

Исследование поведения настоящих тюленей в условиях неволи
На правах рукописи
Березина Ирина Александровна
ИССЛЕДОВАНИЕ ПОВЕДЕНИЯ НАСТОЯЩИХ ТЮЛЕНЕЙ В УСЛОВИЯХ НЕВОЛИ
03.02.04 – зоология
Автореферат диссертации на соискание ученой степени кандидата
биологических наук
Петрозаводск
2013
Работа выполнена в Мурманском морском биологическом институте Кольского научного центра Российской академии наук и Мурманском государственном техническом университете
Научный руководитель доктор биологических наук,
заведующий лабораторией морских
млекопитающих Мурманского морского
биологического института КНЦ РАН
Кавцевич Николай Николаевич
Официальные оппоненты: Илюха Виктор Александрович,
доктор биологических наук, доцент,
Федеральное государственное бюджетное
учреждение науки Институт биологии
Карельского научного центра РАН,
заведующий лабораторией экологической
физиологии животных
Медведев Николай Владимирович,
доктор биологических наук,
Федеральное государственное бюджетное образовательное учреждение высшего
профессионального образования
Петрозаводский государственный
университет, профессор кафедры зоологии
и экологии
Ведущая организация Федеральное государственное бюджетное образовательное учреждение высшего профессионального образования «Мурманский государственный гуманитарный университет»
Защита состоится 15 мая 2013 г. в 14.00 в ауд. 117 на заседании диссертационного совета Д 212.190.01 при Петрозаводском государственном университете по адресу: 185910, Республика Карелия, г. Петрозаводск, ул.Красноармейская, 31.
С диссертацией можно ознакомиться в библиотеке Петрозаводского государственного университета, с авторефератом – на сайтах http://vak.ed.gov.ru и www.petrsu.ru.
Автореферат разослан " " апреля 2013 г.

Ученый секретарь диссертационного совета
кандидат биологических наук Дзюбук И.М.
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы. Хозяйственная деятельность человека в Мировом океане вынуждает морских млекопитающих менять свой образ жизни и поведение, приспосабливаясь к условиям внешней среды. Возрастающая частота контактов морских млекопитающих с человеком вызывают у них изменения поведения, особенно ярко проявляющиеся в условиях неволи и не всегда экологически адекватные.
Зрительный и слуховой аппарат тюленей хорошо приспособлены к функционированию как под водой, так и на суше (Андреев, 1978; Морозов, 1983; Ренуф, 1989; Fobes, Smock, 1981). Однако на больших глубинах, куда не проникает солнечный свет, и в мутных водах зрение не самый надежный анализатор, поэтому значительную часть информации морские животные получают через слух, тем более что в воде, по сравнению с воздухом, звуковые колебания поглощаются в 700 раз меньше (Уолкер, 1969). Поэтому исследование возможностей слуховой системы исключительно важно для охраны и рационального использования морских млекопитающих (Беликов, Белькович, 2002; Супин, 2008).
Экспериментальные исследования поведения дельфинов и ушастых тюленей проводятся достаточно широко. (Крушинский и др., 1972; Крушинский, 1977; Крушинский и др., 1982, 1987; Стародубцев, 1982, 1992, 2000-2008; Надолишняя, Стародубцев, 2000, 2002; Стародубцев, Надолишняя, 2010 Надолишняя и др., 2004, Флёсс и др., 1987, 1988, 1990; Чечина, 2004, 2006, 2010; Лямин, 2012; Якимов, Лебедева, 2012 Roitblat et al., 1990; Pack, Herman, 1995; Harley et al., 1996; Mercado et al., 2000; Tschudin et al., 2001; Jaakkola et al., 2003; Cardwell, Harley, 2005; Pack, Herman, 2007).
Данные об участии зрительной и слуховой систем в организации поведения ластоногих немногочисленны и они важны при решении вопросов охраны, для рационального ведения промысла, при разработке систем управления их поведением. Исследования в этой области особенно актуальны для российского Севера, так как антропогенная нагрузка на арктические моря год от года возрастает. Систематические исследования роли процессов переработки зрительной информации в построении поведенческих программ проводились лишь на ушастых тюленях, северных морских котиках (Близнюк, Дашевский, 1989; Дашевский, Близнюк, 1989).
Однако пока очень мало экспериментов по изучению поведения, анализаторов и познавательных способностей настоящих тюленей (Levenson, Schusterman, 1999; Schusterman, 2000; Bodson et al. 2005, Hanke et al. 2005; Mauck, Dehnhardt, 2005). В то же время, исследования поведенческих реакций на различные зрительные раздражители позволили оценить важнейшие параметры высшей нервной деятельности серого тюленя (Надолишняя и др., 2006) и кольчатой нерпы (Стародубцев и др., 2012). Исследования поведения этих животных в неволе способствуют обогащению знаний этологии и теории адаптации.
Изучение поведенческих адаптаций морских млекопитающих также весьма актуально в связи с использованием этих животных как помощников человека при освоении Мирового океана, участников демонстрационных программ и как возможных объектов доместикации (Близнюк, 1984, Мишин, 1991). С этой точки зрения исследование поведения морских млекопитающих позволит разработать рекомендации по обеспечению необходимых условий, обеспечивающих максимально возможную продолжительность их жизни в неволе, снизить действие стрессовых ситуаций на морских млекопитающих, оптимизировать процессы их приручения и обучения.
Цель исследования. Выявить закономерности формирования двигательных программ при воздействии зрительных и слуховых раздражителей у представителей семейства настоящие тюлени различного возраста и времени пребывания в неволе.
Для достижения цели были поставлены следующие задачи:
1. Изучить поведение представителей трех видов семейства настоящие тюлени при адаптации к условиям неволи.
2. Исследовать динамику выработки двигательных условных рефлексов на двумерные и трехмерные зрительные раздражители. Определить степень различий формы плоских геометрических фигур, распознаваемых тюленями.
3. Оценить способности тюленей к обобщению методом "выбор по образцу".
4. Исследовать динамику выработки двигательных условных рефлексов на звуки чистых тонов. Определить минимальный частотный интервал между двумя разными звуковыми сигналами, распознаваемыми тюленями.
5. Определить характеристики сложных звуковых сигналов, существенные для их распознавания тюленями.
Научная новизна результатов работы. Впервые получены количественные экспериментальные данные о динамике формирования условно-рефлекторных реакций на зрительные и слуховые стимулы у представителей 3-х видов семейства настоящие тюлени. Получены сравнительные данные об особенностях поведения и обучения в ходе приспособления к условиям неволи при содержании ластоногих в океанариуме. Впервые методом двигательных условных рефлексов проведено определение частотно-временных характеристик слухового анализатора у морского зайца, серого и гренландского тюленей.
Практическая и теоретическая значимость работы. Результаты настоящей работы могут быть применены в сравнительных физиологических и зоопсихологических исследованиях морских млекопитающих, а также могут служить основой для оценки способности настоящих тюленей к обобщению и при изучении элементарной рассудочной деятельности этих животных. Полученные данные можно использовать при подготовке ластоногих к демонстрационным выступлениям в океанариуме и для выполнения служебных работ в составе морских биотехнических систем. Сведения об особенностях реагирования тюленей на различные зрительные и слуховые раздражители применимы при выборе и обосновании мер по защите морских млекопитающих в условиях возрастающего антропогенного воздействия.
Полученные материалы используются при чтении лекционных курсов и проведении практических занятий для студентов биологического направления факультета пищевых технологий и биологии Мурманского государственного технического университета.
Основные положения, выносимые на защиту.
1. Содержащиеся в океанариуме тюлени различаются по скорости и прочности выработки условных рефлексов. С возрастом способность к обучению у тюленей не снижается. У них формируется "установка на обучение" с использованием зрительных и слуховых раздражителей, различающихся по определенным признакам. Условные рефлексы второго рада, инструментальные, закрепляются легче и прочнее чем рефлексы первого рода, классические;
2. Представители исследованных видов семейства настоящие тюлени способны к дифференцировке двух- и трехмерных предметов различной формы и размеров;
3. Серые тюлени проявляют высокий уровень способности к обобщению при "выборе по образцу" трехмерных предметов
4. Морской заяц, гренландский и серый тюлени способны различать звуки в диапазоне 200-700 Гц по относительному признаку "низкий-высокий".
5. Ведущую роль при распознавании тюленями сложных звуковых сигналов играет периодичность следования составляющих их простых звуков чистых тонов.
Апробация материалов диссертации. Результаты работы докладывались и обсуждались на региональной научной студенческой конференции "Естественнонаучные проблемы арктического региона" (Мурманск, 2001); на конференции "Морские млекопитающие Голарктики" (Иркутск, 2002); конференциях молодых ученых ММБИ (Мурманск, 2002 и 2011); на научно-практическом семинаре "Морские животные на службе человеку" (Североморск, 2010); школе молодых ученых МГТУ по морской биологии (Мурманск, 2012); международных научно-технических конференциях МГТУ "Наука и образование" (Мурманск, 2007, 2011 и 2012).
Личный вклад автора. Материалы, использованные в диссертации, получены в результате проведения научно-исследовательских работ: № 01.2.00110682 "Экспериментальное изучение поведения настоящих тюленей", № 0120.0509313 "Биологические и научно-технические основы использования морских млекопитающих в системах охраны стратегических объектов", № 01 2009 55546 "Исследование поведения и сенсорных систем ластоногих на суше и под водой", № 01 2011 65652 "Разработка научных основ и методов использования арктических тюленей в биотехнических системах хозяйственного и оборонного значения".
Автор принимал непосредственное личное участие в проведении анализа научной литературы, подборе и обосновании методов исследования, в сборе, обработке первичного материала и формировании исходных баз данных, статистической обработке, анализе и обобщении результатов исследований. Все работы по накоплению, статистической обработке, анализу и обобщению материалов диссертации выполнены автором лично.
Публикации. По теме диссертации опубликовано 18 работ, в том числе 2 статьи в журналах, рекомендованных ВАК РФ.
Структура и объем работы. Диссертация изложена на страницах, состоит из введения, обзора литературы, главы "Материалы и методы исследования", 3-х глав с результатами исследований и их обсуждением, заключения, выводов, списка литературы, включающего источников, из которых на русском языке, содержит рисунок и таблицы.
СОДЕРЖАНИЕ РАБОТЫ
ГЛАВА 2 МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Объекты исследования: серый (Halichoerus grypus) и гренландский (Pagophilus groenlandicus) тюлени, морской заяц (Erignathus barbatus). Работу проводили в океанариуме г. Мурманска (ООО "Арктиксервис – Мурманский океанариум") и в аквариальном комплексе ММБИ в Кольском заливе. Общее число исследованных животных составило: серый тюлень – 6 гренландский тюлень – 1; морской заяц – 1.
Методики исследования. Все эксперименты проводили по стандартной двигательно-пищевой методике. Пищевым подкреплением служила размороженная рыба (целая или резаная на куски в зависимости от ее размера): сельдь, мойва, путассу, ставрида, сайка. Критерием обучения было достижение 95%-го уровня правильных ответов в двух опытах подряд. По методике одновременной дифференцировки стимулы предъявляли одновременно. Задача животного – выбрать "правильный" стимул и ответить на него. При последовательной дифференцировке оба стимула предъявляли по одному. Животное должно было научиться отвечать на "правильный" (подкрепляемый) стимул и тормозить реакцию в присутствии "неправильного" (Дьюсбери, 1981). При проведении экспериментов использовалась либо методика одновременного (распознавание трехмерных раздражителей и выбор предметов по образцу) либо последовательного предъявления стимулов (распознавание двумерных зрительных раздражителей и эксперименты со звуками чистых тонов и голосовыми сигналами).
Занятия с животными проводились 1-3 раза в день, 1-5 дней в неделю, в зависимости от занятости животных в демонстрационных программах океанариума. Только тюлень №3 был свободен от участия в шоу-программах, и с ним занятия проходили 3 раза в день 5 дней в неделю.
Методика обучения распознаванию двумерных и трехмерных зрительных раздражителей. В качестве зрительных стимулов использованы изображения простых геометрических фигур (белые кольцо и треугольник на черных квадратах 2525см). Табличку-символ помещали на демонстрационную доску (белый щит 11.5 м), расположенную на помосте. В экспериментах по выработке зрительной дифференцировки трехмерных зрительных раздражителей использовались тор (внешний диаметр 15 см, внутренний – 10 см), шар диаметром 9.5 см и цилиндр (высота 20 см, диаметр 4.5 см) из белого пластика, которые располагались на стенке бассейна на расстоянии 1-1.5 м от животного, находящегося на стартовой позиции напротив тренера.
При обучении распознаванию зрительных образов первоначально животных приучали в ответ на сигнал-жест экспериментатора выполнять адекватное локомоторное движение, выполнение которого подкреплялось пищей. Затем приступали к выработке реакции на зрительный образ. При этом животному предъявлялся зрительный образ, после чего следовал сигнал-жест экспериментатора, и после выполнения адекватного локомоторного навыка животное получало подкрепление. После упрочения поведенческих реакций у тюленей тестировали реакцию на предъявляемые зрительные раздражители. Правильный выбор и в этом случае подкреплялся пищей.
Методика постоянного выбора тюленем одной из геометрических фигур.
Животных приучали после сигнала-жеста экспериментатора выполнять адекватный локомоторный навык (касание креста, независимо от того, с какой стороны относительно животного он находится). Правильный выбор подкреплялся пищей.
Сторона предъявления фигур менялась псевдослучайно: не более тех раз подряд с одной стороны и так, чтобы каждая фигура занимала положение право-лево относительно животного не более 15 раз на опыт, таким образом, животному фигуры предъявлялись 30 раз за опыт.
Всего в работе было использовано 5 вариантов креста 2525 см с шириной лопастей 5, 9, 13, 17 и 21 см (36.0, 59.0, 77.0, 89.8 и 97.4% от площади квадрата, соответственно), которые предъявляли одновременно с квадратом 2525 см.
Методика выбора трехмерных геометрических фигур в соответствии с образцом. Метод "выбор по образцу" предложен для исследования обезьян Н.Н. Ладыгиной-Котс (1923). Он состоит в том, что животному демонстрируют стимул-образец и два или несколько стимулов для сопоставления с ним, подкрепляя выбор того, который соответствует образцу (Зорина, Полетаева, 2001).
На стенке вольера закрепляли сначала один предмет, впоследствии – два и три предмета. На подготовительном занятии животное лишь касалось носом висящего предмета по жестовой команде тренера. Во время следующего опыта экспериментатор предъявлял тюленю предмет, который требовалось обозначить на стенке вольера (прикоснуться к нему носом) после жестовой команды.
Предъявление предметов в экспериментах осуществлялось с использованием таблицы случайной комбинации двух или трех чисел, каждое из которых соответствовало одному из предъявляемых предметов.
Распознавание простых и сложных звуковых стимулов. В качестве раздражителей при выработке у тюленей новых условных рефлексов использованы звуки чистых тонов. Они получены при помощи цифрового генератора программы "WaveLab 3.0". Для воспроизведения звуковых раздражителей применяли усилитель и акустическую систему для музыкального сопровождения представлений в океанариуме. Эксперимент по определение минимального частотного интервала между двумя разными звуковыми сигналами, распознаваемыми тюленями, проводился на 3-х тюленях: самце морского зайца (тюлень №7), самке гренландского тюленя (тюлень №8), самке серого тюленя (тюлень №5). При обучении распознаванию звуковых раздражителей первоначально животных обучали в ответ на сигнал-жест экспериментатора выполнять адекватное локомоторное движение, выполнение которого подкреплялось пищей. Затем приступали к выработке реакции на звук 200 Гц - "стыдно" (тюлень прикрывает морду ластом), на звук 700 Гц - "ладушки" (тюлень похлопывает себя ластом по боку) при этом животному предъявлялся звук, после чего следовал сигнал-жест экспериментатора и после выполнения адекватного локомоторного навыка животное получало подкрепление. После упрочения поведенческих реакций у тюленей тестировали реакцию на предъявляемые звуковые раздражители. Правильный выбор и в этом случае подкреплялся пищей. В следующих сериях экспериментов частоту первого звукового стимула увеличивали, а второго уменьшали.
Звуки предъявляли попарно в псевдослучайном порядке. Каждый звук в опыте предъявлялся 15 раз, но не более трех раз подряд. Всего за каждый опыт тюлень получает 30 предъявлений. Первоначально была выбрана пара звуков 200 Гц и 700 Гц. Затем, когда в ходе экспериментов достигался 95%-ный уровень правильных ответов в двух опытах подряд, переходили к следующей паре звуков. Шаг с которым менялась частота предъявляемых звуков первоначально составил 20 Гц, после достижения частоты 420 Гц и 480 Гц этот шаг составил 5 Гц и конце эксперимента шаг составил 1Гц.
Для анализа сложных звуковых сигналов использовали программу "WaveLab 3.0".
Методы анализа результатов. Обработку полученных результатов проводили при помощи программ Exel, Statsoft Statistica V6.0, Systat 5W. При статистических оценках и сравнениях использовали стандартные методы (Урбах, 1963). При определении доверительных интервалов для доли случайного выбора стимула из двух или трех, предъявляемых тюленю, использовали статистические параметры альтернативного распределения при Р=0.95. Среднеквадратичное отклонение вычисляли по формуле:
![]()
![]() ,
,
где S – среднеквадратичное отклонение; р – доля выбора в (50% при предъявлении 2-х стимулов, и 33,3% для трех стимулов); N – количество предъявлений за одно занятие. Доверительный интервал (для Р=0.95) составляет 1.96S (Урбах, 1963). Верхняя граница случайного выбора = доверительный интервал + p.
Глава 3 ОБЩАЯ ХАРАКТЕРИСТИКА ПОВЕДЕНИЯ ЭКСПЕРИМЕНТАЛЬНЫХ ЖИВОТНЫХ
3.1 Особенности поведения тюленей в неволе
Детеныши тюленей начинают питаться морскими организмами после ювенильной линьки. Период так называемого "раскорма" в океанариуме совпал со временем приручения животных и выработки у них базовых условных рефлексов, необходимых для последующего обучения. Это, в первую очередь, "таргет" (от англ. target - мишень), когда тюлень по команде касается носом руки экспериментатора или предмета, который он предлагает. Был также выработан условный рефлекс на свисток, "брижд-сигнал" (от англ. bridge - мост), т.е. сигнал, предшествующий пищевому подкреплению после правильной реакции на команду.
Новые движения у тюленей, как выработанные по схеме классического условного рефлекса, так и выработанные по схеме инструментального условного рефлекса, закрепляются очень хорошо и прочно. Всего каждый серый тюлень и морской заяц различает около 30 команд. Гренландский тюлень – около 20. Причиной этого является короткое, по сравнению с серыми тюленями и морским зайцем, время содержания в условиях неволи. В качестве команд-сигналов могут выступать не только жест или голос, но и различные предметы – "таргет" (шарик, закрепленный на короткой палочке), мяч, резиновая груша, обруч, ведро, лодка и др.
В присутствии тренера все тюлени позволяют к себе прикасаться, постоянно наблюдая в то же время за новым для себя объектом. Гораздо лучше, чем у серых тюленей, отработан тактильный контакт у зайца: он позволяет прикасаться к любой части тела, не проявляя при этом какой-либо агрессивной или оборонительной реакции, выполняет все команды которым обучен. Как и другие животные, заяц при резких движениях незнакомых людей настораживается, уходит в воду, если лежал на помосте, или в дальнюю от человека часть бассейна. Тюлени способны перенимать новые движения и друг у друга, и у других тюленей. Животные различают работающих с ними людей не только по внешнему виду, но, вероятно, и по индивидуальным особенностям проведения ими занятий, динамическому стереотипу, склонностям и предпочтениям, которые обусловлены как профессиональным уровнем, так и психологическими установками тренера.
У всех тюленей новые двигательные рефлексы вырабатываются легко и закрепляются прочно, но скорость выработки реакций на различные сигналы зависит от их сложности. После длительного перерыва в занятиях угасшие рефлексы восстанавливаются достаточно быстро – необходимо 2-3 предъявления, и команда снова выполняется правильно. Тюлени предлагают большое количество различных движений, которые впоследствии можно закрепить, используя определенную команду.
В целом, сравнивая поведение морского зайца и тюленей других видов (серого, гренландского) в океанариуме, следует отметить флегматичность, медлительность при выполнении команд. Эти особенности, вероятно, связаны с образом жизни лахтака, кормящегося в природных условиях главным образом малоподвижными бентосными организмами и не являющегося, следовательно, таким активным хищником, как большинство настоящих тюленей.
Глава 4 РАСПОЗНАВАНИЕ ТЮЛЕНЯМИ ЗРИТЕЛЬНЫХ СТИМУЛОВ
4.1 Выработка дифференцировочных условных рефлексов на двумерные зрительные стимулы
На первом этапе работы тюленей обучали "новым", ранее не входившим в их репертуар на представлениях движениям, выполняемым по определенной жестовой команде. 1 – кольцо – самка серого тюленя №2 совершает полный круг вокруг экспериментатора, стоящего на помосте, самец серого тюленя №1 выполняет "колечко" (прогибается в спине, касаясь носом хвоста); 2 – треугольник – тюлень №2 выходит на левый край бассейна, тюлень №1 – "стыдно" (закрывает глаз передним ластом).
Обучение на 1 этапе заняло у животных разное количество занятий: 9 у тюленя №2 и 10 у тюленя №1. У тюленя №2 уровень правильных ответов (выполнение движений 1 и 2) составил 96.0±2.3%), у тюленя №1 – 90.3±2.8%, за последние 5 тренировок 96.3±0.9% тюленю №2 на первом этапе потребовалось 180 предъявлений, тюленю №1 – 250. Высокую скорость выработки новых условных рефлексов можно объяснить тем, что у животных сформирована установка на обучение по жестовым командам.
На 2 этапе проводилось установление и закрепление связи: зрительный стимул (кольцо или треугольник) - жест - движение животного. Данный этап занял у тюленя №1 – 16, у тюленя №2 – 20 занятий. Тюлень №2 получил 330 предъявлений (97.8±1.1% правильных ответов), тюлень №1 - 289 (99.6±0.4% правильных ответов).
В последующих опытах животным предъявляли только таблички-символы (кольцо и треугольник попеременно). В результате среднее число правильных зрительных дифференцировок (соответствие движения животного определенному изображению) после 22 опытов с самцом (тюлень №1) и 20 с самкой (тюлень №2) по 25 предъявлений в каждом незначительно превышало случайную величину 58.3±2.7% и 61.5±3.9% соответственно. При применении дополнительных методических приемов для переключения внимания животных с экспериментатора на табличку с изображением уровень правильных ответов в отдельных опытах повышался до 80-100%. При обучении по этой же стандартной методике морскому котику потребовалось 79 опытов с 51 предъявлением в каждом для достижения уровня дифференцировки 95%. В начале же обучения наблюдалось 35 – 40% межсигнальных (т.е. неправильных) реакций (Дашевский, Близнюк, 1989). Кроме того, в отличие от наших экспериментов, в опытах, которые проводили упомянутые авторы, использовались животные, не участвовавшие в представлениях.
По описанной выше схеме опыт был проведен с самцом серого тюленя №6 и самцом морского зайца (тюлень №7). У животных проводилась выработка условных рефлексов на зрительные раздражители, "крест" и "квадрат". Для установления связи "геометрическая фигура – сигнал-жест тренера – выполнение элемента – подкрепление" были проведены три занятия с комплексным раздражителем: изображение в сочетании с жестом и голосовой командой тренера. На первых 2-х занятиях с тюленем №7 и 3 занятиях с тюленем №6 уровень правильных ответов составил 100%. В последующих опытах, без сопровождения голосовой командой, было замечено, что животные сосредотачиваются на руках тренера, воспринимая любое движение за жест, а предъявляемая фигура остается без внимания. Этот эксперимент подтвердил ранее высказанное мнение (Дашевский, Близнюк, 1989), что при дифференцировке зрительных стимулов у ластоногих ведущую роль играет переключение внимания животного с экспериментатора на предъявляемый зрительный раздражитель.
4.2 Различение тюленями геометрических фигур разной степени подобия
В данной серии экспериментов методика работы по различению двумерных геометрических фигур серым тюленем и морским зайцем была изменена: с целью достижения переключения внимания животного на таблички-символы, их расположили на стенке бассейна, а не на помосте, и животные осуществляли выбор, подплыв к распознаваемым объектам.
С морским зайцем было проведено 17 занятий (рис. 1) с предъявлением ему 5 вариантов креста. В 4-х вариантах (опыты 1-13) морской заяц (тюлень №7) очень уверенно и быстро дифференцировал крест от квадрата. Однако в варианте 5 (ширина сторон креста 21 см) достоверная дифференцировка отсутствовала, из чего можно заключить, что такая фигура воспринимается животным как квадрат. При работе с вариантом 5 было отмечено, что животное использовало тактику выбора в большинстве случаев фигуры, расположенной слева, что позволяло получать подкрепление за случайно выбранный правильный ответ. В отличие от тюленя №7, тюлень №6 уверенно распознал и 5-й вариант креста, наиболее сходный с квадратом.
Вероятно, выявленные различия в способности распознавать геометрические фигуры обусловлено разным образом жизни, способом питания этих животных. Серые тюлени – активные хищники, а морские зайцы бентофаги и при поиске пищи для них большее значение имеет чувствительность вибрисс, чем зрение. Суммируя вышесказанное, можно сделать вывод, что настоящие тюлени хорошо дифференцируют плоские геометрические фигуры по форме, а не только объемные, как считалось ранее.
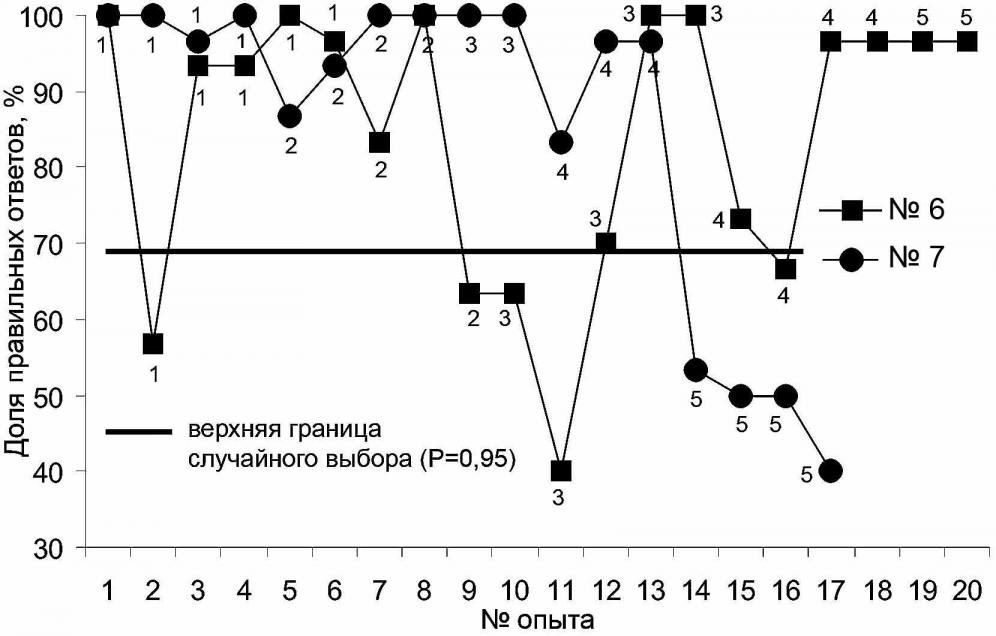
Рис. 1. Различение квадрата и креста самцом морского зайца (тюлень №7) и самцом серого тюленя (тюлень №6), 1-5 – варианты креста
4.3 Распознавание трехмерных зрительных раздражителей
Выработка дифференцировочных условных рефлексов на трехмерные зрительные раздражители
Работа проводилась на двух молодых серых тюленях самце и самке (тюлень №3 и тюлень №4). К началу эксперимента животные имели навыки выполнения различных движений по жестовым командам. Всего с тюленем №3 проведено 24 опыта с числом предъявлений 32 в каждом, с тюленем №4 – 24 (число предъявлений 40).
Эксперименты начали с установления и закрепления связи: предъявление предмета – сигнал-жест экспериментатора – выполнение адекватного движения – подкрепление. Соответствие стимулов и движений было следующим: 1 - тор (кольцо) – кружиться в воде вокруг своей оси; 2 – шар (мячик) – показать зубы; 3 – цилиндр (отрезок трубы) – протянуть из воды правый передний ласт.
Экспериментатор показывал животному предмет, делал определенный жест, после чего тюлень должен был выполнять соответствующее движение. Затем (при правильном ответе) следовало подкрепление. Занятия проводились с тюленем №3 в апреле, а с тюленем №4 в сентябре. Это явно сказалось на результатах опытов, поскольку она(самка серого тюленя) имела возможность наблюдать за занятиями с самцом. В ответ на предъявление кольца во время одной из тренировок при подготовке к выступлению она сразу, без обучения, выполнила движение, которое соответствует предъявлению кольца у самца. В данном случае ярко проявилась способность тюленя к обучению при помощи подражания.
Данный этап работы занял у тюленя №3 и у тюленя №4 по 6 занятий. Тюлень №4 получила 220 предъявлений (79.93±5.08% правильных ответов), тюлень №3 – 161 (71.68±7.32% правильных ответов). В последующих опытах по выработке дифференцировки попеременно предъявляли по одному из трех предметов (кольцо, мячик и трубу). Порядок предъявления стимулов был псевдослучайным (один стимул не более трех раз подряд). Таких занятий проведено по 18 с обоими тюленями. Среднее число правильных ответов у тюленя №3 составило 92.87±1.57%, (182 предъявления мяча, 188 – кольца, 224 – трубы), у тюленя №4 – 81.78±2,84% (181, 204, 383 предъявления, соответственно) (рис. 2).
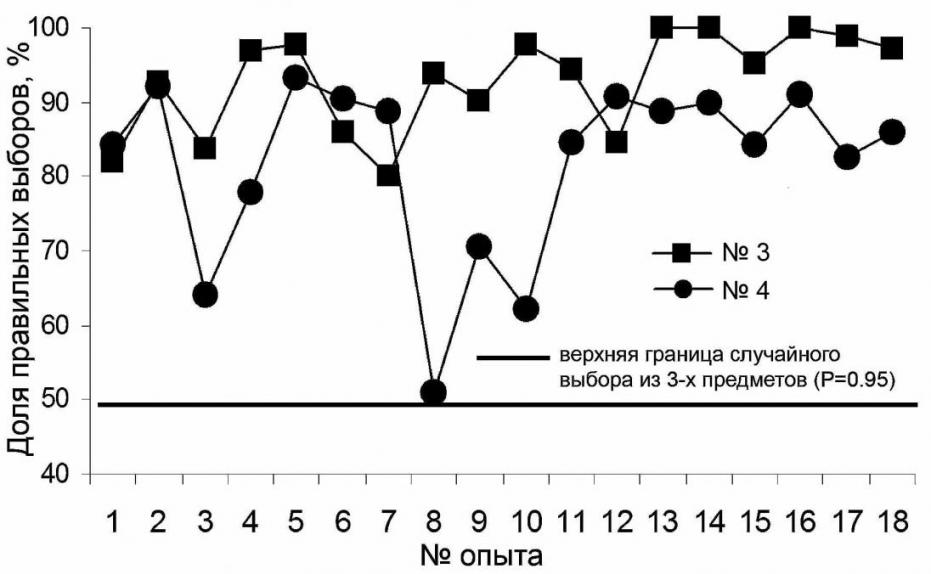
Рис. 2. Динамика выработки дифференцировки трехмерных зрительных раздражителей
Уровень правильных дифференцировок у тюленя №3 был выше, чем у тюленя №4. В отличие от самца(тюлень №3), самка (тюлень №4) в течение 6 месяцев, предшествовавших экспериментам, выполняла движения по жестовым командам во время представлений в океанариуме. Это, по-видимому, и обусловило более низкий уровень дифференцировок. Результаты же самца свидетельствуют о том, что обучение распознаванию трехмерных зрительных раздражителей у этого тюленя достигнуто. Уже в первых опытах (№4 и №5) правильная дифференцировка наблюдалась более чем в 95% случаев, а в опытах №№ 13, 14 и 16 её уровень был близок к 100 процентам. Тем не менее, обучение самки также можно считать успешным: начиная со второй половины серии экспериментов и до их окончания результаты самки отличались высокой стабильностью (рис. 2, опыты №№ 11-18).
4.4 Выбор предметов по образцу
После обучения правилу выбора по образцу, тюлени из трех закрепленных на стенке вольера предметов выбирали, прикасаясь носом, один, соответствующий предъявленному образцу. Таких опытов с самкой (тюлень №4) проведено 10 (575 предъявлений, 61.48±5.79% правильных ответов), с самцом (тюлень №3) 22 (1135 предъявлений, 82.8±3.8% правильных ответов) (в опытах с 9-го по 13-й – не ниже 95%) (рис. 3).
При работе с самкой отмечено, что она не обозначает предмет, который висит дальше всех от помоста. В результате выбор осуществляется между двумя предметами. Если "дальний" из них именно тот, который соответствует предъявленному образцу, то выбор будет заведомо неправильным. Кроме того, самец был постоянно занят только в экспериментальной работе, самка участвовала в ней в то время, когда была свободна от представлений. На представлениях закреплялось и поддерживалось на высоком уровне выполнение рефлексов на сигналы-жесты.
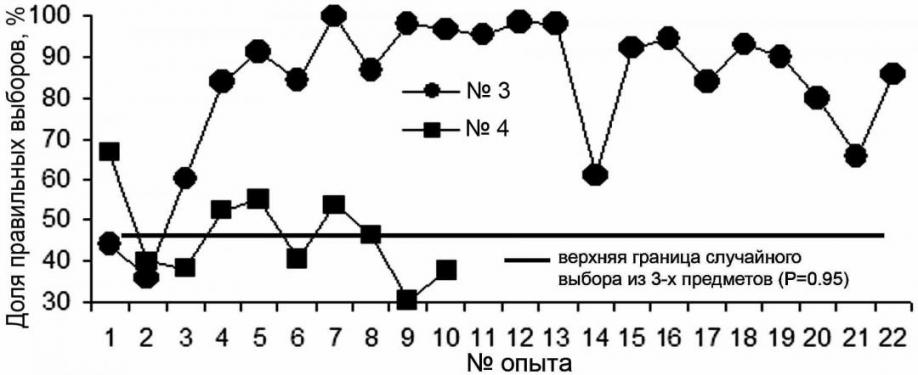
Рис. 3. Множественный выбор по образцу
Предъявление предметов в предыдущих экспериментах осуществлялось "псевдослучайно". Это могло привести к выработке у животных рефлексов не на присутствие определенных предметов в поле зрения, а на интервалы времени их предъявления или другие параметры. Поэтому заключительная серия экспериментов была проведена с использованием таблицы случайной комбинации трех чисел (рис. 4). Всего с самцом (тюлень №3) проведено 21 занятие со средним числом предъявлений 36. Тюлень получил 754 предъявления (258 – мяч, 264 – кольцо, 232 - труба). Среднее число правильных ответов составило (86.76±1.52%). В конце экспериментов (опыты 18 и 19) оно превысило 95%. Таким образом, и при псевдослучайном, и при случайном предъявлении предметов "выбор по образцу" осуществлялся с достаточной достоверностью.
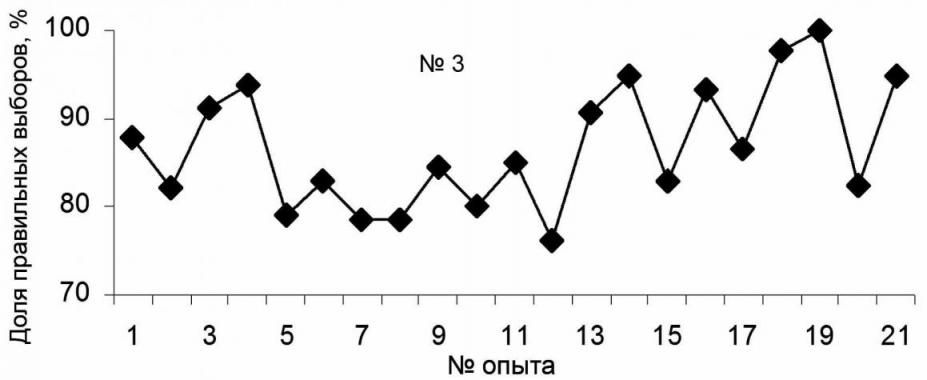
Рис. 4. Динамика обучения тюленя "выбору по образцу" из множества
Глава 5 РАСПОЗНАВАНИЕ ТЮЛЕНЯМИ СЛУХОВЫХ СТИМУЛОВ
5.1 Дифференцировка звуков различной частоты морским зайцем
В результате проведенных экспериментов морской заяц (тюлень №7) был обучен производить определенные движения в ответ на звуковые сигналы различной частоты. Звуки были выбраны в диапазоне вокализации морских зайцев: от 80 до 1500Гц (Свиженко, 1983). Для достижения 100%-й точности выполнения первого из вновь выработанных рефлексов потребовалось 244 предъявления раздражителя (700 Гц – "ладушки"), второго – 70 (1400 Гц – лежать), третьего – 111 (200 Гц – "стыдно"). Ускорение обучения реакции на второй сигнал более чем в 3 раза вполне объяснимо формированием "установки на обучение".
При предъявлении морскому зайцу звуковых раздражителей осуществлялась не только выработка новых условных рефлексов, но и дифференцировка раздражителей. На этапах эксперимента, когда вводили новый звук, происходила конкуренция между двумя, а затем и между тремя условными сигналами. Это выглядело как осуществление тюленем выбора – каким движением следует ответить и, вероятно, значительно повлияло на уровень правильных реакций. Средний уровень дифференцировки двух звуков составил 90.2±4.4%, трёх – 75.2±4.5%. Результат экспериментов, проведенных после четырёхмесячного перерыва, оказался в значительной степени неожиданным: сначала тюлень не вполне уверенно дифференцировал все 3 сигнала (опыт № 28, 200 Гц – 63%, 700 Гц – 75%, 1400 Гц – 100%), а затем (опыт № 29) продемонстрировал 100%-е их распознавание (рис. 5).
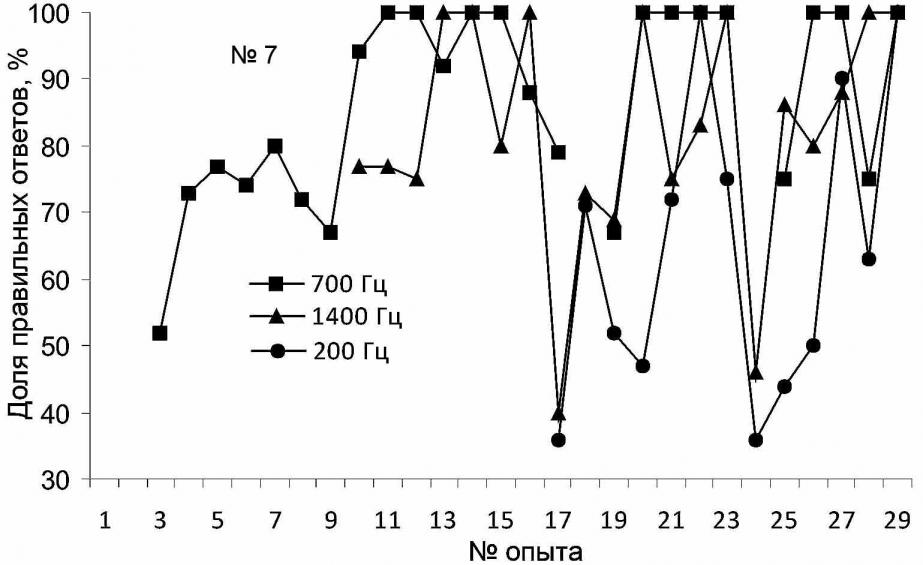
Рис. 5. Динамика формирования у морского зайца дифференцировки звуков различной частоты
5.2 Определение минимального частотного интервала между двумя разными звуковыми сигналами, распознаваемыми тюленями
5.2.1 Морской заяц
Первая пара звуков (200 Гц и 700 Гц), предложена тюленю для распознавания, поскольку ранее проведенные эксперименты позволяют нам, во-первых, не тратить время на выработку новых двигательных навыков, которые животное должно выполнять при предъявлении звукового раздражителя; а во-вторых, эту пару звуков морской заяц способен надежно дифференцировать.
Постепенно (опыты с 18 по 33) (рис. 6), тюлень стал распознавать звуки по относительному признаку "выше – ниже" уже при первом предъявлении новых пар звуков, что может свидетельствовать о высоком уровне установки на обучение и способности тюленя дифференцировать звуки не только по абсолютному, но и по относительному признаку.
Уменьшение числа правильных реакций на звуки 420 Гц и 480 Гц можно объяснить возможным отклонением в здоровье животного. Подобные спады, вплоть до отказа работать, встречаются при обучении всех тюленей в океанариуме. Определить причины этого, как правило, бывает трудно, за исключением случаев, когда явно присутствует отвлекающий фактор (например, сильный необычный запах, шум и т.п.), симптомы заболевания. Снижение пищевой мотивации наблюдается у животных, которые активно участвуют в представлениях океанариума. Если во время первого эксперимента заяц не участвовал в представлениях, то во время проведения данного эксперимента он был включен в ежедневные демонстрационные программы.
В опытах 54–59 частотный интервал между двумя основными звуками составляет 10 Гц, и животное их уверенно дифференцирует с 95%-м уровнем правильных ответов в двух опытах подряд (рис. 6, опыты 58-59). Поэтому в последующих экспериментах частотный интервал между распознаваемыми звуками уменьшали на 1 Гц.
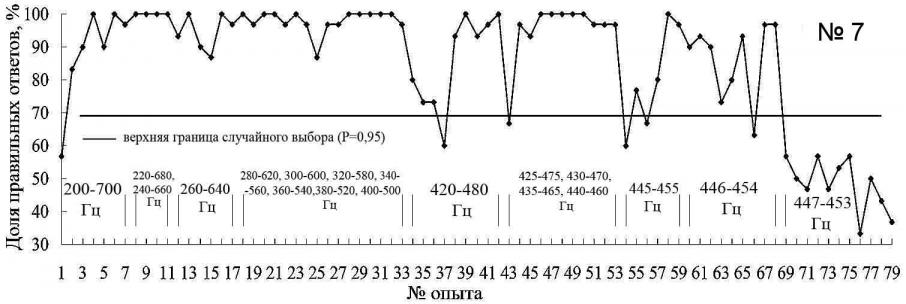
Рис. 6. Распознавание морским зайцем пар звуков разной частоты (над осью обозначены применявшиеся для дискриминации частоты пар звуковых сигналов)
В результате установлено, минимальный интервал между звуками, которые уверенно дифференцирует морской заяц, составляет 8 Гц. Результаты опытов 69–78 (рис. 6), где частотный интервал составил 6 Гц, показали, что морской заяц не дифференцирует эти звуки. При этом отмечено, что при предъявление звуковых раздражителей животное просто выполняет одно из двух движений. Во время опытов 69 – 77 (рис. 6) между предъявлениями звуковых раздражителей, животное вело себя беспокойно: двигалось по помосту, переворачивалось на спину, пытаясь получить пищевое подкрепление.
5.2.2 Гренландский тюлень
Работа с гренландским тюленем началась с выработки условных рефлексов "стыдно" и "ладушки" на голосовую команду тренера. Ранее животное было уже обучено выполнять эти действия по жестовым сигналам.
Первые 3 занятия предъявление звуков 200 и 700 Гц сопровождалось голосовой командой тренера (рис. 7). Два занятия животное уверенно дифференцировало эти две команды, на третьем занятии голосовая команда подавалась выборочно, и животное дифференцировало звуки с точностью 76.6%. С занятия №9 гренландский тюлень со 100%-ой уверенностью дифференцировал эти два звука.
В опытах 41-64 был отмечен спад правильных реакций на предъявляемые звуки. У животного наступил период линьки, что оказало существенное влияние на уровень физиологической активности и пищевой мотивации. Кроме того, сказалось влияние внешних факторов, как, например, в опыте №62 (рис. 7), когда при занятиях с гренландским тюленем присутствовали дети из детской художественной школы, которые рисовали тюленей.
При предъявлении пары звуков 425–475 Гц у гренландского тюленя, в отличие от морского зайца, не отмечено резкого снижения правильных реакций. Но также следует отметить, что животное уверенно дифференцирует пару 440 –460 Гц, тогда как при предъявлении пары 445 – 455 Гц 95%-ый уровень правильных реакций достигнут не был, а в опытах № 97, 104 и 105 составил 53.33% (рис. 7).
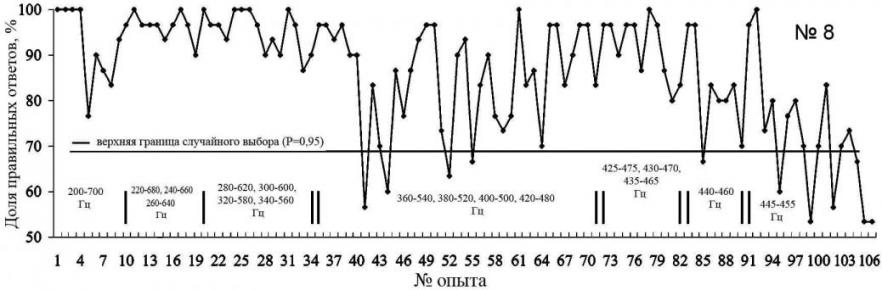
Рис. 7. Распознавание гренландским тюленем пар звуков разной частоты. (над осью обозначены применявшиеся для дискриминации частоты пар звуковых сигналов. Первые три занятия звук сопровождался голосовой командой тренера).
При предъявлении пары звуков 445 – 455 Гц животное выполняло одно из движений, стараясь получить подкрепление. Таким образом, минимальный интервал уверенного распознавания двух звуков гренландским тюленем составил 20 Гц.
5.2.3 Серый тюлень
К началу исследовательской работы самка серого тюленя №5 не имела опыта по выработке условных рефлексов на статические зрительные условные сигналы и звуки чистых тонов.
Тюлень №5 является постоянным участником цирковых представлений океанариума. В эксперименте использованы двигательные реакции "стыдно" и "ладушки", выполнявшиеся ранее при предъявлении жестового сигнала тренера. В первых опытах данной серии формировали связь: "звуковой сигнал – жест тренера – выполнение адекватного движения – подкрепление". Таких занятий было 7. Количество правильных ответов составило 100%, что, скорее всего, свидетельствует о прочности связи: жест – двигательный навык, а не о возникновении связи звук – двигательный навык.
Далее были проведены 24 занятия по предъявлению пары звуков 200 Гц и 700 Гц без подкрепления жестом. В опытах №8-31 животное получило 720 предъявлений звуковых раздражителей частотой 200 и 700 Гц. В целом процент правильных ответов на протяжении всего эксперимента был выше 50%, за исключением опытов № 12, 19, что возможно связано со слабой пищевой мотивацией тюленя во время. В дальнейшем тюлень показывал высокие результаты: 100% правильных ответов в опытах № 29-31, что позволяет говорить об уверенном дифференцировании животным звуков 200 Гц и 700 Гц по относительному признаку "выше–ниже". В опытах № 32-64 определяли величину минимального частотного интервала между двумя звуковыми сигналами, распознаваемыми серым тюленем. До конца эксперимента тюлень демонстрировал хорошую динамику и высокий уровень правильных ответов. Животное с высокой точностью дифференцировало последовательно сближаемые по частоте пары звуков. К 64 опыту серый тюлень уверенно различал звуки с интервалом в 260 Гц. Однако в декабре у животного начался период гона, и эксперимент пришлось прекратить. Тем не менее, полученные результаты свидетельствуют о том, что все исследуемые тюлени способны сравнивать и различать звуки по относительному признаку ("низкий"-"высокий"). Для распознавания очередной предъявляемой пары звуков тюленям необходимо от 1 до 5 занятий. Различение (распознавание, выбор) раздражителей по относительному признаку (выше-ниже, больше-меньше и т.п.) является важнейшим показателем их способности к обобщению (Зорина, Полетаева, 2001).
Уровень правильных ответов при выработке условных рефлексов на звуки 200 и 700 Гц при работе с молодым животным (гренландский тюлень) был значительно выше, чем у половозрелых зверей. Разницу в уровне дифференцировки звуковых раздражителей можно объяснить разным опытом работы по жестовым командам. Отсутствие большого багажа двигательно-пищевых условных рефлексов у молодых животных при осуществлении тюленем выбора – каким движением следует ответить также, вероятно, значительно повлияло на скорость обучения и уровень правильных реакций.
Уровень дифференцировки звуков может также зависеть не столько от возраста, уровня обученности животного и длительности его нахождения в условиях неволи, сколько от образа жизни. Так, морской заяц является бентофагом, а гренландский и серый тюлени – активными хищниками. Высокая чувствительность и избирательность слухового анализатора морского зайца обусловлены значительной ролью акустической информации в биологии данного вида (Белькович, Щекотов, 1990). Морские зайцы активно используют звуковую коммуникацию (звуковые трели разной длительности и частотного диапазона) в брачный период.
5.3 Дифференцировка морским зайцем сложных звуковых сигналов
К началу этой серии экспериментов морской заяц был обучен реагировать на жестовые и голосовые команды различными движениями. На начальных этапах обучения животного условные реакции вырабатывались на комплексные раздражители (жест + голос), затем жест был исключён. В последующем голосовые команды были записаны и транслировались посредством стационарной аудиосистемы.
Было проведено 7 опытов, в ходе которых прозвучало 279 сигналов (женским голосом – 130, мужским – 149). Впоследствии были проведены эксперименты с сигналами, синтезированными при помощи программы "Magic Gooddy". Компьютер озвучил 7 слов, которые использовались ранее.
В итоговом эксперименте из семи "машинных" сигналов морской заяц смог ответить правильно с точностью 100% только на "лежать" (табл. 1).
Таблица 1 Дифференцировка голосовых раздражителей морским зайцем
| Сигнал | Мужской голос | Женский голос | "Машинный" голос | ||||
| Число предъявле- ний | Число правильных ответов, % | Число предъяв-лений | Число правильных ответов, % | Число предъявле-ний | Число правильных ответов, % | ||
| Перевернись | 11 | 72.3 | 8 | 62.5 | 21 | 0 | |
| Ладушки | 5 | 100 | 5 | 100 | 19 | 0 | |
| Зубки | 7 | 100 | 7 | 100 | 18 | 0 | |
| Лежать | 5 | 100 | 8 | 100 | 19 | 100 | |
| В воду | 6 | 83.3 | 4 | 100 | 14 | 0 | |
| Посмотри.. | 4 | 50 | 3 | 100 | 12 | 0 | |
| Здравствуй | 6 | 100 | 8 | 75 | 18 | 6.6 | |
Сигналы значительно различались по частотному диапазону (табл. 2). Однако, поскольку уровень распознавания и "женских", и "мужских" сигналов высок, частота, вероятно, не является определяющим параметром при дифференцировке сложного сигнала.
Таблица 2 Средние частоты различных голосовых сигналов, Гц
| Сигнал | |||||||
| Голос | В воду | Здравст-вуйте | Зубки | Ладушки | Лежать | Перевернись | Посмотри... |
| Женский | 455.6 | 1213.6 | 711.1 | 2396.7 | 1939.1 | 2001.9 | 1875.3 |
| Мужской | 260.9 | 1096.1 | 749.6 | 1362.2 | 349.9 | 1503.5 | 458.0 |
| "Машинный" | 204.8 | 461.1 | 319.3 | 966.9 | 491.5 | 499.2 | 1294.7 |
Для различения звуков слуховым анализатором важна не только частота колебаний, но и их периодичность, т. е. длительность интервалов между максимальными и минимальными амплитудами колебаний.
Из представленных звуков наименьшие интервалы имеют сигналы, воспроизведённые женским голосом, в то время как "машинные" сигналы выделяются большой периодичностью и отличаются по этому показателю не только от женского голоса но и от мужского. Так, средняя периодичность сигнала "ладушки" составила: "машинный" голос – 15мс.; мужской голос – 7мс.; женский голос – 4мс.
Частотный и временной анализ элементов сигналов (слогов и отдельных звуков) свидетельствует также о том, что при распознавании их тюленями значительную роль играют также периодичность и длительность серий звуковых импульсов, составляющих эти элементы. Так, сигнал "ладушки", хорошо распознающийся в "человеческом" варианте, в "машинном" тюленем не распознается. Соотношение элементов этого раздражителя во всех трех его вариантах близки. Однако у "машинного" общая длительность первого элемента ("ла") значительно выше.
В то же время, у единственного уверенно распознанного "машинного" сигнала "лежать" и абсолютные, и относительные характеристики элементов сходны с "человеческими" (табл. 3).
Таблица 3 Временные и частотные характеристики элементов слова "лежать"
| Элемент раздражителя, характеристики | ||||||
| Голос | ЛЕ | Ж | АТЬ | |||
| t | f | t | f | t | f | |
| Женский | 112 | 251 | 65 | 2786 | 248 | 2885 |
| 26.4 | 15.3 | 58.3 | ||||
| Мужской | 130 | 136 | 72 | 2284 | 222 | 529 |
| 30.7 | 16.9 | 52.4 | ||||
| "Машинный" | 137 | 313 | 80 | 2817 | 282 | 373 |
| 27.5 | 16.0 | 56.5 | ||||
Примечание. t – время звучания в миллисекундах (нижняя цифра – в % от общей длительности раздражителя); f – частота, Гц.
Слушателем-человеком "машинные" голосовые сигналы распознаются правильно, несмотря на высокую периодичность звуковых колебаний, поскольку последние входят в состав элементов слов-понятий. Животные же, согласно современным представлениям, не обладают понятийным (вербальным) мышлением. Вероятно поэтому, начиная с некоторой достаточно высокой величины периодичности отдельные звуковые колебания и их группы различаются животным в качестве элементов, образующих новый, не известный ему сложный звуковой раздражитель. Таким образом, варьирование периодичности следования простых звуков в составе сложных звуковых сигналов позволяет, используя метод двигательных условных рефлексов, определять возможности слухового анализатора тюленей при различении звуков с разными частотно-временными характеристиками.
ВЫВОДЫ
1. Содержащиеся в океанариуме тюлени различаются по скорости и прочности выработки условных рефлексов. Эти различия обусловлены как индивидуальным опытом, так и психическими особенностями животных. С возрастом способность к обучению у тюленей не снижается. У них формируется "установка на обучение" с использованием условных раздражителей определенного типа.
2. При выработке зрительной дифференцировки у серых тюленей ведущую роль играет переключение внимания животных с экспериментатора на предъявляемый раздражитель. На поведение тюленей во время экспериментов могут существенно влиять любые, даже незначительные изменения обстановки т.е. играет роль вся совокупность внешних условий, "комплексный раздражитель".
3. Серые тюлени проявляют высокий уровень способности к обобщению при выборе предметов по относительному признаку одинаковости и подобия ("выбор по образцу" зрительных стимулов) и по различию – при распознавании звуковых стимулов.
4. При определении минимальной степени подобия плоских геометрических фигур, различаемых тюленями, результаты показанные серым тюленем значительно выше, чем результаты морского зайца, что связано с видовыми различиями в образе жизни и пищедобывательном поведении.
5. Установлены видовые различия в способности тюленей различать близкие по частоте звуки чистых тонов. Минимальный частотный интервал между звуками, распознаваемыми морским зайцем составил 8 Гц, гренландским тюленем 20 Гц, а серым тюленем 260 Гц.
6. При распознавании тюленями сложных звуковых стимулов ведущую роль играют периодичность следования и длительность составляющих их звуков чистых тонов.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ
Публикации в журналах из перечня ВАК РФ
1. Березина И.А., Кавцевич Н.Н. Распознавание серыми тюленями зрительных раздражителей. // Известия Самарского научного центра Российской Академии Наук. 2011. Т. 13 (39) № 1 (5). С. 1052-1056.
2. Березина И.А. Особенности питания представителей семейства настоящие тюлени в условиях неволи. // Вестник МГТУ, Т. 15. №3, 2012г. с. 505-508.
Публикации в других изданиях
3. Березина И.А. Формирование зрительной дифференцировки у серых тюленей. // Естественнонаучные проблемы арктического региона: Тез. докл. Второй региональной научной студенческой конференции. Мурманск, 15-16 мая 2001. Мурманск. 2001г. С. 10-11
4. Березина И.А., Михайлюк А.Л. Распознавание зрительных образов серыми тюленями // Естественнонаучные проблемы арктического региона: Тр. Второй региональной научной студенческой конференции (Мурманск, 15-16 мая 2001). Мурманск: МГПУ, 2001. C. 80-83.
5. Березина И.А., Михайлюк А.Л. Экспериментальное изучение поведения серых тюленей при выработке условных рефлексов на зрительные раздражители // Материалы ХХ юбилейной конференции молодых ученых Мурманского морского биологического института (Мурманск, апрель 2002). Мурманск: ММБИ КНЦ РАН, 2002. C. 10-11.
6. Михайлюк А.Л., Березина И.А., Степахно Г.В., Кавцевич Н.Н. Экспериментальное изучение поведения серых тюленей при различении зрительных стимулов // Морские млекопитающие Голарктики. Тез. докл. Второй международной конференции. Байкал, Россия 10-15 сентября 2002. Москва, 2002. С. 188-189.
7. Михайлюк А.Л., Березина И.А. Трошичев А.Р., Кавцевич Н.Н. Экспериментальное изучение поведения тюленей при различении зрительных и слуховых стимулов // Современные проблемы физиологии и экологии морских животных. Апатиты: Изд. КНЦ РАН, 2003. С.80-89.
8. Березина И.А., Кавцевич Н.Н. Распознавание тюленями зрительных и слуховых стимулов // Ученые записки МГПУ. Сер. биол. науки. 2004. Вып.1. С. 24-30.
9. Березина И.А., Кавцевич Н.Н, Калинин A.M. Определение минимального частотного интервала между двумя разными звуковыми сигналами, распознаваемыми представителями семейства настоящие тюлени. // Наука и образование [Электронный ресурс] / МГТУ. – электрон. текст дан. (18 Мб) – Мурманск: МГТУ, 2007. – 1 опт. компакт-диск (CD-ROM). С. 505-509.
10. Березина И.А., Калинин A.M., Кавцевич Н.Н. Изучение способностей серых тюленей к формированию зрительной дифференцировки по принципу ахроматичности. // Наука и образование [Электронный ресурс] / МГТУ. – электрон. текст дан. (18 Мб) – Мурманск: МГТУ, 2007. – 1 опт. компакт-диск (CD-ROM). С. 510-513.
11. Кавцевич Н.Н., Михайлюк А.Л., Березина И.А., Юрко А.С. Экспериментальное изучение поведения тюленей // Экспериментальные исследования морских млекопитающих в условиях Кольского залива. Апатиты: Изд. КНЦ РАН, 2007. С. 194-229.
12. Кавцевич Н.Н., Березина И.А. Биотехнические характеристики морских млекопитающих, перспективных для служебного использования. // Морские животные на службе человеку. Материалы научно-практического семинара 4 октября 2010 г., Североморск. Ростов-на-Дону: Изд. ЮНЦ РАН, 2011. С. 25-33.
13. Березина И.А., Калинин А.М., Кавцевич Н.Н. Различение настоящими тюленями геометрических фигур. // Наука и образование – 2011. [Электронный ресурс] / ФГОУВПО "МГТУ". электрон. текст. дан. (30 Мб) Мурманск: МГТУ, 2011. С. 541-548.
14. Березина И.А., Калинин А.М.,Трошичев А.Р. Распознавание звуков различной частоты настоящими тюленями. // Материалы XXIX конференции молодых ученых Мурманского морского биологического института, посвященной 140-летию со дня рождении Г.А. Клюге "Морские исследования экосистем Европейской Арктики" (г. Мурманск, май 2011 г.) Мурманск: Изд. ММБИ КНЦ РАН. С. 11-16.
15. Жарова Ю.А., Михайлюк А.Л. Березина И.А. Изучение суточной активности представителей семейства настоящие тюлени на примере кольчатой нерпы. // Успехи современного образования 2011. Москва "Академия Естествознания" 2011, № 8. С. 37.
16. Клапатюк А.М., Березина И.А. Применение метода этограмм при изучении поведения представителей семейства настоящие тюлени на примере гренландского тюленя. // Успехи современного образования 2011. Москва "Академия Естествознания" 2011, № 8. С. 42.
17. Березина И.А., Калинин А.М., Клапатюк А.М. Этологические наблюдения за ластоногими содержащимися в условиях океанариума и различение настоящими тюленями геометрических фигур с изменённой площадью сторон креста. // "Наука и образование - 2012 [Электронный ресурс] : материалы междунар. науч.-техн. конф., Мурманск, 2 - 6 апреля 2012 г. / Мурман. гос. тех. ун-т. – Электрон. текст. дан. (Мб). Мурманск : МГТУ, 2012. С. 319-321.
18. Березина И.А., Калинин А.М. Различение настоящими тюленями геометрических фигур с изменённой площадью сторон креста. // Биоресурсы и аквакультура: сб. тезисов Школы молодых ученых по морской биологии, Мурманск, 28-29 февраля 2012 г. Мурманск : Изд-во МГТУ 2012. С. 7-10.