

Валовая первичная продукция и биомасса в муссонном тропическом лесу южного вьетнама
На правах рукописи
НОВИЧОНОК
Артём Олегович
ВАЛОВАЯ ПЕРВИЧНАЯ ПРОДУКЦИЯ И БИОМАССА В МУССОННОМ ТРОПИЧЕСКОМ ЛЕСУ ЮЖНОГО ВЬЕТНАМА
03.02.08 – экология
Автореферат диссертации на соискание ученой степени кандидата
биологических наук
Петрозаводск
2013
Работа выполнена в Федеральном государственном бюджетном образовательном учреждении высшего профессионального образования Петрозаводский государственный университет (кафедра ботаники и физиологии растений, эколого-биологический факультет)
Научный руководитель доктор биологических наук
профессор
Марковская Евгения Фёдоровна
Петрозаводский государственный
университет
Официальные оппоненты: Кузнецов Олег Леонидович
доктор биологических наук
Федеральное государственное бюджетное
учреждение науки Институт биологии
Карельского научного центра РАН
старший научный сотрудник, заведующий лабораторией болотных экосистем
Ольчев Александр Валентинович
кандидат географических наук
Федеральное государственное бюджетное учреждение науки Институт проблем экологии и эволюции им. А.Н. Северцова
Российской академии наук (ИПЭЭ РАН)
старший научный сотрудник лаборатрии биогеоценологии и исторической экологии им. В.Н. Сукачева
Ведущая организация: Институт биологии Коми научного центра Уральского отделения Российской академии наук
Защита состоится «27» ноября 2013 г. в 14 часов на заседании диссертационного совета Д 212.190.01 при Петрозаводском государственном университете по адресу: 185910, Республика Карелия, г. Петрозаводск, пр. Ленина, 33, эколого-биологический факультет, тел., факс: 8(8142)763864.
С диссертацией можно ознакомиться в библиотеке Петрозаводского государственного университета, с авторефератом - на сайтах http://vak.ed.gov.ru/ и www.petrsu.ru.
Автореферат разослан: « 27 » октября 2013 г.

Ученый секретарь диссертационного совета
кандидат биологических наук Дзюбук И.М.
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность исследования. Вопросы, связанные с углеродным циклом в естественных экосистемах, остаются одними из наиболее важных в современных экологических исследованиях в связи с глобальными изменениями климата и окружающей среды (Бабкова, 1987; Исаев и др., 1992; МГЭИК, 2007; Усольцев, 2010).
Тропические леса являются самыми продуктивными сообществами нашей планеты, поэтому их участие в глобальном обмене углерода между биосферой и атмосферой велико. Однако оценка роли тропических лесов в глобальном цикле углерода по-прежнему является неоднозначной (Malhi and Grace, 2012). Значительное разнообразие лесов тропического региона приводит к необходимости детальных региональных исследований.
Листопадные и полулистопадные тропические леса относят к одним из самых уязвимых биомов планеты. Они расположены в регионах с относительно сухим сезонным климатом. На протяжении нескольких тысячелетий муссонные леса находятся под интенсивным воздействием хозяйственной деятельности человека. В тропической зоне Юго-Восточной Азии лесистость территории изменятся от 74% (Новая Гвинея) до 28-42% (Вьетнам, Камбоджа) (Структура и функции…, 2011). Во Вьетнаме до Второй Индокитайской войны лесистость территории составляла около 43% (Maurand, 1943). В период войны 1950-1970 гг. леса сильно пострадали от применения дефолиантов, а после войны от сведения лесов под сельскохозяйственное использование. К 1995 г. под лесной растительностью находилось только 28% территории страны. В настоящее время леса Вьетнама имеют мозаичное распространение и, как правило, угнетённое состояние. Сходная картина наблюдается и в других развивающихся странах Юго-Восточной Азии. В настоящее время остро стоит вопрос планирования рационального использования природных ресурсов Юго-Восточной Азии, гармоничного сосуществования человека и природы. Для его решения необходимы комплексные биологические исследования в регионе.
Цель и задачи работы. Цель работы состояла в оценке запасов углерода, валовой первичной продукции (GPP) тропического муссонного леса южного Вьетнама и в оценке влияния внешних гидрометерологических переменных на суточную и сезонную динамику GPP.
Для реализации поставленной цели были решены следующие задачи:
1. Исследование таксационных характеристик репрезентативных для тропического муссонного леса юга Вьетнама типов лесных сообществ;
2. Определение запасов суммарной фитомассы в репрезентативных древостоях тропического муссонного леса на юге Вьетнама;
3. Обеспечение комплексных высокочастотных режимных измерений потоков диоксида углерода на основе методики турбулентных пульсаций в сочетании с актинометрическими и метеорологическими наблюдениями;
4. Оценка суточной и сезонной динамики GPP, её зависимости от гидрометеорологических переменных; оценка кумулятивных значений GPP для различных сезонов и года в целом. Выявление особенностей временной динамики GPP, характерных для экосистемы тропического муссонного леса.
Научная новизна. Впервые на основе непрерывных круглогодичных высокочастотных наблюдений получены данные о валовой первичной продукции равнинного муссонного тропического леса южного Вьетнама Выделены особенности суточной и сезонной динамики GPP и проанализирована связь GPP муссонного тропического леса с факторами внешней среды. На основе лесотаксационных методов получены оценки запаса углерода в основных пулах углерода некоторых лесных сообществ в муссонном тропическом лесу южного Вьетнама.
Теоретическая значимость работы. Проведенное исследование расширило существующие на настоящий день представления о запасах углерода в основных пулах и продуктивности сезонно-влажных тропических лесов. Впервые получены данные по GPP муссонных лесов южного Вьетнама в суточной и сезонной динамике с использованием пульсационного метода исследований.
Практическая значимость работы. Полученные данные могут быть использованы при формировании экологической политики стран Юго-Восточной Азии, а также для разработки проектов рационального использования и сохранения лесных сообществ региона.
Апробация работы. Основные положения диссертации были представлены на международном Симпозиуме Европейского геофизического сообщества (Австрия, Вена, 2013 гг.); Американского геофизического сообщества (США, Сан-Франциско, 2012); на научном семинаре Института Прикладной механики и информатики Вьетнамской Академии Наук и технологий, посвященном Инициативному проекту Asia Geo Grid и развитию мониторинга за потоками парниковых газов на территории Вьетнама (Вьетнам, Хошимин, 2013); на совместном семинаре Института Тропической Биологии Вьетнамской Академии Наук и Технологий и Центра по исследованию Глобальных изменений Чешской Академии Наук «Обзор состояния пульсационных измерений во Вьетнаме, включая новые данные со станции в национальном парке Кат Тьен. Фиксация и баланс углерода и влияние глобальных изменений» (Хошимин, 2012); на семинарах Совместного Российско-Вьетнамского научно-исследовательского и технологического тропического центра (Хошимин, 2012, 2013); на международной конференции сообщества AsiaFlux (Сеул, 2013); на II Международной научно-практической конференции «Теоретические и практические вопросы развития научной мысли в современном мире» (г. Уфа, 2013).
Структура и объем диссертации. Диссертация состоит из введения, 4 глав, выводов и списка литературы. Работа изложена на 156 страницах текста, содержит 9 таблиц, 43 рисунка. Список литературы включает в себя 205 источников, из них – 180 зарубежных.
Благодарности. Исследования были выполнены в рамках плановой темы НИР Совместного Российско-Вьетнамского научно-исследовательского и технологического тропического центра. Автор выражает благодарность руководству и сотрудникам Тропического Центра, без содействия которых было бы невозможно проведение исследований на территории Вьетнама. Автор благодарен дирекции национального парка Кат Тьен, на территории которого выполнены все экспериментальные исследования. Неоценимую помощь в сборе экспериментального материала оказали Новичонок Е.В, Дещеревская О.А., Авилов В.К. Автор выражает благодарность научному руководителю Марковской Е.Ф., руководителю темы НИР Курбатовой Ю.А. за всестороннюю помощь и консультации при написании работы.
Исследования были поддержаны РФФИ, грант 12-04-31234 мол_а «Эколого-физиологические адаптации растений на нарушенных территориях».
КРАТКОЕ СОДЕРЖАНИЕ РАБОТЫ
Глава 1. Углеродный баланс лесных экосистем и продуктивность сезонно-влажных тропических лесов Юго-Восточной Азии: литературный обзор
В главе на основе литературных данных представлены особенности углеродного цикла естественных экосистем, введена терминология, используемая при описании продуктивности. Даны определения пулов углерода, валовой первичной продукции (GPP); чистой первичной продукции (NPP); чистой экосистемной продукции (NEP) (Базилевич и Титлянова, 2008; Горшков, 2002; Пулы и потоки…, 2007).
Показана роль тропических лесов (как источника и стока атмосферного углерода) в цикле углерода на планете в условиях нарастающей антропогенной нагрузки. Представлен краткий обзор исследований продуктивности сезонно-влажных тропических лесов, выполненных с использованием различных подходов – инвентаризационного, дистанционного зондирования (Blanc et al., 2000; Iverson et al., 1993; Schmid, 1974; Trung, 1966). Дан краткий обзор результатов пульсационных исследований, проведённых в регионах, географически близких к Южному Вьетнаму – в Таиланде (участки Sakaerat и Mae Klong) и Малайзии (леса Pasoh и Lambir Hills) (Saigusa et al., 2008).
Глава 2. Объекты и методы исследований
Исследование было выполнено в национальном парке Кат Тьен (11°21'-11°48' с. ш. 107°10'-107°34' в. д.)
Кат Тьен – самый крупный национальный парк на территории Южного Вьетнама, площадь парка 71 920 га. Значительная часть территории парка (около 90%) представлена муссонными тропическими лесами, в целом уровень их антропогенной нарушенности невелик.
Структура лесных сообществ парка очень гетерогенна с подавляющим преобладанием вторичных лесов. Значительные площади покрывают легерстремовые леса, диптерокарповые леса, молодые регенерирующие леса, леса с бамбуковыми примесями и бамбуковые сообщества (Структура и функции…, 2011).
Полевые рекогнесцировочные таксационные описания были выполнены в июле-сентябре 2011 г на территории в непосредственной близости от вышки для пульсационных измерений. Для работы была использована карта инвентаризации лесов национального парка Кат Тьен (1999). На основании анализа этой карты были выбраны три основных типа лесных сообществ, характерных для изучаемого участка: первичный лагерстремовый лес (заложено 3 пробные площади, ПП), вторичный естественно восстанавливающийся (заложено 5ПП) и вторичный наиболее нарушенный – «редколесье» (заложено 2 ПП).
Таксационные описания были выполнены по специально разработанной для тропических лесов методике, с использованием расчетных формул для пересчёта диаметра стволов деревьев в биомассу (Pearson et. al., 2005). Для проведения исследований был использован метод вложенных пробных площадей. На выбранных участках основных типов лесных сообществ были заложены квадраты: для измерения деревьев с диаметром более 20 см – 3535 м, для деревьев с диаметром 5-20 см – 1010 м и для деревьев с диаметром менее 5 см – 1.51.5 м. Биомасса каждого дерева рассчитывалась на основании общего аллометрического уравнения для влажных тропиков (Brown, 1997). Биомасса опада измерялась на 5 пробных площадках, выбранных случайным образом, площадью 0,30,3 м на каждой ПП. Для определения биомассы лиан, был использован расчетный метод, ранее разработанный для экваториального леса Венесуэлы (Putz, 1983). Биомасса корней также определялась расчетным методом по формулам для тропического региона, основанным на биомассе надземных органов (Pearson et. al., 2005).
Потоки углекислого газа измерялись с использованием метода пульсационных наблюдений. Измерительная аппаратура была закреплена над пологом леса на металлической вышке на высоте 51 м. Концентрацию СО2 определяли с использованием инфракрасного газоанализатора открытого типа LI-7500 (LI-COR Inc., США). Кроме того, концентрация углекислого газа измерялась на семи разных высотах (от 0,3 м до 30 м) с использованием инфракрасного газоанализатора закрытого типа LI-820. Скорость ветра фиксировалась звуковым анемометром CSAT3 (Campbell Scientific Inc., США). Регистрация данных производилось с частотой 10 Гц. Одновременно с потоками углекислого газа также фиксировались гидрометеорологические параметры атмосферы (восходящая и нисходящая солнечная и длинноволновая радиация, температура воздуха, влажность воздуха, фотосинтетически активная радиация, скорость и направление ветра, атмосферное давление, суммарные осадки и гидротермические параметры почвы (профиль температуры и влажности почвы).
Энергоснабжение измерительного комплекса осуществлялось за счет солнечной энергии.
Потоки CO2 рассчитывались как ковариация между пульсациями вертикальной скорости ветра и концентрации CO2, измеренными ультразвуковым анемометром и инфракрасным газоанализатором. Результатом работы измерительного комплекса являются высокочастотные круглогодичные оценки баланса СО2 между пологом тропического леса и атмосферой (чистый экосистемный обмен, NEE).
Потоки СО2 рассчитывались в программе EddyPro 4.1.0 (LI-COR Inc., США). Разделение потоков на валовую первичную продукцию и экосистемное дыхание осуществлялось в онлайн-приложении[1], созданном командой Института Биогеохимии Макса Планка (Германия) на основе описанных методов разделения потоков, предложенных в работах Reichstein et al. (2005) и Lasslop et al. (2010).
Глава 3. Результаты исследований
3.1. Исследования запасов фитомассы на типичном участке муссонного тропического леса в национальном парке Кат Тьен
Лес с доминированием Lagerstroemia calyculata Kurz. в верхнем подъярусе из 4-5 древесных является достаточно распространённым на изучаемой территории. В верхнем подъярусе данного типа леса встречаются и другие виды: Afzelia xylocarpa (Kurz.) Craib, Tetrameles nudiflora R. Br. Этот подъярус имеет разомкнутую структуру, деревья лагерстремии имеют высоту 30-40 метров, причём крона начинается на высоте не менее 15 метров. Второй подъярус этого типа леса сомкнутый, имеет высоту около 20 метров или чуть выше; преобладающими видами являются Acronychia laurifolia (L.) Miq., Dalbergia mammosa Pierre, Peltophorum dasyrrachis Pierre, Markhamia stipulate (Dop) Sant, Pterocymbium tinctorium (Blanco) Merr, Pterospermum sp., Rothmannia eucodon (K. Schum.) Bremek., Xylopia vielana Pierre ex Fin. & Gagn., Polyalthia sp., Cinnamomum sp., Phoebe sp., Diospyros spp., Knema sp. (Кузнецов, 2012). Подрост доминантного вида (Lagerstroemia calyculata) не встречается под пологом этого типа леса, но широко распространён в лесных окнах, по обочинам лесных дорог, на вывалах и прогалинах. Вторичный лагерстремовый лес выделяется множественным присутствием крупных стволов Lagerstroemia calyculata. Деревья-доминанты, как правило, имеют диаметр стволов не менее 70 см (самое большое измеренное дерево имело диаметр около трёх метров, см. рис. 1, табл. 1); которые формируют очень значительную биомассу данного типа лесного сообщества в целом[2]. Количество больших (более 25 см в диаметре) деревьев составляет 55-65 стволов на гектар. На месте погибших или упавших деревьев-доминантов временно формируются окна, в которых бурно развивается невысокая растительность, в частности, ротанговая пальма (Calamus spp), а также многочисленные виды лиан. Здесь также иногда можно встретить молодые деревья Lagerstroemia calyculata.
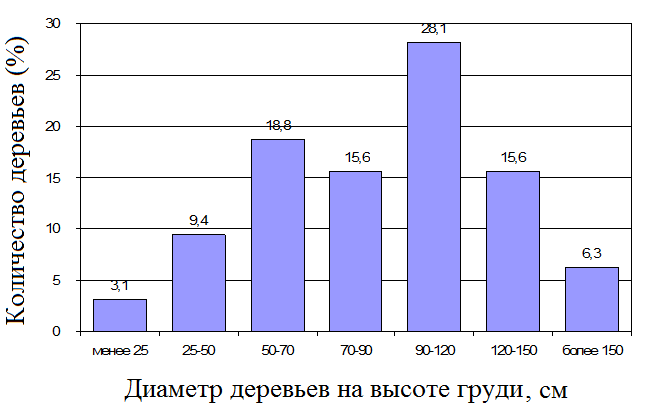
Рис. 1. Распределение деревьев Lagerstroemia calyculata в зрелом лагерстремовом лесу по диаметру ствола. Учтены только деревья с диаметрами более 5 см.
Таблица 1.
Распределение биомассы в Лагерстремовом лесу.
| Составная часть | Биомасса (т/га) на 1-й ПП, | Биомасса т/га на 2-й ПП, | Биомасса т/га на 3-й ПП, | Средняя биомасса, т/га |
| Деревья с диаметром более 20 см (биомасса надземной части) | 631,3 | 510 | 450,6 | 530,6 |
| Деревья с диаметром 5-20 см (биомасса надземной части) | 53,1 | 21,5 | 42,9 | 39,2 |
| Опад | 19,1 | 3,1 | 5,2 | 4,2 |
| Подрост | 7,0 | 6,1 | 4,0 | 6,0 |
| Лианы | 32,0 | 24,3 | 22,7 | 26,3 |
| Мёртвая древесина | 4,5 | 3,3 | 5,1 | 4,3 |
| Подземная биомасса | 112,1 | 89,7 | 83,9 | 95,2 |
| Сумма | 859,0 | 658,0 | 615,3 | 710,8 |
Биомасса деревьев с диаметром более 20 см (подавляющий вклад в которую вносит биомасса доминантного вида) составляет 530,6 т/га (74,6% биомассы сообщества в целом); доля деревьев с диаметрами 5-20 см по биомассе – 5,5% (39,2 т/га). Биомасса деревьев с диаметрами стволов менее 5 см имеет наименьшие значение – 6,0 т/га или 0,8% от общей биомассы.
В естественно восстанавливающемся лесу, как и в лагерстремовом, можно выделить 4-5 древесных подъярусов. Самый верхний уровень занимают флаговые деревья, среди которых Lagerstroemia calyculata L., Afzelia xylocarpa (Kurz.) Craib., Tetrameles nudiflora R. Br., Haldina (syn. Adina) cordifolia (Roxb.) Ridsd., Hopea odorata Roxb. и множество других. Деревья здесь, как правило, довольно тонкие (средний диаметр крупных деревьев [с диаметром более 20 см] около 30 см в отличие от лагерстремового леса, где этот показатель больше 100 см). Естественно восстанавливающийся лес имеет значительно меньшую общую растительную биомассу (~410 т/га). Однако доля больших деревьев (с диаметром более 20 см) также довольно велика – 69,0% по биомассе, доля средних деревьев (с диаметром 5-20 см) по массе здесь значительно выше в процентном отношении, чем в лагерстремовом лесу (14,8% – 56,1 т/га). Средняя высота полога примерно такая же, как и в лагерстремовом лесу (~30 метров). Полог довольно сомкнутый.
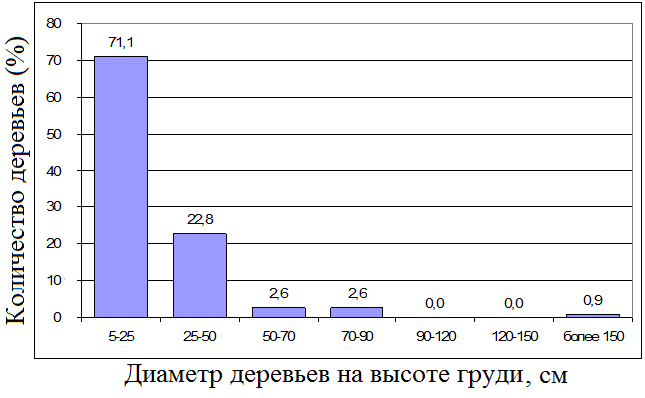
Рис. 2. Распределение деревьев в естественно восстанавливающемся лесу по диаметрам. По оси х – DBH (см), по оси у – процент количества деревьев. Учтены только деревья с диаметрами более 5 см.
Согласно данным по инвентаризации лесов национального парка Кат Тьен, именно этот тип лесного сообщества преобладает на исследуемой территории.
«Редколесье»[3] – наиболее угнетённое сообщество, представленное во многом оконными группировками, а также недоразвитыми участками леса. Крупные деревья (более 20 см в диаметре) здесь лишь единичны и разрозненны, представлены теми же видами, которые упоминались в описаниях предыдущих типов лесных сообществ. Крупные деревья дают лишь 27% (47,7 т/га) общей биомассы сообщества. Деревья с диаметром 5-20 см составляют 35,1% (62,0 т/га) от общей биомассы. Значительный вклад в биомассу вносят деревья с диаметром менее 5 см (6,1% в отличие от 0,8% и 1,8% у лагерстремового и естественно восстанавливающегося лесов соответственно.
Исследование вклада листового опада в общую биомассу сообществ показало, что он составил соответственно 4,2 (0,6%), 5,6 (1,4%) и 3,1 т/га (1,8%) для лагерстремового леса, восстанавливающегося сообщества и «редколесья». При анализе динамики опада обязательно нужно учитывать, что его биомасса во влажный сезон минимальна, а пик приходится на период активного листопада (сухой сезон). Биомасса мёртвых стволов деревьев не имеет значимых различий в разных типах леса. Она составила соответственно 4,3 т/га (0,6% об общей биомассы сообщества) в лагерстремовом лесу, 3,3 т/га (1,0%) в естественно восстанавливающемся лесу и 2,4 т/га (1,4%) т/га в редколесье.
3.2. Валовая первичная продукция углерода (GPP) в суточной и сезонной динамике
3.2.1. Метеорологические условия в 2012 году
Зависимость продукционного процесса от факторов внешней среды в настоящее время подтверждена многочисленными экспериментальными исследованиями и модельными расчетами (Одум, 1986; Молчанов, 2007; Усольцев, 2005; Field, 1998).
Температурно-влажностный режим исследуемой территории определяется муссонным режимом циркуляции и делится на два сезона – сухой и влажный.
Среднегодовая температура воздуха на высоте 30 м в течение года была равна 25.9 °С, самыми тёплыми месяцами являлись март, апрель и май (26.7 – 26.9 °С), а самыми холодными – январь, сентябрь и октябрь (24.8 – 24.9 °С). Среднесуточная, максимальная и минимальная температуры воздуха на высоте 30 м изменялись незначительно в течение года; диапазон варьирования среднесуточных значений составил от 24.5 (в январе) до 27.5 °С (в марте). Максимальная температура в 2012 г., +35.3 °С, была зафиксирована 17 марта в 15 ч, минимальная, +18.7 °С (20 февраля, 6 ч 30 мин).
Рис. 3 показывает средний суточный ход температуры воздуха на высоте 30 м в марте и сентябре 2012 года. Март – самый жаркий и один из самых сухих месяцев года, в то время как сентябрь – один из самых влажных месяцев, с наибольшим количеством осадков. В марте после восхода Солнца увеличение температуры воздуха до максимума (14-15 ч) и снижение её во второй половине дня происходит намного быстрее, чем в сентябре. Средние максимальные значения значительно различаются – 33°С в марте против 28 °С в сентябре; общая амплитуда колебаний температуры воздуха в течение суток в сентябре значительно меньше, чем в марте.
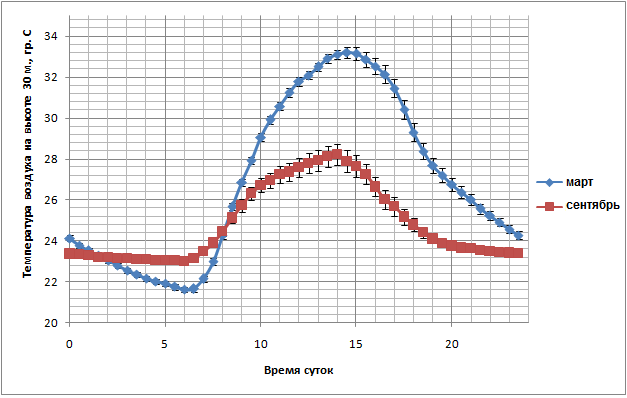
Рис. 3. Средний суточный ход температуры воздуха в марте и сентябре 2012 г. на высоте 30 м.
Суточная динамика температуры воздуха внутри полога в марте и сентябре по своим закономерностям сходны с ходом температуры воздуха на высоте 30 м.
Режим увлажнения. Сезонная динамика суммарных осадков представлена на Рис. 4. В течение 2012 г. выпало 2621 мм осадков, причём 2476 мм (94.5%) пришлось на период с апреля по октябрь (влажный сезон). Сухой сезон характеризовался несколькими необильными дождями. Самый влажный месяц – сентябрь (671 мм осадков), самый сухой – декабрь (3 мм).
В течение влажного сезона осадки выпадали почти каждый день, в то время как в сухой сезон перерывы между отдельными периодами выпадения незначительных осадков составляли 1-2 недели.
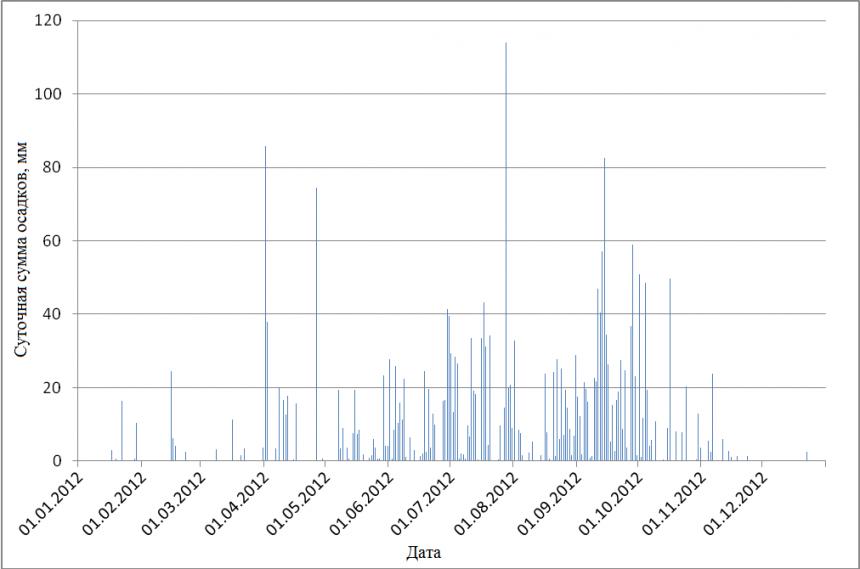
Рис. 4. Годовая динамика суммарных осадков (мм/сут) в национальном парке Кат Тьен в 2012 году.
Суточная динамика дефицита водяного пара в воздухе (VPD). Максимальные значения дефицита водяного пара (которые в течение всего года наблюдались в 14-16 ч) в марте в два с половиной раза больше (30,5 гПа), чем в сентябре (12,6 гПа). В марте минимальное среднее значение VPD (1.4 гПа) наступало в момент восхода Солнца – в 6 ч по местному времени, после чего VPD линейно возрастал до своего максимума. В сентябре это увеличение отмечалось после восхода солнца с задержкой в 2 ч.
Радиационный режим. Восход и закат Солнца на территории южного Вьетнама в течение всего года наблюдается примерно в одно и то же время – в 6 и 18 часов по местному времени соответственно; продолжительность дня примерно равна продолжительности ночи.
Величина радиационного баланса варьирует в течение года в достаточно узком диапазоне: от 335 (сентябрь) до 473 (октябрь) МДж/м2месяц, а месячные суммы фотосинтетически активной радиации (ФАР) от 646 (сентябрь) до 1278 (март) моль/м2месяц. Март и сентябрь – месяцы равноденствий, и количество поступающей солнечной радиации было одинаково в течение обоих месяцев. Следовательно, различия в уровне ФАР на высоте 50 м. обусловлены исключительно различиями в режиме облачности.
Температурно-влажностный режим почвы. Влажность почвы в течение года несколько изменялась на глубинах 5-50 см, но даже в разные сезоны была сравнима, увеличиваясь после периодов выпадения осадков. На большей глубине характер изменения влажности почвы отличался большей стабильностью и меньшей амплитудой последождевых скачков. Минимальные значения влажности почвы (около 20% на всех глубинах) были отмечены в марте и декабре, максимальные значения – в сентябре (от 38% до 41% на глубинах от 5 до 50 см соответственно). Максимальные значения влажности почвы наблюдались в конце сентября – начале октября после мощных и продолжительных ливней (влажность кратковременно достигала значений до 50 % на глубинах 5-10 см).
Температура почвы в течение года на разных глубинах различалась незначительно. Максимальные значения на всех глубинах были отмечены в мае (27 °С), а минимальные – в январе (24.2, 24.5 и 24.7 °С на глубине 5, 30 и 50 см соответственно).
3.2.2. Суточная динамика GPP
Общая картина суточного хода GPP в течение всего года была примерно одинаковой. На рис. 5 показан средний ход суточной динамики валовой первичной продукции в разные месяцы 2012 года (с января по декабрь), рассчитанные двумя различными методами разделения потоков (Reichstein et al., 2005; Lasslop et al., 2010). Результаты за февраль, полученные с использованием световых кривых (метод Lasslop et al., 2010), отсутствуют из-за нехватки исходных данных.
Вклад сториджа. Сторидж (от англ. storage, т.е. запас), или баланс углекислого газа в пологе растительности, позволяет учесть процессы углеродного обмена, происходящие внутри самого сообщества, между уровнем земли и уровнем установки приборов. Сторидж рассчитывался на основании измерений, полученных приборами CO2-профиля – системы воздухозаборов, установленных на разных высотах (0,3 м, 1 м, 2 м, 4,9 м, 10,2 м, 19,2 м, 28,2 м, 46,2 м) с целью измерения концентрации углекислого газа в воздухе.
Для расчёта использовалась формула, опубликованная Jarvis с коллегами (1997).
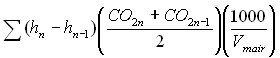 (1).
(1).
Здесь n 1, … n – номера уровней, на которых выполняются измерения концентрации СО2, h – высота измерения концентрации СО2, Vmair - молярный объем воздуха, 22,4 дм3 моль-1.
На рис. 6 показан усреднённый за 2012 г. суточный ход сториджа. Отрицательные значения означают поглощение углерода сообществом, а положительные – его выделение. Большую часть суток сторидж принимает небольшие положительные значения. Пик сториджа приходится на 9-10 ч по местному времени.
Скорость турбулентного перемешивания нижних слоёв атмосферы в среднем значительно ниже ночью, чем днём. В то же время, ночью фотосинтетическая активность полностью отсутствует, а процессы дыхания продолжаются. В результате углекислый газ накапливается под пологом, распределяясь стратифицированно и лишь незначительно перемещаясь в вертикальном направлении. С восходом солнца уровень турбулентого перемешивания резко возрастает, в результате чего накопленный под пологом за ночь углекислый газ высвобождается из сообщества в атмосферу, а также является «сырьём» для утренней фотосинтетической активности. Концентрация углекислого газа под пологом значительно понижается и, как следствие – резкий утренний пик сториджа. В эти часы поток сториджа выше -15 мкмоль/м2с, нередко по модулю больше, чем -20 мкмоль/м2с. Концентрация CO2 под пологом леса в эти часы снижалась на 50-100 ppm относительно максимальных значений. Далее, по мере приближения к полудню и после него полог оставался хорошо «проветриваемым», в результате чего значения сториджа постепенно приближались к нулевым, снова возрастая перед закатом, когда экосистемное дыхание в пологе начинает превалировать над фотосинтезом. Анализ показывает, что не существует значимых отличий в суточном ходе сториджа в разные сезоны года.
Данный характер суточного хода сториджа является типичным для тропических лесов, в т.ч. амазонских (Goulden et al., 1996a, b; Grace et al., 1996), в которых сильно выражен утренний негативный пик сториджа.
В ходе анализа полученных данных сторидж учитывался при определении суммарного потока углекислого газа, который в дальнейшем был разделён на валовую первичную продукцию экосистемы GPP и экосистемное дыхание Reco.
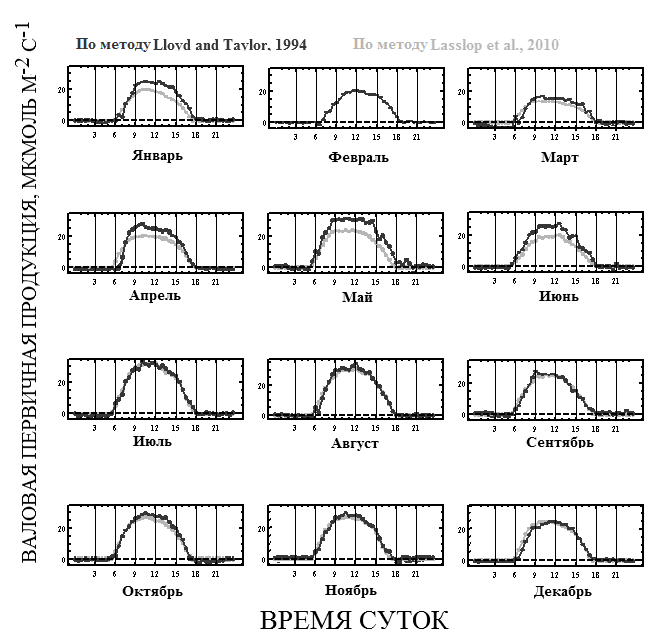
Рис. 5. Средний суточный ход валовой первичной продукции (GPP) по месяцам (мкмоль/м2с), определённой двумя методами. Тёмные линии – классический метод Ллойда-Тэйлора (Lloyd and Taylor, 1994), светлые линии – метод, основанный на исследовании световых кривых (Lasslop et al., 2010).
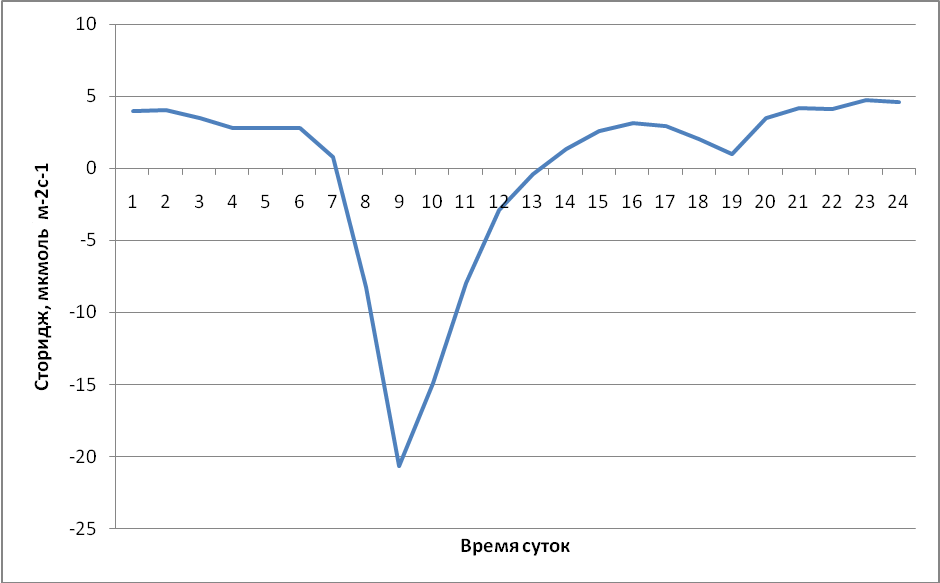
Рис. 6. Средний годовой суточный ход сториджа в 2012 году в муссонном тропическом лесу национального парка Кат Тьен
Роль внешних факторов.
1. Доступность ФАР – один из главных факторов, определяющих интенсивность фотосинтеза и, следовательно, непосредственно отвечающий за величину GPP. Для исследования зависимости получасовых значений GPP от получасовых значений ФАР использовали модифицированную функцию Микаэлиса-Ментен (Йоргенсен, 1985).
P(I(x,t)) = a1 + b1*I(x,t)/(I(x,t) + b2) (2)
Здесь P(I(x,t)) – продуктивность при данном I (ФАР), a1 – параметр, характеризующий продуктивность при I = 0 (т.к. мы изучаем только GPP, этот параметр равен 0); b1 – величина продуктивности при насыщающих значениях ФАР (Pmax); b2 (константа насыщения) – параметр, равный величине I, при которой P = 0.5Pmax, и является характеристикой скорости насыщения световой кривой; t – время.
Использованное уравнение не учитывает влияния на GPP изменяющейся температуры воздуха.
С использованием модифицированной функции Микаэлиса-Ментен выяснено, что насыщающие значения (b2) GPP в марте значительно меньше июльских (23,8 мкмоль/м2с против 40,4 мкмоль/м2с1), при этом константа насыщения (b1) в марте составляет 296 мкмоль/м2с1 против 427 мкмоль/м2с1 в июле (при этом и абсолютные значения продуктивности в июле значительно выше мартовских). Разброс значений GPP относительно среднего суточного хода в марте значительно превышает июльский (R2 = 0,31 и 0,61 соответственно).
GPP и, следовательно, скорость фотосинтеза при одном и том же уровне ФАР была выше в утренние часы относительно постполуденных (рис. 7). Эта полуденная депрессия фотосинтеза была ярко выражена не только в месяцы сухого сезона, но и в течение влажного периода. Например, в марте в утренние часы усреднённая GPP при ФАР 1000 мкмоль/м2с составляла 19 мкмоль/м2с, в то время как в постополуденные время снижалась на 34% и составляла только 12,5 мкмоль/м2с при том же уровне ФАР. В июле в постполуденное время GPP снижалась в среднем на 36%, значения GPP составили 27,5 мкмоль/м2с и 17,5 мкмоль/м2с до и после полудня соответственно. Таким образом, как в сухой, так и во влажный сезон картина снижения GPP в постполуденный период времени была сходна.
2. Температура воздуха. Исследована зависимость GPP от температуры воздуха на высоте 30 м. GPP достоверно коррелирует с температурой воздуха как в сухой, так и во влажный сезоны (R2 = 0,22 и 0,27 соответственно), особенно сильная зависимость наблюдалась в утренние, предполуденные часы, когда R2 может достигать 0,5. При этом с ростом температуры воздуха растёт и интенсивность фотосинтеза, но при определённых значениях температуры воздуха наблюдается ингибирование фотосинтетической активности. Температурное ингибирование при 30 °С отмечено в течение всего года.
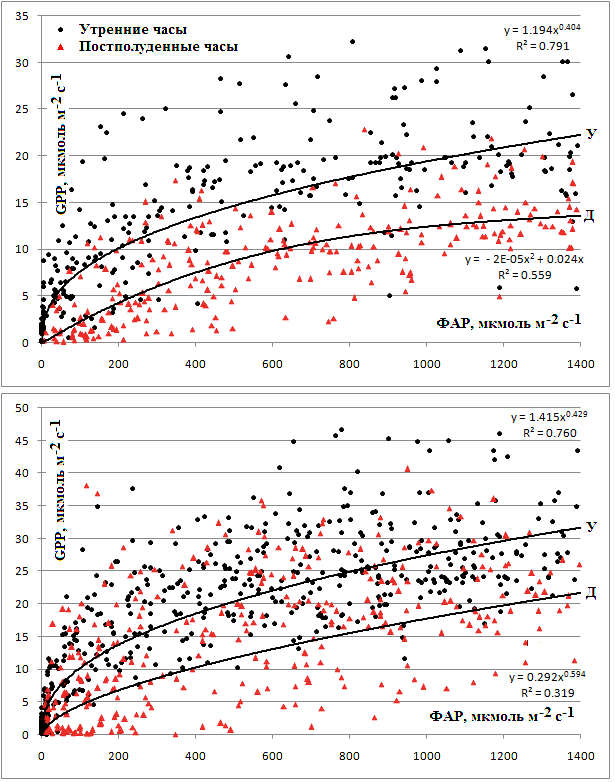
Рис. 7. Полуденная депрессия фотосинтеза в марте (сверху) и июле (снизу). Показана зависимость GPP (в мкмоль/м2с) от ФАР (мкмоль/м2с). Регрессионный анализ проведён отдельно для утреннего периода (до 11 ч местного времени, У) и для постполуденного периода (после 13 ч местного времени, Д)
3. Дефицит водяных паров в воздухе (VPD). Анализ зависимости GPP от дефицита водяных паров в воздухе показал явное ингибирование фотосинтетической активности при высоких значениях VPD как в сухой (март), так и во влажный (июль) сезоны. В марте ингибирование начиналось при VPD 12 гПа, в июле – при VPD 8-10 гПа.
3.2.3. Сезонная динамика GPP
Изменение GPP по месяцам показано на рис. 8.
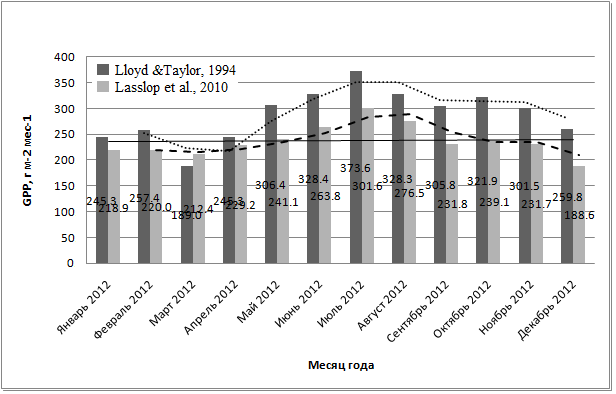
Рис. 8. Суммарная первичная валовая продукции углерода (в г/м2 в месяц) в разные месяцы года, определённая с использованием двух различных способов разделения потоков
Изменение валовой продукции в течение года было небольшим – от 219 г/м2 за месяц в марте до 374 г/м2 в месяц в июле. Наблюдается тенденция некоторого увеличения GPP в течение влажных месяцев, после чего, с резким падением количества осадков, величина продукции постепенно снижается, достигая типичных для сухого сезона относительно небольших значений.
Роль внешних факторов.
1. ФАР. Выявлена небольшая (R2=0,18) достоверная зависимость суточных сумм GPP от суточных сумм ФАР в сезонной динамике (рис. 9). GPP выходит на плато уже при относительно небольшом ФАР (~25 моль/м2сутки). Подобные суммы достигаются почти ежедневно даже в течение влажного сезона (рис. 10).
2. Влажность почвы на глубине 50 см. Не удалось обнаружить значимой корреляции между GPP и влажностью почвы на глубине 50 см.
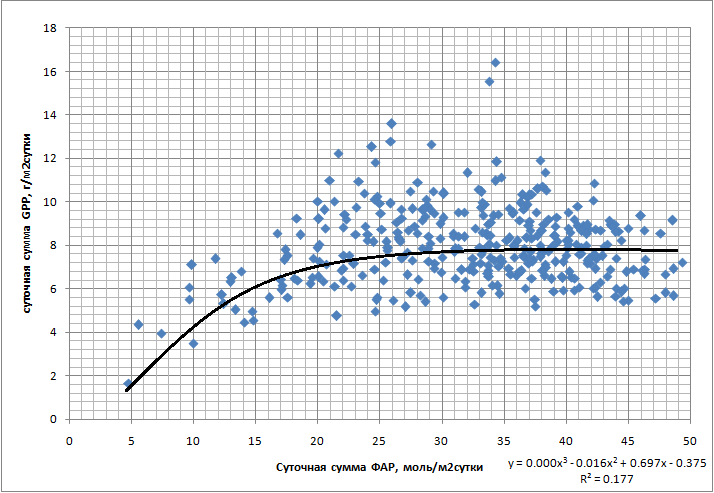
Рис. 9. Обобщённая зависимость суточных сумм GPP (гС/м2сутки) от суточных сумм ФАР (моль/м2сутки)
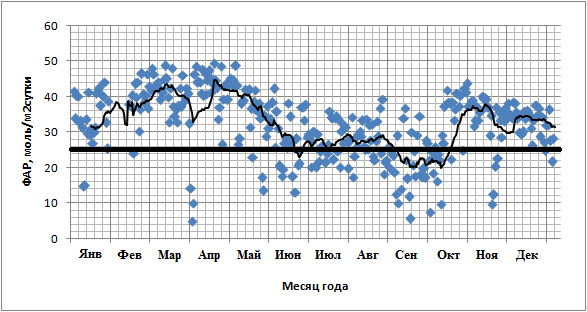
Рис. 10. Изменение суточной суммы ФАР (моль/м2сутки) в течение 2012 года. Жирной линией отмечен уровень насыщения ФАР (25 моль/м2сутки).
Глава 4. Обсуждение результатов
4.1. Таксационные описания
Проведённые таксационные описания выявили различия в биомассе между разными типами лесных сообществ. Полученные данные по биомассе отдельных типов лесных сообществ согласуются с данными литературы по аналогичным сообществам в других экваториальных странах (FAO Forestry paper-134, 1997).
Анализ картографического материала показал, что на территории, где расположен измерительный комплекс для пульсационных исследований, преобладает естественно восстанавливающийся лес, но значительную площадь (10-20%) занимают и два других изученных сообщества (лагерстремовый лес и редколесье). Данное распределение можно считать типичным только для северо-восточной равнинной части парка, но не для всего Кат Тьена в целом (где в значительной мере присутствуют территории, покрытые бамбуком, и другие типы леса).
На основании таксационных описаний можно сделать вывод о том, что в значительной мере леса парка обладают неполным потенциалом биомассы. При этом наибольший вклад в депонирование углерода можно ожидать в результате развития естественно восстанавливающихся древостоев.
Возможно, значительные части «редколесья» образовались в результате мощной деградации в прошлом, однако одной из важных причин являются и природные условия соответствующих территорий (приуроченность к более сырым, заболоченны участкам). Из-за этого потенциал развития у редколесья не должен быть большим, и значительный вклад в будущее депонирование углерода в лесных экосистемах парка этот тип леса оказать не сможет.
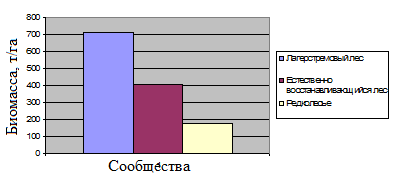
Рис. 11. Общее распределение биомассы в изученных сообществах (т/га).
4.2. Суточная динамика GPP
Максимальные средние суточные значения GPP в течение всех месяцев были сравнимы (~20-35 мкмоль/м2с по методу разделения потоков Lloyd and Taylor, ~15-35 мкмоль/м2с по методу разделения потоков Lasslop. Достижение точки максимума и диапазон области оптимума значений GPP включает время от 9 до 14 ч (по методу Lloyd-Taylor) и от 8 до 13 ч (по методу Lasslop). Сравнение двух методов показало, что при использования метода разделения по Lasslop оптимум сдвигается на более раннее время суток. Следует отметить, что диапазон оптимума по времени сходен и отмечается в обоих вариантах обработки полученных результатов. Коч (Koch et al., 1994), работавший в Панаме и изучавший суточную динамику фотосинтеза деревьев полога, обнаружил, что максимум фотосинтеза наблюдался в 11 часов местного времени, что также смещено относительно астрономического полудня. Сходные с нашими данные по суточной динамике GPP были получены и для других тропических лесов (Grace, 1995; Bonal et al., 2008; Desai et al., 2008; Goulden et al., 2004).
В марте кривая фотосинтеза выходит на уровень насыщения при значениях ФАР 900-1000 мкмоль/м2с, что составляет около 50% от возможных максимальных значений. Диапазон варьирования значений GPP при этом значении ФАР составляет от 12 до 35 мкмоль/м2с (в среднем около 20 мкмоль/м2с). При данном уровне ФАР присутствуют единичные максимальные значения (30-35 мкмоль/м2с). В июле насыщение начинается несколько позже, оно хорошо выражено в диапазоне ФАР 1000-1200 мкмоль/м2с при средних значениях GPP 25-30 мкмоль/м2с (варьирование от 20 до 45 мкмоль/м2с). Очень высокие значения GPP (более 40 мкмоль/м2с) также, как и в марте, являются редкими и заметно выделяются из основного массива данных. Наши данные согласуются с литературными. Например, (Goulden еt al., 2004), отметил начало насыщения при ФАР 500-1000 мкмоль/м2с для зрелых тропических лесов Амазонии. Сходные результаты получены и другими авторами (Malhi et al. 1998, Fan et al. 1990).
Наличие полуденной депрессии фотосинтеза показано для множества экосистем (Larcher, 1995), включая и тропические леса (Koch et al. 1994, Malhi et al. 1998, Zotz and Winter 1996). Исследователи объясняют это явление множеством факторов, включающих устьичный ответ на уменьшение давления водяных паров в воздухе, низкий водный потенциал листа, влияние на фотосинтез повышенной температуры воздуха, накоплением метаболитов и крахмала, внутренние циркадные ритмы и др. Влияние этих факторов является комплексным. Несмотря на это, наши данные позволяют рассмотреть подробнее влияние некоторых факторов на особенности суточного хода GPP в муссонном тропическом лесу южного Вьетнама.
Вероятно, одним из определяющих факторов в формировании полуденной депрессии фотосинтеза является рост дефицита водяных паров в воздухе (VPD). В марте максимальные значения GPP отмечаются при VPD на уровне 12 гПа, что соответствует оптимальным средним значениям GPP – около 17 мкмоль/м2с, после чего отмечается ингибирование фотосинтетической активности. В июле в широком диапазоне водного дефицита (от 1 до 12 гПа) изменения в величине GPP небольшие, но максимальные значения продукции отмечаются уже в области 7-8 гПа, которым соответствует средний оптимальный уровень GPP 23 мкмоль/м2с. Более низкий водный дефицит во влажный сезон (июль) способствует и более поддержанию значений GPP на более высоком уровне. Таким образом, во влажный сезон наблюдается меньшая зависимость GPP от этого фактора. К такому же выводу пришли и Koch с соавторами (1994), которые изучали фотосинтез и проводимость листа деревьев полога в лесах Панамы. Они обнаружили, что фотосинтез главным образом определяется проводимостью листа, которая в первую очередь зависит от VPD (Koch et al., 1994).
Высокая температура воздуха также может являться фактором, ограничивающим интенсивность фотосинтеза. При температурах воздуха выше 30 °С в течение всего года GPP начинает падать. Эта закономерность в большей степени выражена в июле, когда падение после насыщения более резкое. Вероятно, падение интенсивности фотосинтеза в данном случае связано не с повышением температуры воздуха, а с увеличением VPD. Влияние температуры на скорость фотосинтеза многократно описывалась различными исследователями, которые пришли к выводам, что температура до 30-35 °С ещё не ограничивает интенсивность фотосинтеза. Например, Lloyd and Farquhar (2008) показали, что температура 30 °С сама по себе не ограничивает скорость фотосинтеза тропических деревьев Амазонского бассейна.
4.3. Сезонная динамика GPP. В сезонном аспекте GPP варьирует в пределах от 220 гС/м2 в месяц в марте до 300-375 гС/м2 в месяц в июле-августе. При этом месячные суммы продукции в сухие месяцы несколько меньше, чем суммы, полученные во влажные месяцы (Рис. 8).
Стоит отметить, что максимальные месячные суммы GPP леса национального парка Кат Тьен в течение влажного сезона сравнимы с месячными суммами GPP широколиственных лесов в период пика вегетации. Например, в лиственничном лесу Томакомай (Tomakomai, Япония) самым продуктивным месяцем является июнь с GPP ~420 гС/м2 месяц (Saigusa et al., 2008), что выше, чем максимальная месячная продуктивность муссонного тропического леса национального парка Кат Тьен. Таким образом, общие высокие годовые суммы GPP в тропиках достигаются преимущественно не за счёт собственной высокой продуктивности, а за счёт того, что период вегетации длится в течение всего года, без периодов покоя. Это имеет место в том числе в сезонно-влажных тропиках, где наблюдается лишь относительно небольшое снижение GPP в течение засушливого периода (Bonal et al., 2008).
Характер сезонной динамики GPP в муссонном тропическом лесу национального парка Кат Тьен в целом типичен для региона. Так, согласно данным обзорной работы (Saigusa et al., 2008) GPP имеет тенденцию к уменьшению с декабря по март в муссонных диптерокарповых лесах Таиланда, где максимальные значения GPP в июле-сентябре составляют больше 350 гC/м2 в месяц, а наименьшие значения в январе-марте 200 гC/м2 в месяц или менее.
Снижение GPP в сухой сезон можно объяснить несколькими факторами. В первую очередь это может быть связано с тем, что многие деревья полога полностью сбрасывают листья. Пик листопада приходится на декабрь-январь (Кузнецов, 2012). Деревья верхнего яруса не имеют листвы в январе-марте, и именно эти месяцы характеризуются наименьшими значениями GPP. Это предположение может быть подтверждено тем, что в вечнозелёных влажных тропических лесах Амазонии, в отличие от полулистопадных лесов национального парка Кат Тьен, не было найдено разницы в продуктивности в сухой и во влажный сезоны (Grace et all., 1995). Это подтверждается и исследователями газообмена тропических деревьев на уровне листа, не показавших значимых сезонных отличий (McWilliam et al., 1996; Roberts et al., 1996).
Определённое влияние на характер сезонной динамики GPP оказало изменение уровня ФАР, описанное для множества других тропических экосистем (Malhi et al., 1998; Malhi & Grace, 2000; Loescher et al., 2003; Goulden et al., 2004, Huete et al., 2006; Hutyra et al., 2007). Об этом говорит слабая достоверная корреляция между суточными суммами ФАР и суточными суммами GPP (рис. 9, R2=0.18). Сезонные колебания ФАР действительно значительны и при этом преимущественно вызваны не собственным изменением уровня солнечной радиации, а значительно большей облачностью во влажный период. Но насыщение фотосинтеза происходит при ФАР уже 1100 мкмолль/м2с, что составляет лишь 55% от максимальной ФАР, достигаемой в полдень в течение сухого сезона. Таким образом, суточные суммы GPP, близкие к максимальным, достигаются уже при суточной сумме ФАР в 25 моль/м2 в сутки. Дней с такой суммой ФАР практически нет в течение сухого сезона, и даже в течение самых влажных месяцев их менее половины.
Мы предполагали, что влияние на снижение продуктивности в сухой период может также оказывать снижение влажности в верхних слоях почвы. В тропических сообществах Южной Америки описаны эффекты снижения роста деревьев и устьичной транспирации (например, Bonal et al., 2000; Bonal & Guehl, 2001), а также ксилемной эмболии (Tyree et al., 1998) при снижении содержания воды в верхних слоях почвы. Однако, влияние влажности почвы на отмеченное в нашей работе снижение GPP в сухой сезон в лесах национального парка Кат Тьен должно быть незначительным, т.к. нами не было зафиксировано достоверной зависимости между усреднённой суточной влажностью почвы на глубине 50 см и суточными суммами GPP. Возможно, это связано с тем, что глубина залегания корней позволяет растениям избегать засухи в сухой сезон.
Увеличение GPP во влажный сезон может быть также связано с увеличением фотосинтеза, связанного с локальным повышением концентрации CO2 из-за более интенсивного почвенного дыхания (Структура и функции…, 2011).
На месячные суммы GPP может оказывать влияние и температура воздуха, что показано при сравнительном изучении продуктивности горных, предгорных и равнинных тропических лесов (Malhi, 2012). В горных тропических лесах средние температуры воздуха несколько ниже, из-за чего месячные суммы также ниже относительно равнинных лесов, произрастающих на той же самой широте. Однако из-за небольших различий (в пределах 5 градусов) среднесуточных температур воздуха в течение года это влияние также не может быть большим.
Годовая сумма GPP в муссонном тропическом лесу Вьетнама составила 29,30 МгС/га в год (2,93 кгС/м3, при использовании разделения потоков по Lasslop et al., 2010), 35,20 МгС/га в год (3,52 кгС/м3, при использовании разделения потоков по Lloyd & Taylor, 1994). Данные годовые суммы GPP являются типичными для региона. Так, годовые суммы GPP в Mae Klong и Sakaerat (Таиланд) составляют 32.3 (Hirata et al., 2008) и 38.1 МгС/га (Gamo et al., 2005) соответственно. В малайском заповеднике Pasoh годовая сумма GPP равна 31.2 МгС/га (Kosugi et al., 2008). Похожие годовые суммы характерны и для влажных тропических лесов Южной Америки. Например, на биологической станции Ла Сельва (La Selva Biological Station, Коста-Рика) зарегистрирована годовая сумма GPP 29.5 МгС/га (Loescher et al., 2003). В тропическом лесу Paracou (Французская Гвиана) зарегистрирована годовая сумма GPP 37.3 МгС/га (Bonal et al., 2008).
Выводы
- Леса южной части национального парка Кат Тьен очень гетерогенны, демонстрируют значительно различающийся уровень общей биомассы (от 200 до 700 т/га) на участках с различными уровнями антропогенной нарушенности.
- Характер суточного хода валовой первичной продукции в муссонном тропическом лесу схож в течение всего года, как в месяцы сухого сезона, так и в месяцы влажного сезона. Суточная динамика GPP определяется суточной динамикой ФАР.
- Дефицит водяных паров в воздухе, вероятно, основной фактор, приводящий к появлению полуденной депрессии фотосинтеза (после 13 ч), которая была хорошо выражена как в сухой, так и во влажный сезоны.
- Месячные суммы GPP изменяются от 200 гС/м2 в месяц (в марте) до 300-350 гС/м2 в месяц (в июле).
- Основным фактором, определяющим сезонную динамику GPP в муссонном тропическом лесу национального парка Кат Тьен, является изменение индекса листовой поверхности в течение года. Наименьшие значения продукции совпадают с сухим сезоном.
- Суммарная годовая валовая продукция составила 29.3-35.2 МгС/га, что сопоставимо с годовыми суммами продукции в других тропических лесов (в т.ч. сезонно-влажных).
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ:
Статьи в изданиях, рекомендованных ВАК РФ
- Евдокимова Е.В. Особенности водного режима некоторых видов растений в тропическом лесу на юге Вьетнама во влажный сезон / Е.В. Евдокимова, А.О. Новичонок, Е.Ф. Марковская, Ю.А. Курбатова // Ученые записки ПетрГУ. – №4 (125). – 2012. – с. 19-24.
- Novichonok A. Preliminary forest plant biomass inventory in moonson tropical forest in Cat Tien national park (South Vietnam) / A. Novichonok, E. Evdokimova, E. Markovskaya, J. Kurbatova // Proceedings of Petrozavodsk State University. – Vol. 8 (129), part 2. – 2012. – Pp. 13-16.
Материалы научных конференций
- Новичонок А.О. Леса южной части национального парка Кат Тьен (южный Вьетнам) / А.О. Новичонок // Теоретические и практические вопросы развития научной мысли в современном мире: сборник статей II Международной научно-практической конференции. 29-30 апреля 2013 г.: в 4 ч. Ч 4. – Уфа: РИЦ БашГУ, 2013. – с. 21-24.
- Avilov V., Anichkin A., Descherevskaya O., Evdokimova E., Thinh Nguyen Van, Novichonok A., Luu Do Phong, Kurbatova J., and Lopes de Gerenyu V. Soil respiration in tropical seasonal forest of Southern Vietnam // Geophysical Research Abstracts, EGU General Assembly 2013, Vienna, Austria, Vol. 15, EGU2013-9463-8, 2013.
- Deshcherevskaya O., Avilov V., Dinh Ba Duy, Novichonok A., Tran Cong Huan and Kurbatova J. Heat, water and carbon dioxide fluxes at the first Vietnamese eddy covariance site in tropical seasonal forest // Geophysical Research Abstracts, EGU General Assembly 2013, Vienna, Austria, Vol. 15, EGU2013-10397-6, 2013.
- Kurbatova J., Ba Duy Dinh, Avilov V., Deshcherevskaya O., Novichonok A., Phong Luu Do, Cong Huan Tran, Kuznetsov A. Energy and CO2 fluxes at the first vietnamese eddy covariance site in tropical seasonal forest // Proceedings of International Joint Conference of 11th AsiaFlux International Workshop, 3rd HESS, and 14th Annual Meeting of KSAFM. 21-24 August 2013. Seoul, Korea. P. 85.
[1] http://www.bgc-jena.mpg.de/~MDIwork/eddyproc/
[2] По всей видимости, данные по биомассе Lagerstroemia calyculata, приведённые здесь, завышены, т.к. для её вычисления использовалось общее аллометрическое уравнение для влажных тропиков, которое не учитывает особенности данного конкретного вида – формирование у возрастных деревьев значительных пустот внутри стволов.
[3] Условное название; под ним понимается сообщество с малым количеством крупных деревьев, разреженным пологом и относительно густым подростом.