприоцептивной активности и опосредуемые вагусом (Gladwell, Coote, 2002; Gladwell et al., 2005), имеют латентный период, сопоставимый с обнаруженным при стимуляции отолитов у обезьян. Однако при анализе данных обезьян были использованы только те реакции интервала R-R на стимуляцию отолитов, которые не сопровождались двигательной активностью животного и, соответственно, не могли быть опосредованы изменениями проприоцептивного входа, связанными с действием отолитово-моторных рефлексов.
Однотипный характер реакций R-R на затухающие колебания качелей и быстрое изменение позиции качелей (рис. 4 А, Б) свидетельствуют об определяющей роли начального скачка ускорения в развитии реакции СР на стимуляцию отолитов. Однократное вертикальное перемещение кресла с обезьяной с начальным скачком ускорения 0,08 g, которое в отличие от горизонтального можно было реализовать в полете на биоспутнике, вызывало реакцию СР, аналогичную наблюдавшейся при тестировании на качелях (рис. 4 В).
| А | Б | В |
 |  | 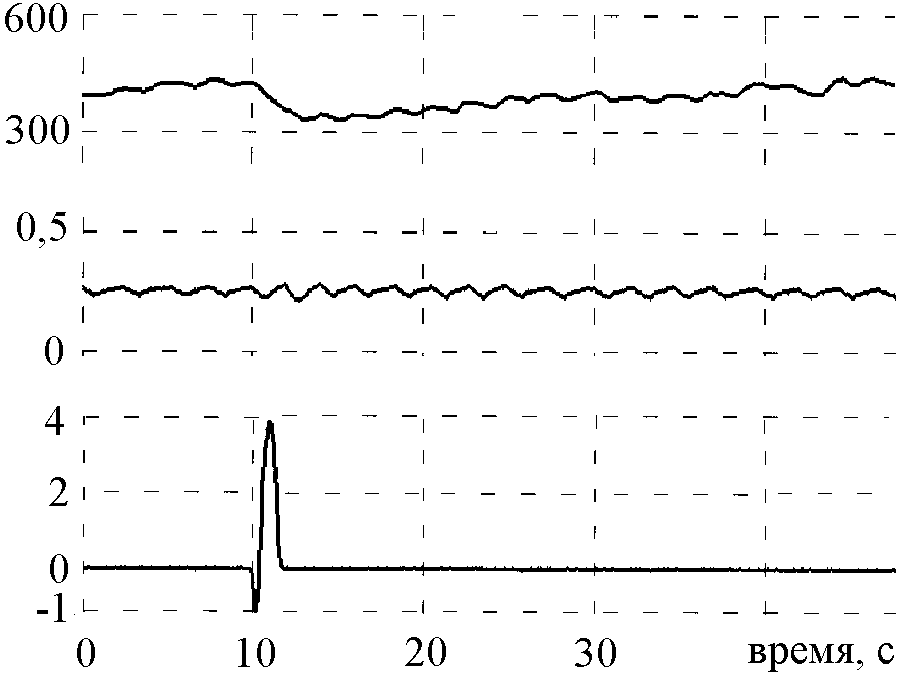 |
Рис. 4. Сравнение реакций интервала R-R у обезьяны на качание качелей (А), изменение их позиции (Б) и вертикальное перемещение кресла. Сверху вниз: интервал R-R (мс), РПнГ (отн. ед.), позиция качелей или кресла (см).
Величина укорочения интервала R-R (RR) в ответ на стимуляцию отолитов как у 8 обезьян при движении качелей, так и у 20 обезьян при кратковременном вертикальном перемещении кресла (с одним и тем же начальным ускорением 0,08 g) зависела от значения исходного интервала R-R (RR0): большим значениям RR0 соответствовало большее абсолютное значение RR (рис. 5). Обе зависимости характеризовались высокими значениями коэффициента линейной корреляции (R=-0,88; P<0,0001 для 8 обезьян и R=-0,91; P<0,0001 для 20 обезьян). При этом различия между реакциями интервала R-R на стимуляцию отолитов в горизонтальном и вертикальном направлениях не были обнаружены: соответствующие линии регрессии (RR = 75,6-0,29·RR0 и RR = 47,7-0,22·RR0) достоверно не отличались.
Обнаруженная закономерность изменения RR от RR0 представляет собой усредненную по группе исследуемых животных характеристику воздействия стимуляции отолитов с определенным начальным ускорением (0,08 g в конкретном случае) на СР и
зависимости реакции СР от его исходного значения. При всех значениях RR0, лежащих правее точки пересечения линии регрессии с осью RR0 (RRмин – минимальное значение RR0), отношение RR к

Рис. 5. Зависимости RR от RR0. А: у 8 обезьян при стимуляции отолитов на качелях (по данным двух тестирований для каждого животного). Б: у 20 обезьян при вертикальном перемещении кресла.
разности между RR0 и RRмин (коэффициент наклона линии регрессии) является постоянным, характеризуя чувствительность СР
к стимуляции отолитов. При увеличении чувствительности СР к стимуляции отолитов коэффициент наклона линии регрессии должен соответственно увеличиваться, а при уменьшении - уменьшаться.
Значение RRмин ограничивает снизу область значений RR0, при которых существует реакция СР на стимуляцию отолитов, характеризуя отсутствие парасимпатических влияний на сердце при RR0= RRмин и, тем самым, отражая усредненный по группе обезьян уровень влияния симпато-адреналовой системы на активность пейсмекера сердца. При такой интерпретации данных разность между RR0 и RRмин у каждого животного отражает индивидуальный уровень парасимпатических влияний на сердце.
Таким образом, кратковременная стимуляция отолитов вызывает у обезьян коротколатентное учащение СР, связанное с укорочением интервала R-R вследствие торможения парасимпатических влияний на сердце. Между величиной уменьшения интервала R-R в ответ на кратковременную стимуляцию отолитов и исходной величиной интервала R-R у обезьян существует линейная зависимость, причем с уменьшением исходной величины интервала R-R ответная реакция снижается, то есть уменьшается влияние отолитовой стимуляции на интервал R-R.
Влияние длительной АНОГ на реакции СР, вызванные стимуляцией отолитов
Исследование влияния АНОГ на реакции СР, вызванные стимуляцией отолитов, показало, что зависимость RR от RR0 у 10 обезьян через сутки после завершения АНОГ (RR = 28,7-0,12· RR0)
достоверно отличалась (P=0,0003) от аналогичной зависимости, полученной в контрольном исследовании у 20 животных (RR = 47,7 - 0,22·RR0) и имела достоверно меньший наклон (P=0,043), а RRмин сместилась в сторону больших значений (239±48 мс по сравнению с 216±9 мс), хотя и не достоверно (рис. 6А). У пяти из десяти животных RR находились за пределами 95 % доверительной области для значений, полученной по результатам контрольного исследования. У двух из них реакция на стимуляцию отолитов отсутствовала.
 |  |
Рис. 6. Зависимости RR от интервала RR0 у 10 обезьян через сутки (А) и через 7 суток (Б) после завершения АНОГ в сравнении с данными контрольного исследования (n=20).
Тонкой линией и пунктиром показаны линия регрессии и ее 95% доверительная область для значений в контрольном исследовании. Жирными линиями показаны линии регрессии, построенные по данным (точки), полученным после завершения АНОГ.
Через 7 сут после завершения АНОГ (рис. 6Б) у всех обезьян реакция интервала R-R в ответ на стимуляцию отолитов находилась в пределах 95% доверительной области для значений, полученной по результатам контрольного исследования. Линия регрессии (RR = 66,89-0,28· RR0) не отличалась достоверно от контрольной.
Влияние АНОГ на реакцию СР, вызванную стимуляцией отолитов, через сутки после завершения АНОГ проявилось главным образом в снижении чувствительности реакции СР к отолитовому стимулу, которое может быть обусловлено двумя причинами: во-первых, снижением отолитовой возбудимости и, во-вторых, изменением состояния механизмов регуляции СР.
Изменение отолитовой возбудимости во время АНОГ при неизменном гравитационном воздействии могло быть вызвано перераспределением жидких сред организма и изменениями мозгового кровообращения, имеющими значительную индивидуальную вариативность (Лапаев и др., 1983). Однако ввиду противоречивости данных о вестибуло-вегетативных расстройствах во время АНОГ, увеличение возбудимости рецепторного отдела отолитовой системы под воздействием АНОГ не считается реальным (Генин, Пестов, 1997). Более вероятной причиной увеличения отолитовой возбудимости и ее последующего адаптивного снижения во время АНОГ может быть, согласно гипотезе И.Б.Козловской о существенной роли опорной афферентации в сенсорном обеспечении деятельности различных физиологических систем (Крейдич и др., 1982; Козловская и др., 1984; Киренская и др., 1985; Григорьева, Козловская, 1987), увеличение возбудимости нейронов вестибулярных ядер, связанное с опорной разгрузкой.
Отсутствие реакции СР на стимуляцию отолитов у двух обезьян через сутки после завершения АНОГ свидетельствует о смещении RRмин в сторону больших значений, что означает сниженное влияние симпато-адреналовой системы на СР у этих животных при отсутствии парасимпатических влияний на СР.
Таким образом, в целом, уменьшение чувствительности СР к стимуляции отолитов после завершения АНОГ может быть результатом как адаптивного снижения возбудимости вестибулярных нейронов, опосредующих передачу возбуждения к бульбарным парасимпатическим структурам, так и изменения состояния механизмов регуляции СР (баланса симпатических и парасимпатических влияний на сердце).
Влияние микрогравитации на реакции СР, вызванные стимуляцией отолитов
Исследование в КП показало, что микрогравитация существенно влияет на реакции СР у обезьян, вызванные стимуляцией отолитов. У обезьяны 357 линии регрессии RR по RR0, построенные для каждого сеанса тестирования в течение КП, достоверно отличались от контрольной во все сутки КП, кроме 7 сут (рис. 7А). Однако коэффициенты корреляции между RR и RR0 высоки и достоверны лишь в 5, 11 и 13 сут КП. Значения RRмин сдвигались в сторону больших значений в течение КП. В 11 и 13 сут КП значения RRмин достигали значений 383±43 мс и 391±39 мс соответственно, достоверно отличаясь (P<0,0001 в обоих случаях) от значения для контрольной линии регрессии (216±10 мс). Угол наклона линии регрессии достоверно (P=0,045) увеличен в 5 сут КП (-0,53 ±0,17) в сравнении с контрольной величиной (-0,22 ±0,02) и в еще большей степени превышает контрольную величину к концу КП, достигая значений -0,81±0,16 в 11 сут (P<0,0001) и -0,98±0,16 в 13 сут КП (P<0,0001).
 |  |
Рис. 7. Динамика зависимости RR от RR0 у обезьян 357 (левая панель) и 484 (правая панель) в течение КП (числа на поле графиков – сутки КП). Тонкой линией показана линия регрессии в контрольном исследовании. Жирными линиями показаны линии регрессии, построенные по данным (точки), полученным в КП. На поле каждого графика приведены значение коэффициента корреляции и его достоверность.
У обезьяны 484 линии регрессии RR по RR0, построенные для каждого сеанса тестирования в течение КП, достоверно отличались от контрольной в 3, 7 и 9 сут КП (рис.8Б). Однако коэффициенты корреляции между RR и RR0 высоки и достоверны лишь в 3, 5, и 9 сут КП. Значение RRмин сдвигалось у обезьяны 484 в сторону больших величин от 3 к 7 сут КП, достигая в 7 сут значения 445±163 мс и достоверно отличаясь от контрольного уровня (216±9 мс; P=0,046), и затем возвращалось к контрольному уровню. Величина угла наклона линии регрессии достоверно превышала контрольный уровень (-0,22 ±0,02) в 7 и 9 сут КП (-0,88±0,47, P=0,044 и -0,67±0,21, P=0,019, соответственно), и не отличалась от контрольного значения в остальные сут КП.
Полученные результаты показали, что у обеих обезьян чувствительность СР к стимуляции отолитов возрастала в начальный период КП, продолжала увеличиваться у обезьяны 357 и возвращалась к контрольному уровню у обезьяны 484 к концу КП. Динамика величины RRмин в течение КП была аналогична динамике чувствительности СР: значение RRмин увеличивалось у обеих обезьян в первой половине КП, продолжало увеличиваться у обезьяны 357 и возвращалось к контрольному уровню у обезьяны 484 к концу КП
Увеличение чувствительности СР к стимуляции отолитов можно рассматривать как результат увеличения возбудимости нейрональной цепи, опосредующей передачу стимула на бульбарные парасимпатические структуры, тогда как смещение RRмин в область больших значений и уменьшение разности между RR0 и RRмин свидетельствует об уменьшении как симпатических, так и парасимпатических влияний на сердце. Возможной причиной уменьшения парасимпатических влияний может быть изменение возбудимости системы хронотропной регуляции сердечной деятельности в микрогравитации вследствие ослабления действия барорефлекторных влияний на сердце (Cooke et al., 2000; Cox et al., 2002).
Увеличение чувствительности СР к стимуляции отолитов в микрогравитации может быть результатом увеличения возбудимости вестибулярных нейронов, опосредующих передачу возбуждения к бульбарным парасимпатическим структурам.
У обеих обезьян активность популяций нейронов была зарегистрирована в области комплекса вестибулярных ядер, лежащей каудальнее ЛВЯ, включающей в себя как МВЯ, так и медиальную часть нижнего вестибулярного ядра, и опосредующей вестибуло-вегетативные ответы (Yates et al., 1993a,b, 1994; Xu et al., 2002). Визуальный анализ активности вестибулярных нейронов, зарегистрированных у обезьяны 357 в течение КП (рис. 8), показал, что амплитуда явно различимых спайков не изменялась в течение КП, а модуляция частоты спайков была связана с колебаниями позиции кресла. Увеличение частоты спайков предшествовало положительной фазе скорости поворота головы, т.е. не являлось следствием стимуляции вертикальных полукружных каналов. В целом, данные визуального анализа позволили считать, что в течение КП у обезьяны 357 регистрировалась активность одной и той же популяции вестибулярных нейронов, отвечающих на отрицательную фазу ускорения перемещения кресла с обезьяной в направлении продольной оси тела.

Рис. 8. Фрагменты регистрации активности популяции вестибулярных нейронов и скорости поворота головы в сагиттальной плоскости при синусоидальном перемещении кресла с животным с частотой 1,67 Гц у обезьяны 357 в течение КП. В каждом фрагменте длительностью 2 с показаны (сверху вниз): сутки КП, НА, скорость поворота головы и позиция кресла.
Модуляция активности исследуемой популяции вестибулярных нейронов, вызванная отолитовой стимуляцией и характеризующая их возбудимость (оцениваемая по изменению амплитудного коэффициента аппроксимации нейрональной активности популяции вестибулярных нейронов) у обезьяны 357 (рис. 9А) увеличивалась от 2 к 6 сут полета (от 2,9±3,6 до 99,6±3,5 имп/с), снижалась, возможно, вследствие адаптивного торможения, к 10 сут до уровня 2 сут и вновь увеличивалась, хотя и в меньшей мере, в 12 и 14 сут КП (50,9±3,8 и 50,4±3,8 имп/с соответственно), оставаясь повышенной по сравнению со 2 сут к концу полета.

Рис. 9. Изменение амплитудных коэффициентов аппроксимации нейрональной активности популяции вестибулярных нейронов у обезьян 357 (А) и 484 (Б) в течение КП. В – изменение амплитуды модуляции «пачковой» нейрональной активности у обезьяны 484. * – достоверные отличия от значения во вторые сутки КП (P<0,01).
В отличие от обезьяны 357, у которой в популяции регистрируемых нейронов выделялись высокоамплитудные и низкоамплитудные спайки, НА у обезьяны 484 состояла из спайков, близких друг к другу по форме и амплитуде. Поэтому на основании
визуального анализа амплитуд спайков нельзя судить о неизменности состава исследуемой популяции нейронов. Однако в течение всего КП (за исключением 10 сут) в области, соответствующей положительному ускорению перемещения кресла, наблюдалась характерная «пачковая» НА, что позволяет предполагать постоянство состава исследуемой популяции нейронов у обезьяны 484 в ходе КП. Соответственно, к анализу изменения НА у обезьяны 484 в КП (рис. 9Б) была добавлена оценка изменения амплитуды модуляции «пачковой» нейрональной активности на участке цикла колебания кресла, соответствующего положительной фазе ускорения перемещения кресла (рис. 9В).
Амплитудный коэффициент аппроксимации нейрональной активности популяции вестибулярных нейронов у обезьяны 484 (рис.9 Б) был достоверно увеличен лишь в 8 сут КП по сравнению со значением в 2 сут КП (14,2±2,6 имп/с по сравнению с 0,6±2,6 имп/с; P=0,011) и не отличался достоверно от уровня 2 сут к концу полета. Изменения амплитуды модуляции «пачковой» нейрональной активности (рис. 10 В) близки к динамике амплитудного коэффициента аппроксимации: амплитуда была увеличена в начальной части КП (148,4±22,7 имп/с в 4 сут по сравнению с 59,7±16,2 имп/с в 2 сут; P=0,031) и не отличалась в конце полета от уровня во 2 сут КП.
Таким образом, у обезьяны 484 модуляция активности исследуемой популяции вестибулярных нейронов, вызванная стимуляцией отолитов, увеличивалась, как и у обезьяны 357, в начальный период КП, однако к концу КП, в отличие от данных обезьяны 357, глубина модуляции вернулась к уровню 2 сут КП, вероятно, вследствие адаптивного торможения.
Обнаруженные изменения чувствительности вестибулярных нейронов к стимуляции отолитов у обеих обезьян в КП близки к наблюдавшимся у шести обезьян в полетах трех биоспутников («Космос-1667», «Космос-1887» и «Космос-2044») и полученным при регистрации нейронов в медиальном вестибулярном ядре во время «лифтной» реакции (Sirota et al., 1987; Kozlovskaya et al., 1989; 1990; Ильин и др.,. 1990; Cohen et al., 2005). В этих исследованиях чувствительность вестибулярных нейронов к отолитовому стимулу в ходе полета постепенно возрастала, достигая максимума к концу первой недели КП, и затем снижалась к концу полета, оставаясь на повышенном уровне или достигая исходного. В некоторых случаях динамика возбудимости нейронов имела многофазный характер: вслед за первичным повышением возбудимости следовало ее снижение и повторное увеличение (Cohen et al., 2005). Можно предполагать, что усиление активности любых нервных клеток, участвующих в различных вестибуло-вызванных реакциях с участием отолитового входа, отражает влияние увеличения возбудимости периферического отдела вестибулярного аппарата или уменьшения активности проприоцептивного входа – предполагаемых факторов, изменяющих возбудимость вестибулярных нейронов второго порядка.
В характере изменений реакций СР у обеих обезьян имеется определенное сходство с изменениями возбудимости вестибулярных нейронов. У обезьяны 357 как чувствительность реакции СР на стимуляцию отолитов, так и вестибулярная возбудимость, увеличивались в начале полета и оставались на повышенном уровне к концу КП, то есть, отсутствию адаптивного торможения вестибулярной возбудимости у обезьяны 357 к концу КП соответствовало сохранение повышенной чувствительности реакции СР на стимуляцию отолитов. При этом значение RRмин сдвигалось в область больших значений. У обезьяны 484 как чувствительность реакции СР на стимуляцию отолитов, так и вестибулярная возбудимость увеличивались в первой половине полета. При этом значение RRмин также сдвигалось в область больших значений. В конце полета ни чувствительность реакции СР на стимуляцию отолитов, ни RRмин у обезьяны 484 не отличались от характерных для контрольной группы животных, а вестибулярная возбудимость не отличалась от исходного уровня в КП, вероятно, вследствие адаптивного торможения.
Тестирование реакции СР на стимуляцию отолитов с помощью четырехштанговых качелей через сутки после КП показало, что у обезьяны 357 реакция интервала R-R на стимуляцию отолитов отсутствовала (рис. 10). У обезьяны 484 через сутки после завершения КП реакция СР была существенно снижена по сравнению с результатами двух тестирований в предполетный период при близких значениях RR0.
 | Рис.10. Амплитуда ответа интервала R-R на стимуляцию отолитов с помощью качелей у обезьяны 357 (кружок) и 484 (заполненные треугольники) после завершения КП в сравнении с предполетными данными обезьяны 484 (открытые треугольники) и линией регрессии по данным 8 животных (рис. 5А). |
Изменения СР в ответ на стимуляцию отолитов через сутки после завершения КП находятся в логическом соответствии с результатами тестирования реакций СР и вестибулярных нейронов на стимуляцию отолитов в конце КП. Смещение RRмин у обезьяны 357 до значения 391±39 мс в 13 сут КП объясняет отсутствие реакции СР при RR0 = 324 мс в послеполетном тестировании. Отсутствие смещения RRмин у обезьяны 484 и предполагаемое адаптивное торможение возбудимости вестибулярных нейронов в конце КП объясняет уменьшение реакции СР у этой обезьяны после завершения КП.
Таким образом, сходный характер изменения чувствительности реакции СР и возбудимости вестибулярных нейронов на стимуляцию отолитов у каждой из двух обезьян в КП и сниженная реакция СР на стимуляцию отолитов в послеполетном тестировании у обезьяны 484 свидетельствуют в пользу предположения (Yates, 1992; Yates, Kerman, 1998) об адаптивном торможении вестибулярных структур в микрогравитации как одной из причин уменьшения участия отолитового входа в компенсаторных реакциях кровообращения на постуральные возмущения в послеполетный период. Вместе с тем отсутствие реакции СР на стимуляцию отолитов у обезьяны 357 в послеполетном тестировании соответствует смещению RRмин в область больших значений исходного RR0 у этого животного в конце КП, показывая, что изменение состояния механизмов регуляции СР вследствие пребывания в микрогравитации также может быть причиной снижения участия отолитового входа в компенсаторных реакциях кровообращения на постуральные возмущения в послеполетный период.
Сопоставление влияний длительной АНОГ и микрогравитации на реакцию СР, вызванную стимуляцией отолитов
Тестирование реакции СР на стимуляцию отолитов у обезьян после завершения АНОГ и КП обнаружило значительное сходство влияния факторов АНОГ и КП на исследуемую реакцию. После завершения АНОГ чувствительность реакции СР на стимуляцию отолитов была достоверно снижена, у двух обезьян реакция отсутствовала. После завершения КП реакция СР на стимуляцию отолитов была уменьшена у одной обезьяны (484) и отсутствовала у другой (357).
Во время КП наряду с увеличением чувствительности реакции СР к стимуляции отолитов у обеих обезьян наблюдалось смещение RRмин в сторону больших исходных значений RR0, что свидетельствует об уменьшении влияния симпато-адреналовой системы на СР. При этом, если у обезьяны 484 к концу КП RRмин вернулся к контрольному уровню, то у обезьяны 357 он оставался повышенным, что, по всей видимости, привело к отсутствию реакции СР на стимуляцию отолитов после завершения КП. У двух обезьян после завершения АНОГ реакция СР также отсутствовала, что свидетельствует о смещении RRмин в сторону больших исходных значений RR0 у этих животных. Так как стимуляция отолитов вызывает торможение парасимпатических влияний на сердце, отсутствие реакции СР у обезьяны 357 после КП и у двух обезьян после АНОГ означает, что сниженное влияние симпато-адреналовой системы на СР у этих животных сопровождалось отсутствием парасимпатических вляний на СР.
Наиболее существенное различие между условиями исследований, проведенных в АНОГ и в КП, заключается в существовании гравитационного воздействия в АНОГ и его практическое отсутствие (микрогравитация) в КП, тогда как перераспределение жидких сред организма в краниальном направлении, опорная разгрузка и гипокинезия присутствуют в обоих исследованиях, хотя и в неодинаковой мере. Сходство реакций СР на стимуляцию отолитов у обезьян после завершения АНОГ и КП свидетельствует о том, что предполагаемое изменение возбудимости рецепторной части отолитовой системы в микрогравитации не является единственным и доминирующим фактором, определяющим динамику возбудимости нейронов вестибулярных ядер, опосредующих влияние стимуляции отолитов на СР, в КП.
Начальное увеличение возбудимости нейронов вестибулярных ядер как во время АНОГ, так и в КП, может быть результатом возникающей опорной разгрузки, дальнейшего снижения мышечного тонуса и сопутствующего уменьшения тормозных влияний проприоцептивных афферентов, конвергирующих на вестибулярных нейронах (Репин, 1981; Крейдич и др., 1982; Kozlovskaya et al., 1985). Последующая нормализация возбудимости вестибулярных нейронов, связанная с их адаптивным торможением и адекватная условиям АНОГ и КП, должна приводить к ослаблению реакций на стимуляцию отолитов после завершения АНОГ и КП.
Снижение эффективности отолитовой стимуляции после завершении АНОГ и в послеполетный период может быть связано также с уменьшенным уровнем парасимпатических влияний на СР, сопутствующим снижению влияния симпато-адреналовой системы на СР, вызванному изменением состояния механизмов регуляции СР как в АНОГ, так и в микрогравитации.
ВЫВОДЫ
- Кратковременная стимуляция отолитов вызывает у обезьян коротколатентное учащение сердечного ритма, связанное с укорочением интервала R-R ЭКГ вследствие торможения парасимпатических влияний на сердце.
- Между величиной уменьшения интервала R-R в ответ на кратковременную стимуляцию отолитов и исходной величиной интервала R-R у обезьян существует прямолинейная зависимость, причем с уменьшением исходной величины интервала R-R ответная реакция снижается. Угол наклона этой прямой характеризует чувствительность реакции интервала R-R к стимуляции отолитов, а точка пересечения с осью абсцисс – минимальное значение исходного интервала R-R, при котором парасимпатические влияния на сердце отсутствуют.
- 30-суточная АНОГ вызывает уменьшение чувствительности реакции интервала R-R на стимуляцию отолитов у обезьян через сутки после завершения АНОГ. Через 7 суток чувствительность реакции не отличается от контрольной.
- Чувствительность реакции интервала R-R на стимуляцию отолитов у обезьян увеличивается в начальный период КП, а ее изменения в течение полета отражают изменения возбудимости вестибулярных нейронов.
- Увеличение чувствительности реакции интервала R-R на стимуляцию отолитов у обезьян в КП сопровождается смещением минимального значения исходного интервала R-R в область больших значений.
- Уменьшение реакции сердечного ритма на стимуляцию отолитов у обезьян в послеполетный период соответствует адаптивному торможению возбудимости вестибулярных нейронов или смещению минимального значения исходного интервала R-R в область больших значений к концу КП.
- В совокупности полученные результаты свидетельствуют о том, что гравитационная разгрузка вызывает уменьшение участия отолитовой системы в быстрых хронотропных реакциях сердца вследствие изменений как вестибулярной возбудимости, так и состояния механизмов регуляции сердечного ритма.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ
- Влияние микрогравитации на отолитовую функцию у обезьян / А.М.Бадаква, Д.В.Залкинд, Н.В.Миллер, С.Н.Рязанский // Тезисы докладов Х1 конференции «Космическая биология и авиакосмическая медицина», Москва, Россия, 22-26 июня 1998 г.-Том 2.-«Слово».-Москва, 1998.--С.400-401.
- Отолитово-вегетативные реакции у бодрствующих обезьян. / А.М.Бадаква, Д.В.Залкинд, Н.В.Миллер, С.Н.Рязанский // Авиакосм. и экологич. мед.-1999.-т.33.-№3.- С.42-47.
- Исследование отолитово-вегетативных ответов в экспериментах на обезьянах Macaca mulatta / А.М.Бадаква, Н.В.Миллер // Матер. Российск. Конф. «Организм и окружающая среда», Москва, 26-29 сентября 2000 г. «Слово», Москва.-2000.-Том 2.- С.199-200.
- Влияние антиортостатической гипокинезии и ее сочетания с сеансами ускорения +Gz на регуляцию сердечного ритма у обезьян. / А.М.Бадаква, Н.В.Миллер, С.Н.Рязанский // Авиакосм. и экологич. мед.-2001.-т.35.-№1.-С.32-37.
- Cardiac rhythm responses to otolith stimulation of monkeys in microgravity. / A.M.Badakva, D.V.Zalkind., N.V.Miller // J. Grav. Physiol.-2000.-Vol.7.-N1.-P.S107-S111.
- Head pitch movement and vestibular neuronal activity in response to otolith stimulation of monkeys in space | A.M.Badakva, D.V.Zalkind., N.V.Miller, S.N. Riazanski // J. Grav. Physiol.-2000.-Vol.7.-N1.-P.S99-S105.
- The effect of +Gz acceleration on monkey cardiac rhythm autonomic regulation during head-down bedrest / A.M.Badakva, N.V.Miller, S.N.Riazanski // Abstract volume of 13-th IAA “Humans in Space” Symposium, 20-26 May, 2000. Santorini, Greece. P.162.
- Effect of Spaceflight and Head-Down Bedrest on Autonomic Control of Cardiac Rhythm in Monkeys / A.Badakva, N.Miller // In: 8th European Symposium on Life Sciences Research in Space. 23rd Annual International Gravitational Physiology Meeting. 2-7 June 2002, Karolinska Institutet, Stockholm, Sweden. Abstracts SP-501, p.78.