

Взаимоотношения чумного микроба (yersinia pestis) и блох (siphonaptera) (на примере сибирских природных очагов чумы)
На правах рукописи
БАЗАНОВА
Любовь Петровна
ВЗАИМООТНОШЕНИЯ ЧУМНОГО МИКРОБА (YERSINIA PESTIS)
И БЛОХ (SIPHONAPTERA)
(на примере сибирских природных очагов чумы)
03.00.16 – экология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
доктора биологических наук
Иркутск – 2008
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы
Антропогенная трансформация ландшафтов, вовлечение территорий природной очаговости чумы в хозяйственную деятельность человека, существующая угроза террористических актов с использованием биологических объектов могут привести к пока непредсказуемым последствиям, что определяет необходимость усиления контроля за особо опасными инфекциями, в том числе, чумой (Anisimov et al., 2004; Смирнова, Кутырев, 2006; Eisen et al., 2006; Трухачев, Лебедева, 2006, 2007).
Природный очаг чумы – сложная экологическая система. Трофические связи в этой системе между чумным микробом (Yersinia pestis), носителями инфекции (теплокровными животными) и переносчиками (блохами – Siphonaptera) обеспечивают формирование трансмиссивного пути поддержания энзоотии (Bacot, Martin, 1914; Иофф, 1941; Бибикова, Классовский, 1974; Williams, Cavanaugh, 1983; Korenberg, 1989; Литвин, Коренберг, 1999; Иннокентьева и др., 1999; Атшабар, 1999; Анисимов и др., 2001).
Актуальность настоящей работы обусловлена следующим:
1. Сибирские природные очаги чумы (Забайкальский, Тувинский и Горно-Алтайский) в России являются северной окраиной Центрально-Азиатской зоны природной очаговости этой инфекции, основная часть которой расположена на территории Монголии и Китая (Сафонова, Саран, 1974; Голубинский и др., 1987; Попов и др., 2005). В Монголии и Китае ежегодно регистрируются вспышки заболевания людей чумой, и в последние годы прослеживается тенденция к увеличению числа заболевших (Tserennorov et al., 2003; Enkhbaatar.et al., 2003; Марамович и др. 2008).
На территории двух горных природных очагов Сибири (Тувинского и Горно-Алтайского) эпизоотии регистрируют ежегодно. При этом в Тувинском очаге с 1989 г. наблюдается трансформация сообществ носителей и переносчиков и расширение ареала чумного микроба (Галацевич, 1994; Вержуцкий, 2005). В Горно-Алтайском очаге происходит изменение структуры таксоценоза блох с возрастанием доли одних видов и сокращением численности других (Корзун, 2007).
2. Большинство культур чумного микроба, изолированных в сибирских очагах, получено от массовых видов блох. Так, в Тувинском очаге чумы на долю этих насекомых приходится 70,6 % штаммов возбудителя (Вержуцкий, 2005), Горно-Алтайском – 82,6 % (Корзун, 2007). Учитывая высокую естественную зараженность блох чумным микробом, считается более эффективным контролировать эпизоотическую ситуацию на территории природных очагов посредством борьбы с переносчиками, чем с носителями инфекции (Очиров и др., 1999; 2000; Вержуцкий и др., 2002; Матросов и др., 2004; Попов и др., 2004; Lorange et al., 2005; Никитин и др., 2005).
Результативность такого контроля зависит от глубоких знаний фауны, экологии, особенностей взаимоотношений блох с чумным микробом. Однако весь комплекс взаимоотношений чумного микроба и блох, влияющих на закономерность трансмиссии возбудителя в природных очагах Сибири, недостаточно исследован. Очевидна необходимость выявления таких особенностей, которые уточняют и расширяют наши представления о механизме энзоотии чумы. Изучение закономерностей системы паразит-хозяин, отношений между сочленами данной паразитарной системы (переносчик-чумной микроб) актуальны, поскольку напрямую связаны с проблемой эпидемиологического надзора и совершенствованием мероприятий по неспецифической профилактике этой инфекции.
Цель и основные задачи исследования
Цель работы – выявление особенностей взаимоотношений чумного микроба и блох, обеспечивающих трансмиссивный механизм поддержания энзоотии чумы в сибирских природных очагах.
Задачи:
1. Определить сезонную динамику процесса блокообразования у блох, их заражающей способности, роль самок и самцов этих эктопаразитов в трансмиссии чумного микроба.
2. Установить влияние абиотических факторов на алиментарную активность, смертность блох и их взаимоотношения с чумным микробом.
3. Рассмотреть возможность реализации и эпизоотологическую значимость второстепенных способов передачи чумного микроба блохами.
4. Оценить влияние межвидовых и межпопуляционных особенностей блох на их взаимоотношения с чумным микробом.
5. Установить особенности контактов между географически разобщенными популяциями чумного микроба и блох и исторически не контактировавшими с возбудителем чумы насекомыми и микробом.
6. Определить значение явления агрегированности чумного микроба в организме имаго блох для его передачи и сохранения насекомыми.
7. Выявить основные механизмы устойчивости сибирских природных очагов чумы, экологически связанные с блохами-переносчиками.
Научная новизна и теоретическая значимость
Впервые для сибирских природных очагов чумы установлена сезонная динамика блокообразования у блох и передачи чумного микроба этими насекомыми. Максимальные значения данных показателей в экспериментах совпадали по времени с пиком эпизоотии в природном очаге. Выявлено, что при кровососании на лабораторных животных (белых мышах), не имеющих выраженных сезонных различий в чувствительности к возбудителю чумы, динамика заражающей способности насекомых реализуется аналогично отмеченной при питании на длиннохвостом суслике. Это может указывать на коэволюцию эктопаразитов и чумного микроба, ритм взаимоотношения которых сохраняется при смене прокормителя.
Впервые показано, что процессы распространения и сохранения чумного микроба блохами связаны с полом насекомого: первый лучше осуществляется самцами, второй – самками. У самцов большинства массовых видов блох чаще и в более короткие сроки возникает «блок» преджелудка, чем у самок, и они активнее осуществляют передачу микроба.
Выявлены межпопуляционные различия во взаимоотношениях переносчиков с чумным микробом в Тувинском природном очаге. Более активное взаимодействие (по частоте формирования в организме насекомых бактериальных «глыбок») отмечены в парах (блоха – чумной микроб), происходящих с одного участка очаговости, или обитающих на участках со сходными биоценотическими характеристиками.
Впервые обнаружена возможность установления функциональных контактов между чумным микробом и блохами исторически не контактировавшими с возбудителем (с неочаговых по чуме территорий, внутривидовыми гибридами). Выявлена связь между уровнем флуктуирующей асимметрии билатеральных признаков у насекомых и их векторной активностью.
Впервые установлено, что инфицированные «неблокированные» особи Citellophilus tesquorum altaicus – основного переносчика в Тувинском природном очаге – способны более 600 суток сохранять чумной микроб и осуществлять его передачу длиннохвостым сусликам. При массовом питании «неблокированных» блох исходом такой передачи является развитие острой генерализованной формы инфекции у зверьков, или подострое течение заболевания с бактериемией, достаточной для заражения насекомых. У чувствительных к возбудителю чумы животных при укусах даже небольшого числа таких блох развивается острая форма инфекции с агональной септицемией.
Выявлено, что второстепенные способы передачи чумного микроба инфицированными насекомыми (внесение через поврежденную кожу зверька из раздавленных блох и их экскрементов, при поедании эктопаразитов грызуном) при определенных условиях могут быть не менее эффективными, чем укусы «блокированных» блох, приводить к заболеванию зверьков с генерализацией инфекции и обеспечивать дальнейшую трансмиссию возбудителя.
Впервые показано, что состояние агрегированности чумного микроба в организме переносчика, сопровождающееся изменением морфологии и ультраструктуры микроорганизма, обусловливает не только его способность к блокообразованию и дальнейшей трансмиссии, но и сохранение микроорганизма при неблагоприятных условиях окружающей среды.
На основании изложенных материалов могут быть обоснованы следующие теоретические положения. Первое – трансмиссия чумного микроба в природных очагах Сибири осуществляется преимущественно укусами «неблокированных» особей массовых видов блох, содержащих конгломераты микроба. Второе – агрегированная форма чумного микроба в организме блох – его особое состояние, определяющее как трансмиссию, так и сохранение микроорганизма в зимний сезон, что обеспечивает непрерывность его циркуляции в сибирских горных очагах чумы и их устойчивость.
Практическая значимость. Работа проведена в рамках выполнения семи научных тем Иркутского научно-исследовательского противочумного института Сибири и Дальнего Востока: «Разработка методов борьбы с носителями и переносчиками в плане оздоровления природных очагов Сибири» // № госрегистрации 81004192. – Иркутск, 1985; «Оценка современного состояния сибирских очагов чумы и изучение факторов их устойчивости при проведении оздоровительных мероприятий» // № госрегистрации 01860020801. – Иркутск, 1990; «Изучение роли Л-форм в персистенции чумного микроба в природных очагах Сибири» // № госрегистрации 01860020809. – Иркутск, 1990; «Оптимизация эпидемиологического надзора в сибирских природных очагах чумы» // № госрегистрации 01910033788. – Иркутск, 1995; «Совершенствование системы эпизоотологического мониторинга в сибирских очагах чумы в связи с обеспечением эпидемиологического благополучия населения» // № госрегистрации 01960000188. – Иркутск, 2001; «Оптимизация методов регуляции численности синантропных членистоногих (блохи, комары, тараканы) в условиях крупного города» // № госрегистрации 01980010186. – Иркутск, 2002; «Популяционно-экологические аспекты природной очаговости чумы в Сибири в связи с ее профилактикой» // № госрегистрации 012000113852. – Иркутск, 2005.
На основании выполненных экспериментальных работ рекомендовано применять противочумным станциям при проведении бактериологического исследования блох и клещей наряду со стандартными средами сухую питательную среду, предназначенную для выделения Л-форм чумного микроба (ЛС). Применение среды ЛС позволило повысить результативность эпизоотологического обследования сибирских очагов чумы на 25-30 %.
Материалы диссертации включены в нормативно-методические документы федерального и регионального уровня: «Методические рекомендации по повышению результативности исследований эктопаразитов в природных очагах чумы» (Иркутск, 1990), «Паспорт Тувинского природного очага чумы» (Иркутск, 2000), «Методические указания. Сбор, учет и подготовка к лабораторному исследованию кровососущих членистоногих – переносчиков возбудителей природно-очаговых инфекций» (М.: Федеральный центр госсанэпиднадзора Минздрава России, 2002), «Методические рекомендации по эпизоотологическому обследованию Тувинского природного очага чумы» (Иркутск, 2004), «Методические рекомендации по лабораторному разведению блох для научных исследований» (Саратов, 2007).
Основные положения, выносимые на защиту
1. В сибирских горных природных очагах чумы, несмотря на их различные характеристики по гостальности и векторности, основным механизмом поддержания энзоотии является трансмиссия чумного микроба инфицированными «неблокированными» блохами.
2. Блокообразование как специфический способ передачи чумного микроба блохами имеет выраженную сезонную динамику, обусловленную половыми и возрастными особенностями эктопаразитов и адаптацией к ним возбудителя. Наиболее активен этот процесс в пик эпизоотии в природном очаге.
3. Второстепенные способы передачи чумного микроба блохами могут быть в определенных условиях эффективными и играть существенную роль в циркуляции микроба в природных очагах Сибири.
4. Возможно установление функциональных контактов между чумным микробом и переносчиками из географически разобщенных популяций (из разных природных очагов), а также между возбудителем и блохами, никогда не контактировавшими с чумным микробом (насекомые с неочаговых территорий и межподвидовые гибриды блох). Адаптация Yersinia pestis к экологическим особенностям блох увеличивает способность микроба приживаться в их организме, о чем свидетельствует связь уровня ФА билатеральных признаков насекомых с частотой блокообразования.
5. Агрегированность чумного микроба является функцией его адаптации к организму блох и отражает векторную способность переносчиков.
Апробация работы. Результаты исследований представлены на научно-практической конференции «Профилактика природноочаговых инфекций» (Ставрополь, 1983); Х конференции Украинского общества паразитологов (Киев, 1986); X Съезде Всесоюзного энтомологического общества (Ленинград, 1989); региональном совещании противочумных учреждений по эпидемиологии, эпизоотологии и профилактике особо опасных инфекций (Уральск, 1989); межгосударственной научно-практической конференции «Организация эпиднадзора при чуме и меры ее профилактики» (Алма-Ата, 1992); региональной научно-практической конференции «Проблемы природноочаговых инфекций в Сибири и на Дальнем Востоке» (Чита, 1993); научной конференции «Проблемы инфекционной патологии в регионах Сибири, Дальнего Востока и Крайнего Севера» (Новосибирск, 1998); IY международном симпозиуме по блохам (Казахстан, Алматы, 1999); Первой международной юбилейной конференции «Актуальные проблемы инфектологии и паразитологии» (Томск, 2001); юбилейной научно-практической конференции «Эпидемиологическая безопасность на Кавказе», посвященной 50-летию Ставропольского НИПЧИ (Ставрополь, 2002); научной конференции «Проблемы инфекции в клинической медицине» и YIII съезде Итало-Россйского общества по инфекционным болезням (Санкт-Петербург, 2002); международной конференции «Проблемы современной паразитологии» и III съезде Паразитологического общества при РАН (Петрозаводск, 2003); конференции, посвященной 70-летию Противочумного центра (Москва, 2004); YII межгосударственной научно-практической конференции государств-участников СНГ «Чрезвычайные ситуации международного значения в общественном здравоохранении в решениях Санкт-Петербургского саммита «Группы восьми» и санитарная охрана территорий государств-участников СНГ» (Оболенск, 2005); I Всероссийском совещании по кровососущим насекомым (Санкт-Петербург, 2006); Всероссийской конференции с международным участием «Синантропизация растений и животных» (Иркутск, 2007); YIII межгосударственной научно-практической конференции государств-участ-ников СНГ «Международные медико-санитарные правила и реализация глобальной стратегии борьбы с инфекционными болезнями в государствах-участниках СНГ» (Саратов, 2007).
Личный вклад соискателя. Лабораторные эксперименты и полевые исследования, а также анализ материалов проведены при непосредственном участии автора. На полевые исследования затрачено 275 дней, на экспериментальные (работа с зараженным материалом) – 2006 дней, из них 739 непосредственно в природных очагах чумы.
Публикации. По теме диссертации опубликовано 72 работы, в том числе 32 в научных журналах и изданиях, рекомендуемых ВАК для публикации материалов докторских диссертаций, 40 – в других журналах, научных трудах и материалах международных, всесоюзных, региональных конференций (на русском, английском, китайском и монгольском языках).
Структура и объем диссертации. Диссертация состоит из введения, обзора литературы, семи глав собственных исследований, заключения, выводов и списка использованной литературы. Работа изложена на 325 страницах, иллюстрирована 35 рисунками и 60 таблицами. Список использованной литературы включает 488 источников, из них 121 в иностранных изданиях.
Благодарности. Автор признателен за помощь и поддержку администрации и сотрудникам зоолого-паразитологического, эпидемиологического отделов, лаборатории экспериментальных животных Иркутского научно-иссле-довательского противочумного института Сибири и Дальнего Востока, специалистам Тувинской противочумной станции. Искренне благодарю Т.И. Иннокентьеву, А.Г. Трухину, Ю.Д. Очирова, М.П. Маевского, А.Я. Никитина, А.Ф. Попкова, В.Т. Климова, Д.Б. Вержуцкого, Е.А. Вершинина, В.М. Корзуна, Е.Г. Токмакову, Е.А. Чипанина, Г.А. Воронову за разнообразную помощь в проведении исследований и подготовке диссертации.
СОДЕРЖАНИЕ РАБОТЫ
Глава 1. СОВРЕМЕННЫЕ ПРЕДСТАВЛЕНИЯ
О ВЗАИМООТНОШЕНИЯХ ЧУМНОГО МИКРОБА И БЛОХ
(ОБЗОР ЛИТЕРАТУРЫ)
Рассмотрены представления о природном очаге чумы как экологической системе (Павловский, 1947, 1960, 1964; Беклемишев, 1956; Балашов, 1972; Korenberg, 1982, 1989; Коренберг, 1983, 1985, 1986; Леви, 1985; Литвин и др., 1998; Литвин, Коренберг, 1999; Атшабар, 1999), особенностях взаимоотношений чумного микроба и блох (Pollitzer, 1954; Иофф, 1941; Бибикова, Классовский 1974; Алексеев, 1989; Ващенок, 1999; Анисимов и др., 2002; Anisimov et al., 2004; Hinnebusch, 2005), состоянии агрегированности (биопленке) микроба в организме блохи, а также значении этого явления в трансмиссии (Burrows, Bacon, 1956; Акиев, 1970, 1991; Бурделов, 1987; Ващенок, 1988; Брюханова и др., 1989, 1999; Hinnebusch et al., 1998, 2002; Jarret et al., 2004; Duplantier et al., 2005; Erickson et al., 2006).
В 80-х годах прошлого века факты длительных межэпизоотических периодов в отдельных природных очагах чумы, одномоментности возникновения эпизоотий на разных участках, регистрации одиночных заражений носителей стали вступать в противоречие с традиционными представлениями о постоянном сохранении и непрерывной циркуляции чумного микроба в паразитарной системе. Это привело к обоснованию концепции «первичных» заражений, не связанных с трансмиссией (Попов и др., 1979; Солдаткин, Иванов, 1980; Мастюков и др., 1980; Солдаткин и др., 1984), и мысли о необходимости пересмотра традиционных взглядов на передачу чумы блохами как основного фактора поддержания энзоотии (Солдаткин, Руденчик, 1988). Кризис идеи трансмиссивного механизма породил большое количество гипотез, основанных как на внутривидовой изменчивости чумного микроба (Дунаев и др., 1981, 1982; Дятлов, 1988, 1989; Ларина и др., 1992), так и «непаразитарном» механизме энзоотии (Солдаткин, Руденчик, 1988; Домарадский, 1993; Слудский, 1998; Попов, 2002; Слудский и др., 2003). Факты зараженности нематодами личинок и имаго блох (Чумакова, Товканев, 1978; Чумакова, Козлов, 1983, 1984) привлечены для объяснения хранения и передачи возбудителя из почвенного биоценоза теплокровным животным по цепочке: матрикс биопленки чумного микроба – личинка нематоды – личинка блохи – грызун (Попов и др., 2006; Попов и др., 2007; Попов и др., 2008). Большинство гипотез, объясняющих длительные межэпизоотические периоды в отдельных очагах чумы нетрансмиссивными способами передачи возбудителя, до настоящего времени не получили прямого подтверждения или доказательства. Напротив, многочисленные экспериментальные данные, результаты эпизоотологических наблюдений в природных очагах свидетельствуют об определяющей роли блох в поддержании чумной энзоотии и обосновывает необходимость дальнейших исследований особенностей их взаимоотношений с чумным микробом (Леви, 1985; Бурделов, 1987; Сулейменов, 1995; Кокушкин, 1995; Анисимов и др., 2002; Eisen et al., 2006, 2007; Wilder et al., 2008).
Даны основные характеристики сибирских природных очагов чумы, особенности их биоценотической структуры и эпизоотических проявлений на очаговой территории (Голубинский и др., 1987; Вершинин, 1993; Иннокентьева, 1997; Вержуцкий, 2005; Корзун, 2007).
Глава 2. МАТЕРИАЛ И МЕТОДЫ
Районы работ и объекты изучения. Полевые исследования проведены в 1974-1994 гг. на базе стационаров и эпидотрядов Читинской, Тувинской и Алтайской противочумных станций (ПЧС). Экспериментальные исследования выполнены в 1985-2007 гг. на базе паразитологического, эпидемиологического отделов и лаборатории экспериментальных животных Иркутского научно-исследовательского противочумного института (НИПЧИ). Основной объем работ в условиях природного очага выполнен на базе экспериментальной лаборатории Тувинской ПЧС вблизи пос. Мугур-Аксы Республики Тыва.
В экспериментах использованы лабораторные культуры блох и насекомые природных популяций. Блохи диких грызунов получены из биотопов долин рек Каргы и Барлык Тувы, природных стаций Юго-Восточного Забайкалья (падь Крементуй, район Торейских озер), Ольхонского района (Тажеранская степь), окрестностей пос. Усть-Орда Усть-Ордынского Бурятского автономного округа, Республики Бурятия (Тункинская долина, окрестности пос. Никольское). Опыты также проведены с Xenopsylla cheopis как классическим объектом экспериментальных исследований. Для заражения эктопаразитов, мелких млекопитающих и лабораторных животных использовали штаммы Y. pestis subsp. pestis, Y. pestis subsp. altaica, Y. pestis subsp. ulegeica из природных очагов Сибири (Тувинского, Горно-Алтайского, Забайкальского) и Монголии (Баян-Улэгейский и Хэнтейский аймаки). Поставлено 108 опытов, использовано 25500 блох 13 видов и подвидов, 1746 зверьков четырех видов и 13 штаммов чумного микроба.
Методы исследований взаимоотношений чумного микроба с блохами. Лабораторное разведение эктопаразитов для экспериментальных исследований осуществляли в инсектарии НИПЧИ и на базе Тувинской ПЧС (Методические рекомендации..., 1983; Кондрашкина и др., 1984). В качестве прокормителей блох использовали золотистых хомячков и белых мышей.
В опыты брали в основном «молодых», питавшихся после выплода имаго. Применяли два способа заражения переносчиков чумным микробом. При естественном способе блох инфицировали на длиннохвостых сусликах, монгольских пищухах, плоскочерепных полевках, белых мышах в период развития у них агональной чумной бактериемии. При искусственном заражении на биомембране для кормления насекомых использовали смесь равных частей дефибринированной крови морской свинки и 2 млрд. взвеси чумного микроба из 48-часовой агаровой культуры, выращенной при 28 С. Между подкормками насекомых содержали в стерильном песке при соответствующей задачам опыта температуре и относительной влажности воздуха 80-93 %.
Взаимоотношения чумного микроба и блох оценивали по доле особей с «блоком» преджелудка от количества насекомых при первой подкормке, количеству всех особей за подкормку со сформировавшимися бактериальными «глыбками-конгломератами», частоте передачи чумного микроба животным, использованным для подкормок насекомых. Алиментарную активность определяли по доле пивших особей от числа живых, а смертность – по доле мертвых от общего количества эктопаразитов при каждой подкормке.
Сезонные особенности трансмиссии чумного микроба блохами изучали при разной температуре содержания насекомых, близкой к естественной температуре воздуха во время периода исследования. В опытах, поставленных при температуре от 5° до 27° С, использованы насекомые разных, условно выделенных календарных возрастов: «перезимовавшие», инфицированные летом или осенью предыдущего года и зимовавшие без прокормителя в условиях искусственной норы длиннохвостого суслика, и «молодые» – выплода и инфицирования текущего года. Преимущественно в эти эксперименты взяты особи II и III физиологических возрастов по шкале В.Н. Прокопьева (1958).
Активность трансмиссии оценивали по доле «блокированных» блох и всех особей, содержащих конгломераты микроба, относительному количеству инфицированных сусликов и среди них с генерализованной формой инфекции. Инфекциозность укусов «блокированных» блох определяли путем ежедневного кормления 1-3 таких особей на суслике в течение 1,5-2 часов, используя при каждой подкормке интактного зверька. В отдельном опыте эктопаразитов содержали в термостате при 25-27 С, исходя из данных О.Ф. Пауллер и В.Е. Мамаева (1954), что в центре жилого гнезда даурского суслика температура колеблется от 16 до 34 С, а в среднем равняется 26,7 С.
Изучение длительности сохранения чумного микроба C. tesquorum altaicus проводили по разработанной нами методике (Базанова, 1993). Зараженных имаго помещали в стаканчики-гильзы из органического стекла, дно и крышка которых выполнены из асбестового стерилизующего фильтра, обеспечивающего свободный доступ воздуха. «Гильзы» закладывали в ниши бункера, имитирующие гнездовую камеру суслика. Температуру и влажность воздуха измеряли перед началом опыта, в середине и в момент изъятия насекомых из «гнезда». За время проведения эксперимента температура колебалась от +2 С до -6 С. Между подкормками блох содержали: осенью и весной при 5-6 С, летом при 8-10 С. Общая продолжительность эксперимента составила 692 дня (с 14 сентября 1987 г. по 7 августа 1989 г.).
Заражение длиннохвостого суслика алиментарным способом проводили путем нанесения на слизистую рта зверька суспензии из 1-5 инфицированных блох. Возможность заражения суслика возбудителем чумы через поврежденную кожу устанавливали, втирая раздавленных насекомых или их экскременты в предварительно скарифицированную кожу зверька. Исследовали длительность сохранения чумного микроба в инфицированном экскрементами и трупами эктопаразитов субстрате гнезда. Для этого использовали смесь из равных частей песка и мелких опилок, где содержали зараженных насекомых в период между подкормками. Такой субстрат хранили в течение 8-9 месяцев в специальных нишах лаборатории-бункера, имитирующих нежилые гнезда суслика, а затем использовали для заражения сусликов в виде подстилки. Суспензии из трупов блох высевали на экспериментальные серии сред: УДС (универсальная диагностическая среда) и ЛС (среда для выделения Л-форм чумного микроба). Суспензию из субстрата высевали на среду УДС; этим же материалом заражали белых мышей подкожно или внутрибрюшинно.
Изучение межпопуляционных особенностей взаимоотношений переносчиков с чумным микробом. Исследование проведено на двух массовых видах блох длиннохвостого суслика C. tesquorum altaicus и Rhadinopsylla li transbaikalica из Каргинской и Барлыкской природных популяций Тувинского очага чумы. Взаимоотношения с чумным микробом оценивали по уровню инфицирования насекомых, накоплению в их организме микроба в агрегированном состоянии и образованию «блока» при заражении и подкормках на длиннохвостом суслике. Кроме того, изучали особенности взаимоотношений C. tesquorum altaicus лабораторной популяции, полученной от блох с Каргинского участка очаговости, и чумного микроба, изолированного в Саглинском и Каргинском участках Тувинского очага (штаммы Y. pestis И-3327 и И-3428) при искусственном заражении и подкормках блох на белых мышах.
При исследовании уровня флуктуирующей асимметрии (ФА) у C. tes-quorum внутри каждого варианта опыта сопоставляли две группы блох: одна – особи со сформировавшимся «блоком» преджелудка, другая – те, у которых в течение опыта «блок» не был отмечен. Оценивали признаки: число щетинок на эпистерне (1), метэпимере (2), наружной поверхности бедра (3), внутренней поверхности бедра (4), наружной поверхности голени (5) с правой и левой стороны тела имаго. Уровень ФА характеризовали через показатель дисперсии разности в проявлении признака с двух сторон тела (d2), а также путем расчета доли асимметричных имаго в проанализированной выборке.
Изучение особенностей взаимоотношений блох и чумного микроба из географически разобщенных популяций проведено на массовых видах эктопаразитов монгольской пищухи, длиннохвостого суслика и штаммах возбудителя, полученных из разных природных очагов.
Изучение особенностей взаимоотношений чумного микроба с блохами, исторически не имевшими контакта с возбудителем, выполнено на имаго C. tesquorum и Frontopsylla luculenta с неочаговых территорий, а также гибридах F1, F3, F4 двух подвидов C. tesquorum (C. tesquorum altaicus и C. tesquorum sungaris). В серии экспериментов изучали способность насекомых к инфицированию, «блокированию» чумным микробом и его передаче зверькам.
Исследование инфицированных блох проводили:
1. Визуально путем микроскопирования после каждой подкормки на животном выявляли блох с бактериальными «глыбками», частичными и полными «блоками». Учет средней за подкормку доли насекомых, содержащих микроб в различной степени агрегированности, позволил провести сравнительный анализ динамики взаимоотношений переносчика и возбудителя.
2. Исследовали содержимое желудочно-кишечного тракта блох путем свето-оптического, фазово-контрастного и электронного микроскопирования. Мазки для исследования на светооптическом уровне готовили по методике А.П. Бейер с соавт. (1974). Ультратонкие срезы преджелудка, приготовленные согласно методическим рекомендациям (Волгоград, 1983) и (Саратов, 1989), просматривали в электронном микроскопе Jem 100 SX (JeOL, Япония). Проводили фильтрацию суспензии из содержимого желудков инфицированных блох через мембранные фильтры с диаметром пор 0,45 мкм и высев полученных фильтратов на стандартные питательные среды, а также среду для культивирования Л-форм чумного микроба.
3. Для детекции микроба в насекомых использованы бактериологический и молекулярно-биологический – постановка полимеразной цепной реакции (ПЦР) методы. Подробно методики подготовки блох к исследованию и постановки ПЦР изложены в работах (Балахонов и др., 1998, 1999, 2000).
4. Взаимоотношения блох и чумного микроба исследовали с учетом эколого-физиологических особенностей насекомых: сроков важнейших сезонных явлений в их жизни, возрастного и полового состава опытных групп, алиментарной активности, смертности, продолжительности жизни.
Исследование зверьков, использованных для кормления инфицированных насекомых. Манипуляции с животными проводили с соблюдением правил, принятых Европейской конвенцией по защите позвоночных животных, используемых для экспериментальных и научных целей (Страсбург, 1986). Факты передачи чумного микроба животным устанавливали бактериологическим (посев органов на стандартные среды или среду для Л-форм чумного микроба), биологическим (заражение биопробы суспензией из органов животных) и серологическим (реакция пассивной гемагглютинации – РПГА, реакция нейтрализации антител – РНАт и реакция нейтрализации антигена – РНАг) методами исследования. Часть опытных животных, на которых кормили «перезимовавших» блох и без видимого «блока», дополнительно исследовали в иммунологических тестах: определяли показатель повреждения нейтрофилов (ППН) по методу В.А. Фрадкина (1962), исследовали органы с помощью радиоиммуного анализа (РИА).
Статистическая обработка экспериментальных данных. Статистическую обработку проводили с помощью стандартных методов (Рокицкий, 1967; Закс, 1976; Поллард, 1982; Боровиков, Ивченко, 2000; Боровиков, 2001; Никитин, Сосунова, 2003) с применением программы «Excel». Различия между двумя группами по средним показателям оценивались с применением критерия Стьюдента, по изменчивости – F критерия. Использованы методы регрессионного, дисперсионного и корреляционного анализов и метод множественных сравнений по Шеффе. В работе приняты стандартные уровни значимости: * – P<0,05; ** – P<0,01; *** – P<0,001.
ГЛАВА 3. ВЛИЯНИЕ СЕЗОНА ГОДА И ПОЛОВЫХ РАЗЛИЧИЙ БЛОХ НА ВЗАИМООТНОШЕНИЯ С ЧУМНЫМ МИКРОБОМ
Сезонная динамика взаимоотношений чумного микроба и блох. Для Тувинского природного очага чумы свойственна сезонность эпизоотического процесса, связанная с эколого-физиологическими особенностями основного носителя – длиннохвостого суслика, (в том числе, с сезонным колебанием чувствительности зверька к чумному микробу) и основного переносчика – блохи C. tesquorum altaicus (Обухов, 1969; Устюжина и др., 1975; Крюков, 1984; Олькова и др., 1984; Равдоникас, 1985)
Изучена в эксперименте заражающая способность C. tesquorum altaicus в различные сезоны года при подкормках блох на суслике (рис. 1).
 | Рис. 1. Сезонная динамика заражающей способности C. tesquorum altaicus при питании на длиннохвостых сусликах: |
При групповых подкормках доля «блокированных» особей (рис. 1) достоверно увеличивалась от весны к лету (по сравнению с началом лета t=2,09, P<0,05; с серединой лета t=2,86, P<0,01). Частота заболеваний грызунов весной и в начале лета была на одном уровне, но отмечена тенденция к ее увеличению от начала к середине лета. В июле-августе этот показатель по сравнению с весной значительно вырос (t=2,18, P<0,05). Частота генерализации инфекции у сусликов также возросла в июле-августе (в среднем 43,1 %) по сравнению с весенне-раннелетним периодом (в среднем 13,2 %).
Инфекциозность укусов единичных «блокированных» блох (отношение числа заболевших животных к общему числу зверьков, на которых кормили особей с «блоком») также изменялась в зависимости от сезона года (табл. 1).
Таблица 1
Инфекциозность укусов «блокированных» C.tesquorum altaicus
в различные периоды эпизоотического сезона
| № опыта | Время проведения опыта | Количество блох | Коли- чество сусликов | Заболело чумой | Показатель инфекциозности |
| 1 | III декада апреля – май | 1-3 | 10 | 1 | 0,10 ± 0,095 |
| 2 | Июнь – I декада июля | 1-3 | 10 | 3 | 0,30 ± 0,145 |
| 3 | III декада июля – август | 1-2 | 10 | 7 | 0,70 ± 0,145 |
| 4 | То же | 1-2 | 10 | 6 | 0,60 ± 0,155 |
| 5 | Сентябрь | 1-3 | 10 | 5 | 0,50 ± 0,158 |
Самые высокие значения данного показателя отмечены в III декаде июля-августе. Различия между результатами, полученными весной (май) и в середине лета, высоко достоверны (t=2,74, P<0,05 для первого опыта, t=2,34, P<0,05 для второго и t=2,85, P<0,01 для двух опытов). Кроме того, прослеживается тенденция к возрастанию инфекциозности укусов особей с «блоком» в июле-августе и по сравнению с началом лета (июнь-I декада июля) (табл. 1).
Сезонная динамика взаимоотношений чумного микроба с C. tesquorum altaicus исследована при подкормках на неспецифичном для этого вида блох прокормителе – белой мыши (рис. 2).
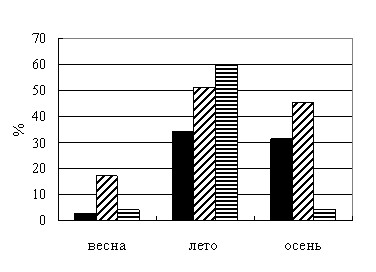 | Рис. 2. Сезонная динамика векторной способности C. tesquorum altaicus при питании на белых мышах: |
Блох, происходящих с Каргинского участка очаговости, инфицировали штаммами чумного микроба, изолированными на Каргинском (И-3327) и Саглинском (И-3428) участках Тувинского очага.
В данных экспериментах все показатели, характеризующие векторную способность блох, повышались от весны к лету (рис. 2) аналогично опытам на суслике (рис. 1). Однофакторный дисперсионный анализ выявил существенное значение сезона в частоте формирования конгломератов микроба в организме блох (F=13,84, P<0,001 для штамма И-3327; F=16,43, P<0,001 для штамма И-3428). Летом численность имаго, содержавших конгломераты возбудителя, выше, по сравнению с весной и осенью. Различия по этому показателю для штамма И-3428 существенны между весной и летом (t=5,50, P<0,001), а также летом и осенью (t=3,17, P<0,01). Для штамма И-3327 различия достоверны между весной и летом (t=5,08, P<0,001).
Частота передач микроба самками в течение опытов сохранялась примерно на одном уровне. Самцы передавали возбудителя летом чаще, чем весной (t=2,76, P<0,01), осенью эффективность передачи у них не снижалась (t=1,06, P>0,05). Достоверное возрастание частоты передач от весны к лету (t=2,06, P<0,05) выявлено для обоих полов. Очевидно, что передача чумного микроба C. tesquorum altaicus как сусликам, так и белым мышам, активизируется от весны к лету, в основном за счет векторной способности самцов.
Результаты опытов указывают, что на заражающую способность насекомых оказывает существенное влияние сезон года. При этом данная способность не всегда напрямую зависит от температурного режима содержания и частоты подкормок инфицированных эктопаразитов, и, возможно, обусловлена какими-то другими, не установленными биологическими факторами. В любом случае этот феномен свидетельствует об адаптации возбудителя, циркулирующего в Тувинском природном очаге, к жизненному циклу основного переносчика. Их взаимодействие имеет выраженную сезонную динамику, которая проявляется даже при питании блох на неспецифичном прокормителе.
Проанализированы особенности накопления конгломератов чумного микроба в организме Amphipsylla primaris primaris, блокообразования, алиментарной активности и смертности этих насекомых при питании на плоскочерепной полевке, а также влияния сезона года на эти показатели.
Всего «блокированных» A. primaris primaris без учета пола выявлено осенью 10,2 %, весной – 5,4 % (t=2,34; P<0,05). Осенью образование «блока» происходило у самок и самцов в одни и те же сроки. От общего количества взятых в опыт блох доля таких самок составила 11,6 %, самцов – 8,5 % (t=1,29; P>0,05). Весной у самцов блокообразование начиналось и заканчивалось раньше, чем у самок. Самцов с «блоком» отмечено 7,5 %, самок – 4,5 % (t=1,19; P>0,05). Осенью «глыбки» и «блоки» у блох формировались примерно в те же сроки, что и весной, но количество самок с «блоком» было выше (t=2,87; P<0,01). Частота «блокирования» самцов по сезонам не различалась.
Образование бактериальных «глыбок» осенью как у самок, так и у самцов отмечали примерно в два раза чаще, чем весной (табл. 2). Доля самцов с «глыбками» превысила долю самок в 1,5-2 раза в оба сезона. Но в среднем «блокированных» самцов весной отмечено почти в три раза больше, чем самок, а осенью – в два раза меньше. Среди блох с частичным «блоком» в оба сезона преобладали самки. В осеннем опыте таких самок в среднем выявляли почти в 6 раз больше, чем весной, а самцов отмечали исключительно осенью.
Таблица 2
Средняя доля A. primaris primaris, содержавших конгломераты чумного микроба, от числа пивших за подкормку особей (%)
| Время проведения опыта | Самки | Самцы | ||||||
| с глыбками | с частичным блоком | с полным блоком | всего | с глыб- ками | с частичным блоком | с полным блоком | всего | |
| Осень 1982 г. | 28,6 | 8,7 | 3,9 | 41,2 | 42,1 | 5,1 | 2,1 | 49,3 |
| Весна 1983 г. | 13,3 | 1,6 | 0,9 | 15,8 | 27,3 | 0 | 2,5 | 29,8 |
Более частое разрушение полных «блоков» у самок, вероятно, связано с большим количеством насасываемой ими крови, по сравнению с самцами. Причем осенью эта тенденция усиливается у блох обоих полов. Видимо, осенью микробу более «выгодна» адаптация к организму блохи для сохранения в холодный период, чем образование «блока» и дальнейшая трансмиссия.
На алиментарную активность и смертность A. primaris primaris оказывали влияние сезон года и пол насекомого. Активность кровососания весной была более стабильна (у самок 2=103,9, у самцов 68,7), чем осенью (у самок 2=247,4, F=2,38, P<0,05, у самцов 262,6, F=3,82, P<0,01). Весной гибель самцов выше, чем самок (t=5,32; P<0,001), а по сравнению с осенью, она увеличилась вдвое (t=3,51; P<0,001). Гибель самок в эти сезоны не различалась.
Таким образом, способность чумного микроба к блокообразованию у A. primaris primaris разного пола от весны к осени может изменяться противоположным образом, что обусловлено различиями в сезонных изменениях физиологических показателей блох – активности кровососания и смертности.
Влияние пола блох на их взаимоотношения с чумным микробом. Предполагалось, что выраженный половой диморфизм у блох должен отражаться на способности к сохранению и распространению чумного микроба самками и самцами. Для выявления таких половых различий проведен анализ данных многолетних экспериментальных исследований.
Частота и динамика образования «блока» преджелудка. Во всех опытах с X. cheopis «блоки» у самцов формировались чаще, чем у самок (рис. 3).
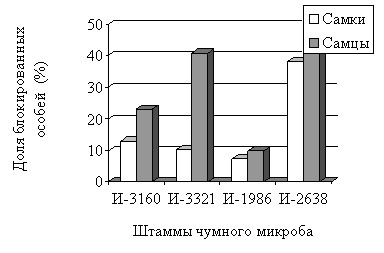 | Рис. 3. Блокообразование у самок и самцов X. cheopis |
Необходимо отметить, что смешанные группы X. cheopis (самки + самцы) заражали алтайскими (И-3160 и И-3321) и тувинским (И-2638) штаммами чумного микроба. В опытах с забайкальским штаммом (И-1986) группы формировали раздельно по полу. При разных условиях опытов более активно образование «блоков» происходило у самцов X. cheopis.
У самцов большинства исследованных видов блох из природных популяций Тувинского очага выявлено аналогичное с самками или более активное формирование «блоков» (рис. 4).
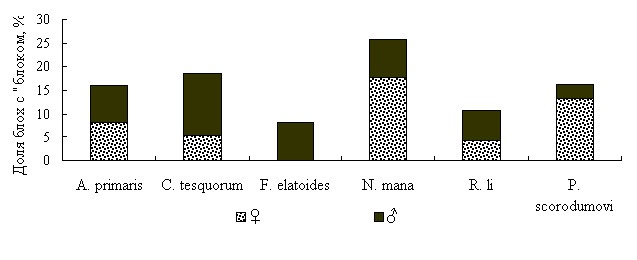 Рис. 4. Блокообразование у самок и самцов массовых видов блох из Тувы
Рис. 4. Блокообразование у самок и самцов массовых видов блох из Тувы
Представляет интерес, что доля «блокированных» самцов C. tesquorum altaicus при подкормках на суслике в июле-августе значительно превышала долю таких самок. В одном опыте выявили 13,8 % самцов и 5,4 % самок с «блоком», во втором – 7,7 % и 1,3 %, в третьем – 24,7 % и 3,7 %. Половые различия по этому признаку достоверны (для первого и второго опытов t=1,97, P<0,05, для третьего t=4,34, P<0,001). Кроме того, во всех проведенных на суслике опытах процесс блокообразования у самцов начинался и заканчивался раньше, что говорит о более коротком его периоде у этого пола.
При подкормках C. tesquorum altaicus на мышах в среднем за все опыты (без учета сезона и штамма Y. pestis) доля «блокированных» самок составила 11,4 %, самцов – 2,5 % (t=3,03, P<0,01). Летом процесс блокообразования активизировался у особей обоих полов, однако доля самцов с «блоком» была меньше, чем самок. То есть наблюдали ситуацию прямо противоположную отмеченной при кормлении блох на суслике, что связано с использованием неспецифичного для блох прокормителя – белой мыши.
Исследования показали, что у блох существуют явные половые различия, как в частоте, так и динамике процесса блокообразования. Эти показатели существенно зависят не только от видовых особенностей эктопаразитов и используемых прокормителей, но и сезона года, а также условий содержания самок и самцов в эксперименте (совместно или раздельно). Так, в опытах с C. tesquorum altaicus, проведенных в сроки, совпадающие с активизацией эпизоотического процесса в очаге (июль-август) и на естественном для данного вида прокормителе (длиннохвостый суслик), «блок» преджелудка отмечали у самцов значительно чаще, чем у самок. В то время как среди самцов, подкармливаемых кровью белых мышей, ни в одном случае не обнаружено больше «блокированных» особей, чем среди самок. На естественном прокормителе у самцов C. tesquorum altaicus зарегистрированы и более ранние сроки формирования «блоков», что в большинстве случаев характерно для особей этого пола и других видов. При совместном содержании полов значительно уменьшается продолжительность жизни блох, особенно, самцов (Цэрэнноров, 1999), что, видимо, снижает способность последних противостоять неблагоприятным факторам, в том числе и инфицированию чумным микробом. Видимо, блокообразование у самцов более зависимо от условий среды, и отражает меньшую степень адаптации микроба к особям этого пола.
Частота передачи чумного микроба. Различия между полами в частоте трансмиссии у C. tesquorum altaicus, F. luculenta luculenta, Х. cheopis изучали в опытах, где экспериментальные группы насекомых формировали раздельно из самок и самцов (табл. 3).
Таблица 3
Передача чумного микроба самками и самцами блох (без учета сезона года)
| Вид блохи | Прокормитель | Штамм Y. pestis | Количество передач (%) | ||
| самки | самцы | t | |||
| C. tesquorum altaicus | Длиннохвостый суслик | И-3226 | 38,5 | 46,2 | 0,40 |
| Белая мышь | И-3327 | 56,9 | 30,7 | 1,54 | |
| И-3428 | 59,5 | 43 | 0,98 | ||
| F luculenta luculenta | -«- | И-1996 | 35,7 | 78,6 | 2,29* |
| X. cheopis | -«- | -«- | 30 | 50 | 0,91 |
При подкормках C. tesquorum altaicus на суслике в июле-августе несколько большую заражающую способность наблюдали у самцов, но различия между полами в передаче возбудителя недостоверны. В опытах с белыми мышами (без учета сезона), наоборот, самки более активно передавали возбудителя. Существенные половые различия в передаче чумного микроба мышам выявлены весной (самки в 51,6 % случаев, самцы – в 14,3 %), летом и осенью они незначительны. Летом самцы передавали возбудителя достоверно чаще (43,2 %), чем весной (t=2,76, P<0,01), осенью (55,6 %) по сравнению с летом их эффективность как переносчиков не снижалась (t=1,06, P>0,05).
Из приведенных данных следует, что подкормки на неспецифичном прокормителе оказывают значительное влияние на взаимоотношения монозоидных видов блох с чумным микробом. Поэтому различия в передаче возбудителя чумы самками и самцами, установленные в опытах на мышах, в природе могут проявляться по-другому. Тем не менее, они характеризуют определенные тенденции сезонной динамики трансмиссии чумного микроба.
Самки F. luculenta luculenta передали чумной микроб мышам в пяти (35,7 %), а самцы – в 11 (78,6 %) случаях из 14 (табл. 3). Половые различия по этому показателю достоверны (t=2,3, P<0,05), самцы передавали возбудителя чумы вдвое чаще самок. Половые различия у X. cheopis в трансмиссии недостоверны, однако прослеживается тенденция к увеличению данного показателя в группе, сформированной из самцов (табл. 3).
Таким образом, половые различия в трансмиссии чумного микроба выявлены у всех исследованных видов блох и они довольно значительны. Особенности штаммов, используемых в опытах, оказывали влияние на количественные (больше или меньше доля насекомых с «блоком» и число передач ими чумного микроба животным), но не на качественные характеристики этих показателей (тенденция к большим значениям у самцов по сравнению с самками). Приведенные данные свидетельствуют, что взаимоотношения чумного микроба с самцами блох обеспечивают в основном распространение возбудителя. Это связано с более высокой частотой кровососания самцов (Новокрещенова и др., 1968), их большей подвижностью (Кондрашкина, Дудникова, 1962) и склонностью к смене прокормителя (Rust, 1994).
Влияние абиотических факторов на алиментарную активность, смертность блох и их взаимоотношения с чумным микробом. Температура содержания зараженных насекомых является одним из мощных факторов, влияющих на частоту и скорость образования «блока» у любого вида блох (Козлов, 1979; Алексеев, Кондрашова, 1985; Ващенок, 1988; Hinnebusch et al., 1998). Наиболее интенсивен процесс блокообразования при температурах, благоприятных для жизни конкретного вида этих насекомых (Бибикова, Классовский, 1974). Представляет интерес исследование взаимоотношений чумного микроба и блох в зависимости от температуры, наблюдаемой в период активного проявления эпизоотического процесса в природном очаге.
Исследована векторная способность С. tesquorum altaicus при различной температуре с учетом активности кровососания и смертности насекомых. Имаго, инфицированных осенью и перезимовавших в условиях искусственной норы, содержали весной и летом следующего года при 2-10 С (с постепенным повышением температуры от весны к лету). Температура содержания оказывала существенное влияние на алиментарную активность «перезимовавших» насекомых (F=5,01, P<0,05). При низкой температуре (2-3 С) активность кровососания у самцов и самок не различалась, а при 9-10 С пивших самцов было больше: средняя за подкормку доля таких самцов составила 100±0,0 %, самок – 99,2±0,36 %. Необходимо уточнить, что наблюдения проведены над выжившими после зимовки особями, среди которых преобладали самки с остаточной зараженностью 50-64,3 % (при исходной 80-100 %).
С «молодыми» блохами опыты поставлены при более широком диапазоне температуры (от 5 до 27 С). По результатам однофакторного дисперсионного анализа (без учета пола имаго) температура содержания оказывала значительное влияние на активность кровососания «молодых» блох (F=4,50, P<0,01). Менее активно они питались при 7-8 С и 24-27 С. Сравнение данного показателя при трех промежуточных значениях температуры (7-8, 9-10 и 16-18 С), показало, что наиболее предпочтительна как для самок, так и самцов температура 9-10 С. При 7-8 С алиментарная активность выше у самок (94,9±0,90), чем у самцов (90,0±2,20), а при 16-18 С различий между полами не выявлено. При дальнейшем повышении температуры этот показатель снижался как у самок, так и у самцов. Так, при 22 С среди самок отмечено 98,9±0,83 % пивших особей, среди самцов – 100±0,0 %. При 24-27 С доля пивших самок составила 93,2±1,77 %, а самцов – 97,3±1,27 %.
Повышение температуры от 7-8 до 16-18 С влияло на смертность блох (F=4,56, P<0,01). Доля погибших за подкормку особей увеличивалась от 1,4±0,94 % до 8,3±2,03 %. При температуре 22-27 С относительное количество погибших в среднем за подкормку самцов (3,2±0,67 %) было выше, чем самок (1,2±0,46 %), почти в три раза.
Сравнительный анализ алиментарной активности «перезимовавших» и «молодых» блох при температуре 9-10 С показал, что она выше у переживших зиму особей (99,2±0,81 %), по сравнению с «молодыми» (92,6±1,14 %). Различия в смертности между этими группами насекомых незначительны (5,1±1,10 % для «молодых» блох, 4,7±1,72 % для «перезимовавших»).
Проанализировано влияние температуры на блокообразование и векторную способность насекомых. В первой серии экспериментов «перезимовавших» имаго содержали при температуре от 2 до 10 С, максимальное относительное количество особей с «блоком» (4,8 %) выявлено при 9-10 С. Формирование «блоков» отмечено при первой подкормке блох на сусликах, но в отдаленные после их заражающего кормления сроки (на 230-278 сутки).
Во второй серии экспериментов имаго выплода текущего года содержали при температуре от 5 до 27 С. Наибольшая доля «блокированных» особей (15,7 %) отмечена при 22-27 С, а формирование первых «блоков» происходило на 3-6 сутки после заражения блох.
У «перезимовавших» имаго существенных различий в частоте «блокообразования» при 2-10 С не выявлено, но отмечена тенденция к увеличению данного показателя с повышением температуры. При этом число передач чумного микроба зверькам достоверно возрастало (t=2,06, P<0,05).
У имаго выплода текущего года частота блокообразования достоверно увеличивалась с повышением температуры (t=2,69, P<0,01). Число передач при повышении температуры от 5-6 до 16-18 С несколько возросло, но различия недостоверны. При этом доля зверьков с генерализованной формой инфекции значительно увеличилась (t=2,25, P<0,05).
Необходимо отметить, что при максимальной температуре содержания насекомых (22-27 С) у них наблюдали высокий уровень блокообразования (15,7 %). Однако доля заболевших сусликов и среди них с генерализацией инфекции была ниже (10,5±7,03 %), чем в опытах с менее высокой температурой, но проведенных в пик эпизоотии в очаге, в июле-августе (46,2±13,83).
Таким образом, оптимальной для блокообразования и эффективной передачи С. tesquorum altaicus чумного микроба является температура от 9-10 до 16-18 С; более высокие или низкие значения приводят к снижению активности переносчика. Полученные экспериментальные данные позволяют прогнозировать характер изменения векторной способности блохи в исследованном диапазоне температур на энзоотичной территории Тувинского очага.
Влияние температуры и влажности на элиминацию и питание блох изучено достаточно хорошо (Брюханова, 1973; Бибикова, Классовский, 1974). В то же время даже на благоприятном гигротермическом фоне наблюдаются значительные суточные колебания гибели и активности кровососания имаго. Для установления возможных причин этого явления предпринято сравнительное изучение величины колебаний атмосферного давления, алиментарной активности и частоты гибели блох в разные сезоны (весна-лето-осень).
По результатам опытов доля пивших за подкормку самок и самцов C. tesquorum altaicus была ниже в периоды наибольших перепадов атмосферного давления (март-апрель, конец сентября-ноябрь). Летом исследуемый фактор менее влиял на алиментарную активность блох. Осенью алиментарная активность самцов зависела от величины атмосферного давления (F=4,17, P<0,05). Так, при давлении <716 мм рт. ст. в среднем за подкормку отмечали 68,7 % пивших самцов, при 716-725 – 87,1 %, а при >725 – 93,1 %.
Доля погибших особей значительно изменялась как среди самок (от 0 до 22,4 %), так и самцов (от 0 до 26,2 %). Чем сильнее амплитуда колебаний атмосферного давления, тем выше смертность насекомых в опытах. Коэффициент корреляции показателей составил 0,348 для самок (P<0,01) и 0,399 для самцов (P<0,01). Максимальные значения смертности (22,4 и 26,2 %) отмечены в марте-апреле, когда колебания давления достигали 32 мм рт. ст.
Следовательно, в природных условиях изменения атмосферного давления в зависимости от сезона года также могут оказывать влияние на алиментарную активность и смертность зараженных чумным микробом блох, что в конечном итоге может отражаться на их инфекционном потенциале. Вероятно, менее активные эпизоотические проявления в Тувинском природном очаге весной и осенью (по сравнению с летом) можно объяснить низкой алиментарной активностью и высокой частотой гибели блох в эти периоды.
ГЛАВА 4. ВТОРОСТЕПЕННЫЕ СПОСОБЫ ПЕРЕДАЧИ
ЧУМНОГО МИКРОБА БЛОХАМИ
Оценка различных способов передачи чумного микроба блохами дана в обобщающих публикациях И.Г. Иоффа (1941) и В.А. Бибиковой, Л.Н. Классовского (1974). Со времени опубликования последней работы прошло более 30 лет. За этот период накопился новый фактический материал, дающий возможность уточнить и расширить существующие представления о значимости второстепенных способов передачи чумного микроба в энзоотии этой инфекции. Показано, что лабораторные и дикие грызуны могут заражаться чумой при трансмиссивном, аэрогенном, контактном (через поврежденные покровы) и оральном введении возбудителя, причем эффективность перечисленных способов весьма высока (Кокушкин, 1995). Эти способы передачи могут осуществляться с непосредственным или опосредованным участием зараженных блох: укусами насекомых, при поедании зверьком инфицированных имаго, внесением возбудителя через поврежденную кожу грызуна из раздавленных эктопаразитов, или их экскрементов. С учетом полученных в экспериментах с C. tesquorum altaicus из Тувинского очага данных оценена роль второстепенных способов передачи чумного микроба блохами в циркуляции возбудителя (Базанова и др., 1991, 1997; Базанова, Маевский, 1999). Регистрировали частоту передачи блохами чумного микроба длиннохвостым сусликам и белым мышам при разных способах заражения грызунов.
При подкормках больших групп «неблокированных» блох (от 100 особей) отмечена более частая передача микроба сусликам «молодыми» имаго (36,4 %), чем «перезимовавшими» (9,1 %). Передача микроба небольшими группами (до 10 особей) «молодых» блох зависела от вида прокормителя. Так, гибели сусликов при кормлении на них по 10 блох как «перезимовавших», так и «молодых», не отмечено, а доля зверьков с иммуноаллергической перестройкой организма, установленной с помощью ППН, не различалась (57,9 % для «перезимовавших» блох, 55,0 % для «молодых). Однако при подкормках «молодых» особей на белых мышах гибель грызунов была высокой (31,7 %), достоверно превышая таковую среди сусликов (t=2,89, P<0,001).
Частота генерализации инфекции у сусликов и мышей при питании на них «молодых» имаго, несмотря на разные по количеству особей группы, не различалась (t=1,28, P>0,05). В некоторых случаях на сусликах питалось блох в 10-15 раз больше, чем на мышах. Возможно, это связано с тем, что использованный в опытах штамм Y. pestis (И-3226) высоко вирулентен для белых мышей (ЛД50 1 м.к.), для суслика этот показатель составлял 100 м.к.
Очевидно, что «неблокированные» блохи играют существенную роль в активизации эпизоотического процесса в природном очаге чумы. Возможность трансмиссии чумного микроба насекомыми зависит от количества особей, питавшихся на зверьке, интенсивности их заражения, физиологического состояния носителя и переносчика, сезона года (Базанова и др., 2003).
Установлена чувствительность длиннохвостого суслика к заражению возбудителем чумы алиментарным путем. Доля заболевших чумой зверьков в опытах достигала 26,7-40,0 %. Нужно отметить, что использование для инфицирования грызунов блох с различной степенью зараженности не повлияло на частоту заболевания зверьков и генерализации у них инфекции.
Заражение носителя чумой через поврежденную кожу возможно из раздавленных на теле животного насекомых или их выделений. Количество вносимых при этом микробов может резко возрастать при расчесах, которые провоцирует введенный блохами в ранку укуса секрет слюнных желез, обладающий аллергической активностью (Ващенок, 1988). В наших опытах передачу чумного микроба сусликам через поврежденную кожу из раздавленных насекомых и их выделений регистрировали в 65-70 % случаев. При этом у заболевших грызунов, зараженных таким способом, отмечена как затяжная форма инфекции с неинтенсивной бактериемией, так и генерализованная, с агональной септицемией (Базанова и др., 1992), что может служить существенным фактором дальнейшей трансмиссии чумного микроба.
Наиболее вероятным субстратом для сохранения чумного микроба в почве нор являются экскременты инфицированных блох и их трупы (Величко и др, 1980; Величко, 1981; Величко, Кондрашкина, 1984). Кроме того, зараженные насекомые во время дефекации выделяют во внешнюю среду мелкие бактериальные «глыбки», обсеменяя ее живыми, защищенными от неблагоприятных воздействий бактериями и заражая как шерсть зверька – хозяина, так и его убежище – гнездо (Коннов и др., 1984).
Установлена в эксперименте возможность заражения сусликов при контакте с субстратом гнезда, инфицированным экскрементами и трупами чумных блох. Чумной микроб сохранялся в субстрате до 427 суток, глубоко изменяя при этом свою морфологию (Базанова и др., 1997).
Следовательно, субстрат гнезда, инфицированный в летне-осенний период, может стать причиной заражения сусликов чумой в следующий эпизоотический сезон, особенно во время массового расселения молодых зверьков при посещении ими нежилых нор и активной роющей деятельности.
Сравнительная эффективность различных способов передачи чумного микроба блохами. Анализ данных показал, что более часто передача микроба происходила при заражении зверьков через поврежденную кожу, затем укусами «блокированных» и «неблокированных» насекомых (рис. 5).
 | Рис. 5. Эффективность различных способов передачи чумного микроба C. tesquorum altaicus длиннохвостому суслику: 1) укусами блох с «блоком»; 2) укусами «неблокированных» блох; 3) алиментарным путем; 4) через контакт с кожей зверька; 5) через субстрат гнезда. |
Характерно, что передача чумного микроба при групповых подкормках эктопаразитов осуществлялась в равной степени с участием «блокированных» особей и без их участия во все сезоны проведения опытов: весна – лето – осень. Однако генерализацию инфекции у заболевших чумой сусликов чаще определяло наличие в группе «блокированных» имаго. Очевидно, что блохи с «блоком» обеспечивают большую возможность возбудителю чумы для инвазии в организм интактного животного. Установлены различия в способности к передаче между «молодыми» и «перезимовавшими» насекомыми. Весной и летом «молодые» блохи передавали чумной микроб более активно, чем «перезимовавшие», даже при наличии среди последних особей с «блоком». Вероятно, это связано с изменчивостью микроба, находившегося длительное время в организме голодающих блох при низкой температуре.
Поскольку агональная бактериемия у теплокровных является одним из основных факторов, обеспечивающих возможность массового инфицирования блох, проведен сравнительный анализ частоты генерализации инфекции у грызунов при различных способах заражения. Генерализацию инфекции у сусликов достоверно чаще (t=3,78, P<0,001) регистрировали при питании на них «блокированных» особей (85,7 %), чем «неблокированных» (36,2 %). Однако частота генерализации инфекции у зверьков, зараженных через укусы «блокированных» имаго (85,7 %), алиментарным способом (78,6 %), а также через поврежденную кожу (66,7 %) не имела достоверных различий. В то же время укусы блох без видимого «блока» вызывали генерализацию инфекции у зверьков реже, чем алиментарный способ заражения (t=2,80, P<0,01) и через поврежденную кожу (t=2,53, P<0,05), а алиментарный уступал инфицированию через кожу (t=2,53, P<0,05).
Представляет интерес, что передача микроба укусами «блокированных» насекомых, алиментарным путем и внесением через поврежденную кожу зверька в эксперименте одинаково часто заканчивалась генерализацией инфекции у сусликов. Отсюда следует, что второстепенные способы передачи играют существенную роль в активизации эпизоотического процесса в природном очаге чумы, по отдельным показателям (например, генерализации инфекции) не уступая специфическому (укусам «блокированных» блох).
Высказано мнение (Солдаткин, Руденчик, 1988), что механизм передачи чумы, базирующийся на феномене бактериального «блока» преджелудка блох, по своим характеристикам не может обеспечивать эффективную трансмиссию микроба в природных очагах. Как показали результаты наших экспериментальных исследований, оценивая эпизоотологическую роль переносчиков, следует учитывать возможность осуществления ими всех известных способов передачи. Реализация этих способов передачи определяется исторически сложившейся системой взаимоотношений теплокровных животных, блох и чумного микроба, специфичной для каждого природного очага чумы.
ГЛАВА 5. МЕЖВИДОВЫЕ И МЕЖПОПУЛЯЦИОННЫЕ РАЗЛИЧИЯ
ВО ВЗАИМООТНОШЕНИЯХ БЛОХ С ЧУМНЫМ МИКРОБОМ
Важным моментом трансмиссии является взаимосвязь между способностью переносчиков к передаче чумного микроба и их экологическими характеристиками (Ващенок, 1999; Krasnov et al., 2006). Видовые различия и межпопуляционная дифференциация блох позволяют предположить их неоднородность в плане взаимоотношений с микробом, что может оказывать влияние на характер течения и другие параметры эпизоотического процесса.
Векторная характеристика массовых видов блох длиннохвостого суслика в Тувинском природном очаге чумы. Большинство экспериментальных исследований в очаге посвящено изучению взаимоотношений чумного микроба с C. tesquorum altaicus (Воронова, Крюков, 1975; Крюков, 1984; Воронова, 1978; Базанова и др., 1991). Эпизоотологическая роль других массовых видов блох суслика изучена слабо. Однако, по данным отчетов Тувинской ПЧС за 1964-2008 гг., на их долю также приходится большое количество штаммов чумного микроба, изолированных от блох. Так от C. tesquorum altaicus получено 722 культуры, R. li transbaikalica – 109, Oropsylla alaskensis – 69, F. elatoides elatoides – 52, N. mana – 21.
Проведен сравнительный анализ межвидовых различий C. tesquorum altaicus, N. mana, R. li transbaikalica, O. alaskensis, F. elatoides elatoides из природных популяций во взаимоотношениях с чумным микробом основного подвида при подкормках на сусликах. По результатам дисперсионного анализа фактор «вид блохи» оказывал влияние на алиментарную активность (F=7,23, P<0,001) блох и частоту формирования у них «глыбок» чумного микроба (F=4,96, P<0,01). Более активно питались на зверьке и погибали значительно реже других C. tesquorum altaicus. В организме C. tesquorum altaicus и O. alaskensis чаще, чем у двух других видов, формировались бактериальные «глыбки». Блохи R. li transbaikalica уступали по всем исследованным показателям C. tesquorum altaicus и O. alaskensis, а по алиментарной активности и N. mana. Частота гибели наиболее высока среди O. alaskensis и F. elatoides elatoides. Результаты опытов свидетельствуют, что основная эпизоотологическая роль C. tesquorum altaicus в очаге обусловлена биологическими особенностями вида. Так, блохи, у которых не менее активно формировались «глыбки» (O. alaskensis, N. mana, F. elatoides elatoides), уступали C. tesquorum altaicus либо в алиментарной активности (N. mana), либо характеризовались более высокой смертностью (F. elatoides elatoides).
Оценено влияние прокормителя на частоту возникновения «блока» преджелудка у блох суслика. Данный анализ проведен без учета пола насекомых, сезона проведения опытов и использованного штамма чумного микроба. Частота блокообразования у C. tesquorum altaicus при питании на длиннохвостом суслике (6,5 %) и белой мыши (6,7 %) достоверно не различалась (t=0,29, P>0,05). У F. elatoides elatoides и O. alaskensis формирование «блока» отмечено только при кровососании на суслике. У R. li transbaikalica этот показатель был выше при питании на сусликах (3,6 %) и пищухах (4,3 %), чем на мышах (1,8 %). Таким образом, для всех исследованных видов, кроме C. tesquorum altaicus, отмечена тенденция к снижению частоты блокообразования при кровососании на неспецифичных хозяевах.
Результаты проведенных опытов свидетельствуют, что роль прокормителя в процессе блокообразования у блох нельзя оценивать без учета влияния на этот фактор пола насекомого и сезона года. Так, весной при подкормках C. tesquorum altaicus на суслике выявляли «блоки» у особей обоих полов достоверно чаще (t=3,25, P<0,01), по сравнению с блохами, кормившимися на мышах, а летом эта тенденция выявлена только у самцов (t=3,44, P<0,001).
Характер проявления инфекционного процесса зависел от сроков проведения опыта, что отмечено при подкормках как групп блох, так и единичных особей с «блоком». Зверьки, на которых насекомых кормили в первой половине августа, погибали от генерализованной формы инфекции, во второй половине месяца у них регистрировали только затяжную форму заболевания.
В результате исследования для отдельных видов впервые получены факты блокообразования (O. alaskensis) и передачи чумного микроба (R. li transbaikalica, N. mana). Установлено существенное влияние на эти процессы половых различий насекомых и сезона проведения опытов, что необходимо принимать во внимание при оценке эпизоотологической роли вида блохи.
Межпопуляционные различия блох из Тувинского природного очага в трансмиссии чумного микроба. Среди сибирских очагов чумы наиболее изучена пространственная структура Тувинского, в пределах которого выделено 6 автономных участков очаговости (мезоочагов): Кара-Бельдырский, Монгун-Тайгинский, Толайлыгский, Барлыкский, Саглинский и Боро-Шайский (Вержуцкий, 1999). Блох C. tesquorum altaicus и R. li transbaikalica, добытых на территории Монгун-Тайгинского и Барлыкского участков, заражали штаммом И-3226 с Боро-Шайского участка. По биологическим свойствам, в том числе питательным потребностям, варианты микроба из Барлыкского и Боро-Шайского участков идентичны (Логачев, 1999).
Различий между C. tesquorum altaicus и R. li transbaikalica двух популяций в алиментарной активности, смертности и частоте образования «блока» не установлено. Однако межпопуляционные различия выявлены в частоте формирования в организме имаго «глыбок» микроба (F=9,02, P<0,01), которая была выше у насекомых обоих видов Барлыкской популяции (рис. 6).
 |  |
Рис. 6. Формирование бактериальных «глыбок» в организме блох из разных популяций:
а) – C. tesquorum altaicus, б) R. li transbaikalica. По оси абсцисс – подкормки, по оси ординат – доля блох с «глыбками (%).
Таким образом, более интенсивно исследуемый штамм возбудителя чумы формировал «глыбки» в организме блох, исторически контактировавших со сходными вариантами микроба.
Серия опытов выполнена с инсектарной культурой C. tesquorum altaicus, происходящей от насекомых Каргинской популяции (Монгун-Тайгинский мезоочаг). Инфицировали блох штаммами чумного микроба И-3327 и И-3428, изолированными в Саглинском и Монгун-Тайгинском мезоочагах соответственно. Оба штамма типичны для Тувинского очага, но отличаются по питательным потребностям в аминокислотах (Логачев, 1999). Подкормки насекомых проводили на белых мышах с той же периодичностью, что и на сусликах. Активность кровососания и частота гибели инфицированных разными штаммами насекомых не различалась. Достоверные различия между штаммами установлены в частоте формирования «блоков». По этому показателю штамм И-3428, происходящий с той же территории, что и исследуемая популяция блох, значительно превосходил штамм И-3327 (t=3,88, P<0,05).
Показано, что активность проявления эпизоотий на различных участках очаговости неравнозначна и зависит от ряда факторов, в том числе и от популяционных характеристик видов блох – основных переносчиков чумы, населяющих данную территорию (Сержанов и др., 1979; Князева и др., 1989; Корзун и др., 1998, 1999). Полученные нами результаты исследования предполагают специфичность исторически сложившихся взаимоотношений переносчика и возбудителя чумы, обитающих на участках Тувинского природного очага со сходными биоценотическими характеристиками.
Особенности взаимоотношений чумного микроба и блох из географически разобщенных популяций (из разных природных очагов). Осеннюю активизацию эпизоотического процесса в Горно-Алтайском очаге связывают с возрастанием численности монгольской пищухи и паразитированием на ней эффективного и массового в этот сезон переносчика – Paradoxopsyllus scorodumovi (Якуба и др., 1978; Иннокентьева и др., 2004). Эпизоотологическое значение этого вида блох в Тувинском очаге существенно ниже (Климов, Вержуцкий, 1984). Исследовали взаимоотношения массовых видов блох монгольской пищухи из Тувинского очага с чумным микробом основного и алтайского подвидов (из Тувинского и Горно-Алтайского очагов).
Имаго P. scorodumovi, P. dashidorzhii, Paramonopsyllus scalonae, F. hetera из природных популяций заражали штаммами чумного микроба основного (И-2638) и алтайского (И-3160) подвидов. Подкармливали насекомых на пищухах. Достоверных различий между использованными в опытах штаммами в частоте блокообразования не выявлено (Базанова, Климов, 1990).
При этом сравнительный анализ результатов исследования показал, во-первых, что доля погибших особей была выше среди блох, инфицированных возбудителем алтайского подвида (И-3160). Такая тенденция отмечена для всех четырех исследованных видов, а для F. hetera различия достоверны (t=6,02, P<0,001). Возможно, это связано с отсутствием адаптации блох из Тувы к микробу из Горно-Алтайского очага. Во-вторых, прослеживалась тенденция к более активному образованию «блоков» у насекомых из Тувы микробом алтайского подвида. В-третьих, P. scorodumovi и P. scalonae из Тувы, «блокированные» возбудителем чумы алтайского подвида чаще осуществляли передачу пищухам (t=2,29, P<0,05), чем особи с «блоками», сформированными чумным микробом основного подвида. Этот факт может являться дополнительным свидетельством адаптации микроба алтайского подвида к организму блох монгольской пищухи. Именно такие условия преимущественно обеспечивают его циркуляцию в Горно-Алтайском очаге чумы.
Как уже упоминалось, сибирские очаги являются северной окраиной Центрально-Азиатской зоны природной очаговости чумы, основная часть которой расположена в Монголии и Китае. В отдельных из них основными переносчиками являются одни и те же виды блох, как, например, C. tesquorum sungaris в Забайкалье (Вершинин, 1993) и Монголии (Цэрэнноров, 1999).
Проведено сравнительное исследование активности образования конгломератов возбудителя у C. tesquorum altaicus и C. tesquorum sungaris. Блох инфицировали чумным микробом, изолированным в Тувинском и Хэнтейском (Монголия) природных очагах (табл. 4).
Таблица 4
Активность формирования конгломератов микроба у C. tesquorum
(без учета пола насекомых)
| Подвид блохи | Средняя за подкормку доля блох (%) | |||
| Штамм И-3482 (Тува) | Штамм И-3230 (Монголия) | |||
| с конгломератами | с блоками | с конгломератами | с блоками | |
| C. tesquorum altaicus | 41,4 ± 0,50 | 1,9 ± 0,62 | 18,4 ± 3,03 | 0,6 ± 0,24 |
| C. tesquorum sungaris | 67,8 ± 5,61 | 5,4 ± 1,44 | 56,0 ± 8,36 | 6,2 ± 1,90 |
Среди C. tesquorum altaicus, происходящих из Тувинского очага, конгломераты чумного микроба формировались у большей доли особей за подкормку при инфицировании штаммом из того же очага (И-3482). Различия по этому показателю, рассчитанные в дисперсионном анализе, высоко достоверны (F=20,554, P<0,001), причем как у самок (F=9,436, P<0,01), так и у самцов (F=12,503, P<0,01). Средняя за подкормку доля «блокированных» разными штаммами самок и самцов не различалась.
Без учета пола у C. tesquorum sungaris из Забайкальского очага не выявлено различий между штаммами микроба из разных очагов как в формировании всех визуально учитываемых агрегированных форм, так и полных «блоков». Средняя за подкормку доля «блокированных» особей среди насекомых, инфицированных как тем, так и другим штаммом чумного микроба, практически была равной. Отмечено некоторое превышение доли таких блох при заражении возбудителем чумы из Монголии (И-3230) к концу опыта.
Более высокая частота образования «блоков» преджелудка у самок C. tesquorum altaicus отмечена при инфицировании чумным микробом из Тувинского очага (И-3482) (t=3,023, P<0,01). У самцов различий по этому показателю не выявлено. У самок C. tesquorum sungaris из Забайкалья «блок» достоверно чаще (t=2,314, P<0,05) возникал при инфицировании штаммом чумного микроба из Хэнтейского очага (И-3230), у самцов различий не установлено. Нужно отметить, что лабораторная популяция C. tesquorum sungaris происходит от имаго, добытых на участке территории Забайкальского природного очага, расположенном близко к границе с Монголией.
Очевидно, что взаимоотношения у переносчиков складывались более активно (по частоте выявления агрегированных форм) с теми вариантами чумного микроба, которые циркулируют или среди популяций основных хозяев блох, или на той же территории, откуда добыты насекомые, или на участках, непосредственно граничащих с этой территорией, что предполагает схожесть условий формирования паразитарных систем.
Особенности взаимоотношений чумного микроба и блох, исторически не контактировавших с возбудителем. Природный очаг чумы включает три обязательных компонента: популяцию чумного микроба, зверька – носителя инфекции и специфичного для данного хозяина вида блох как основного фактора передачи микроба. На неочаговой территории возможен тот же видовой состав зверьков и их эктопаразитов, но отсутствует возбудитель. Существует ли вероятность реализации эпизоотического процесса на неочаговой территории, например, при его заносе?
Исследованы инсектарные культуры C. tesquorum sungaris, начало которым положили насекомые, добытые на территории Усть-Ордынского Бурятского автономного округа и Тункинской долины (Республика Бурятия), то есть популяции блох, географически удаленные от природных очагов чумы. Контролем послужили блохи C. tesquorum altaicus, происходящие от особей из Тувинского природного очага. Установлены достоверные различия между популяциями C. tesquorum sungaris из разных географических мест в частоте образовании «блока» преджелудка при инфицировании штаммом И-3266 из Тувы (Базанова, Никитин, 2003). По совокупности результатов всех опытов у особей Усть-Ордынской популяции «блоки» формировались (6,7 %) достоверно чаще (t=2,330, P<0,05), чем у блох из Тунки (0,9 %). Частота блокообразования у C. tesquorum sungaris из Усть-Орды, инфицированных штаммом И-3266 (Тува), не зависела от сезона (весна, зима) проведения эксперимента (t=0,261, P>0,05), что может дополнительно свидетельствовать об отсутствии адаптации возбудителя к организму насекомых этой популяции. Тогда как у C. tesquorum altaicus из Тувы, инфицированных тем же штаммом чумного микроба, частота блокообразования зависела от сезона проведения опыта.
Наряду с C. tesquorum sungaris и C. tesquorum altaicus – основными переносчиками чумного микроба в сибирских природных очагах, опыты проведены с F. luculenta luculenta – второстепенным переносчиком. Предположена низкая эффективность этой блохи как переносчика чумы в Забайкалье (Феоктистов и др., 1974) и Монголии (Цэрэнноров,1999). Однако в период последней эпизоотии в Забайкальском очаге в 1970 г. культуры чумного микроба изолировали от блох F. luculenta luculenta, снятых с даурских сусликов, а также добытых из их гнезд и входов нор (Вершинин, 1993).
Особенности взаимоотношений с чумным микробом изучали у F. luculenta luculenta двух популяций, с учетом алиментарной активности и смертности блох. Исследуемые культуры насекомых получены от особей из Забайкальского очага (Забайкальская популяция) и неочаговой по чуме территории (Ольхонская популяция). Инфицировали блох типичными для Забайкальского природного очага штаммами чумного микроба. Формирование бактериальных «глыбок» наблюдали у блох обеих популяций.
Зарегистрирована эффективная трансмиссия возбудителя чумы блохами F. luculenta luculenta обеих популяций. Это произошло при первой подкормке имаго с исходной зараженностью 100 %, что подтверждено бактериологическим методом. В большинстве случаев факты передачи микроба животным установлены серологическим методом (в системе РПГА-РНАг). Частота передачи возбудителя чумы смешанными группами блох (самки и самцы) из Забайкалья равнялась 30-40 %. Самки Ольхонской популяции передали микроб 5 белым мышам из 14 (35,7 %), самцы – 11 из 14 (78,6 %), что, вероятно, обусловлено более высокой частотой формирования «глыбок» у самцов. Половые различия в передаче микроба блохами достоверны (t=2,3, P<0,05).
По результатам однофакторного дисперсионного анализа алиментарная активность выше у «ольхонских» особей (F=4,34, P<0,05), а частота гибели у «забайкальских» (F=6,47, P<0,05). У самцов обеих популяций эти показатели выше, чем у самок (F=21,68, P<0,001 и F=14,23, P<0,001).
Опыты показали, что F. luculenta luculenta как из Забайкальского природного очага, так и неочаговой территории могут осуществлять трансмиссию возбудителя чумы белым мышам (Bazanova et al., 2006). Возможна эффективная передача чумного микроба «неблокированными» блохами этого вида (с генерализацией инфекции у грызунов), которая, по мнению некоторых исследователей (Eisen et al., 2006, 2007; Wilder et al., 2008), способствует быстрому распространению возбудителя на ранней стадии заражения насекомых. Кроме того, у F. luculenta luculenta Ольхонской популяции активность кровососания выше, а смертность ниже, чем у имаго из Забайкалья, что может обусловить векторный потенциал блох с неочаговой территории.
Известно, что в одном природном очаге чумы могут обитать блохи – переносчики, относящиеся к разным популяциям, подвидам и видам (Лабунец, 1968; Голубинский и др., 1987; Князева, 1987; Князева и др., 1989). Факты возможной гибридизации отдельных подвидов (Лабунец, 1968; Золотова, Исханова, 1979; Якунин, Куницкая, 1992; Никитин, Нечаева, 1992; Ye R., 1997; Рую Е., 2000) побудили исследовать возможную роль этого процесса как мощного фактора физиологической перестройки организма (Захаров, 1987) в формировании общей эпизоотической картины в очаге.
Изучена способность чумного микроба к формированию «блоков» у гибридов двух подвидов C. tesquorum (табл. 5).
Таблица 5
Блокообразование у гибридов F1 и родительских подвидов С. tesquorum
| Вариант опыта | Штамм И-3266 | Штамм И-1996 | ||||
| заражен- ность блох (%) | число блох в группе | доля блох с блоком (%) | заражен-ность блох (%) | число блох в группе | доля блох с блоком (%) | |
| F1 | 100 | 114 | 6,1 ± 2,25 | 100 | 124 | 8,9 ± 2,56 |
| С. t. altaicus | 100 | 112 | 12,5 ± 3,12 | 80 | 110 | 8,2 ± 2,62 |
| С. t. sungaris | 100 | 83 | 7,2 ± 2,84 | 100 | 116 | 6,9 ± 2,35 |
Инфицировали блох штаммами Y. pestis из Забайкальского (И-1996) и Тувинского (И-3266) очагов. Выявлено, что у гибридных блох, также как и у родительских форм, формируется «блок» преджелудка (табл. 5), и они способны передавать возбудителя белым мышам. «Блокированные» гибриды заражали животных при кровососании небольших групп (1-3 особи).
Различия в трансмиссии чумного микроба гибридами F1 и родительскими подвидами блох при заражении разными штаммами обнаружены только в одном случае. Гибриды F1 передали чумной микроб из Забайкалья (И-1996) большему количеству мышей, чем С. tesquorum sungaris (t=2,25, P<0,05).
У особей F3 и F4 чаще формировался «блок» преджелудка, особенно у F3. Гибриды F3 и F4 передавали возбудителя белым мышам активнее, чем особи обоих родительских подвидов (рис. 8). В опытах с F3 различия в трансмиссии достоверны по сравнению и с С. tesquorum altaicus (t=2,52, P<0,05), и С. tesquorum sungaris (t=2,21, P<0,05). Гибриды F4 также чаще осуществляли передачу чумного микроба, чем С. tesquorum altaicus (t=3,35, P<0,01) и С. tesquorum sungaris (t=2,86, P<0,01).
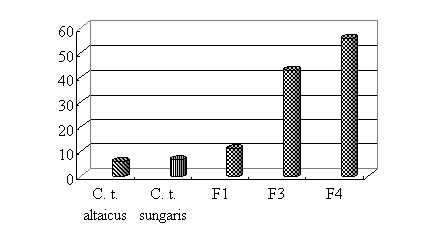 | Рис. 8. Трансмиссия чумного микроба родительскими подвидами и гибридами C. tesquorum. По оси абсцисс – группы блох, по оси ординат – количество заболевших чумой мышей (%). |
Частота блокообразования у гибридов F3 (в отдельных опытах 33 %) значительно превышала таковую у родительских подвидов (t=6,68, P<0,001 и t=6,93, P <0,001) и превосходила все известные (Воронова, Феоктистов, 1979; Голубинский и др., 1987; Базанова, 1993) значения (8-12 %) для этих блох.
В целом по трем опытам, гибриды передали чумной микроб 19 из 36 мышей (52,8 %), С. tesquorum altaicus – 10 из 37 (27 %), С. tesquorum sungaris – 7 из 50 (14 %). Среди гибридных блох выявляли больше особей с конгломератами микроба по сравнению с родительскими подвидами, и они обладали в 4–6 раз большей заражающей способностью, чем С. tesquorum sungaris. Данный феномен позволяет предположить возможность активизации эпизоотического процесса в местах потенциально «пригодных» для гибридизации насекомых. Не исключено, что подобная активизация эпизоотий неоднократно имела место в историческом плане и могла являться одним из механизмов генезиса существующих природных очагов чумы.
ГЛАВА 6. МОРФОЛОГИЧЕСКАЯ ИЗМЕНЧИВОСТЬ У БЛОХ
КАК ИНДИКАТОР ХАРАКТЕРА ИХ ВЗАИМООТНОШЕНИЙ
С ЧУМНЫМ МИКРОБОМ
Морфологические особенности и различия уровня флуктуирующей асимметрии билатеральных морф у блох из разных популяций Тувинского природного очага чумы. Для нескольких таксономических групп животных показана связь уровня флуктуирующей асимметрии (ФА) в проявлении билатеральных признаков с устойчивостью к воздействию паразитов (Захаров, 1987; Корзун, Никитин, 1997; Токмакова, 1998; Корзун и др., 1999).
Проведено сравнительное изучение C. tesquorum altaicus из Каргинской и Барлыкской природных популяций Тувинского очага по морфологическим особенностям и уровню ФА. При 100 % исходной зараженности дисперсионный анализ выявил различия в частоте формирования конгломератов у имаго Каргинской и Барлыкской популяций (F=9,02, P<0,01). Исследование морфологических особенностей показало, что у самок из Каргы размеры головы больше, чем из Барлыка (t=3,97, P<0,001), у самцов различий не обнаружено. Вооруженность щетинками по всем пяти исследуемым признакам выше как у самок, так и у самцов Каргинской популяции.
Установлено, что средний по четырем признакам хетотаксии уровень ФА у особей Барлыкской популяции в два раза выше, чем Каргинской (рис. 9). При этом в организме блох Барлыкской популяции более интенсивно формировались конгломераты чумного микроба (Базанова и др., 2006).
 | Рис. 9. Интенсивность агрегирования чумного микроба у C. tesquorum altaicus из двух популяций, различающихся по уровню ФА признаков хетотаксии |
Сравнивали уровень ФА у выборок самок и самцов блох Каргинской популяции со сформировавшимся «блоком» преджелудка и при его отсутствии. Результаты показали, что в шести случаях из восьми дисперсия разности больше у «блокированных» имаго (самок), при этом в двух – на достоверном уровне (F=0,867, P<0,05). Среди самцов больший уровень ФА по признаку «число щетинок на наружной поверхности голени» (F=0,800, P<0,05) также зарегистрирован у «блокированных» особей, по остальным четырем признакам статистически значимых различий не выявлено (Базанова и др., 2004).
Предполагается, что эпизоотологическая ситуация на участках очаговости чумы, наряду с комплексом других факторов, связана со структурой популяций переносчиков (Корзун, Никитин, 1997). Так как уровень ФА ниже у блох Каргинской популяции, чем Барлыкской, их разная векторная способность зависит либо от различий в восприимчивости к исследованному варианту микроба, либо от различного уровня ФА.
Уровень флуктуирующей асимметрии и блокообразование у блох с неочаговой по чуме территории. Проведено изучение уровня ФА с исследованием выборок имаго C. tesquorum sungaris, происходящих из Усть-Ордынской и Тункинской природных популяций (рис. 10).
 | Рис. 10. Доля «блокированных» блох в популяциях C. tesquorum sungaris, различающихся по относительному количеству асимметричных особей |
Анализ данных в совокупности по всем опытам позволил установить, что блохи из первой популяции «блокируются» достоверно чаще (t=2,33, P<0,05). Морфологический анализ выявил большую долю асимметричных особей среди насекомых из Усть-Орды. В среднем для четырех признаков уровень асимметрии у них составлял 55,3 %, а у имаго из Тунки (для этих же признаков) равнялся 40,7 % (рис. 10). Таким образом, у C. tesquorum sungaris из группировки с большей долей асимметричных особей отмечен и более высокий уровень блокообразования.
Уровень флуктуирующей асимметрии и блокообразование у гибридов блох. По сумме трех опытов у гибридов уровень ФА в среднем выше, чем у исходных форм (t=2,074, P<0,001), и у них чаще образуются «блоки» преджелудка, чем у родительских подвидов (табл. 6).
Таблица 6
Уровень флуктуирующей асимметрии (d2) у блох без «блока» и с «блоком»
в экспериментах с гибридными блохами
| № опыта | Вариант | Число особей | Признак 1 | Признак 2 | |||
| без блока | с блоком | без блока | с блоком | без блока | с блоком | ||
| 1 | C. t. altaicus | 26 | 3 | 0,234 | – | 1,994 | – |
| C. t. sungaris | 10 | 4 | 0,400 | – | 0,544 | – | |
| Гибриды F1 | 38 | 11 | 0,425 | 1,400** | 0,945 | 0,818 | |
| 2 | C. t. altaicus | 33 | 6 | 0,468 | 1,367* | 0,684 | 0,967 |
| C. t. sungaris | 71 | 8 | 0,514 | 0,571 | 1,140 | 1,923 | |
| Гибриды F3 | 42 | 23 | 0,527 | 0,846 | 0,967 | 2,439** | |
| 3 | C. t. altaicus | 60 | 2 | 0,451 | – | 1,080 | – |
| C. t. sungaris | 56 | 5 | 0,418 | 0,700 | 0,831 | 1,200 | |
| Гибриды F4 | 83 | 20 | 0,499 | 0,590 | 1,050 | 2,197* | |
| Всего | C. t. altaicus | 119 | 11 | 0,404 | 1,091** | 0,915 | 1,018 |
| C. t. sungaris | 137 | 17 | 0,471 | 0,485 | 0,890 | 1,279 | |
| Гибриды | 163 | 54 | 0,491 | 0,832** | 1,006 | 2,074*** | |
Результаты исследования уровня ФА и частоты блокообразования у гибридов аналогичны характеру различий между блохами из Усть-Орды и Тунки, а также полученными ранее данными о связи флуктуирующей асимметрии блохи Amphalius runatus и эпизоотической активности участков Горно-Алтайского очага чумы (Токмакова, 1998; Корзун и др., 1999, 2004).
Поскольку уровень ФА является информативным показателем, отражающим устойчивость индивидуального развития организмов к неоптимальным условиям внешней среды (Захаров, 1987), его высокие значения могут свидетельствовать об отклонении в развитии, которые обусловливают большую восприимчивость особей к заселению паразитами. Этим можно объяснить высокую частоту блокообразования у гибридов блох и насекомых с неочаговых по чуме территорий (у имаго «Усть-Ордынской» популяции).
ГЛАВА 7. СОХРАНЕНИЕ ЧУМНОГО МИКРОБА БЛОХАМИ
В ЗИМНИЙ СЕЗОН
Выживаемость интактных и зараженных чумой насекомых в зимний период. Для сибирских природных очагов чумы немаловажное значение имеет способность блох к переживанию холодного периода года, что в конечном итоге определяет сохранение ими возбудителя данной инфекции в зимний сезон. В Тувинском очаге, по результатам зимних раскопок гнезд длиннохвостого суслика, чумной микроб переживает холодный период в организме C. tesquorum altaicus (Ткаченко и др., 1983; Крюков, 1984; Вержуцкий, 2003). В Забайкальском очаге имаго массовых видов блох даурского суслика C. tesquorum sungaris и F. luculenta luculenta способны к переживанию зимнего периода (Вершинин, 1993).
Исследовали способность к выживанию при низких температурах у незараженных C. tesquorum altaicus, C. tesquorum sungaris и F. luculenta luculenta лабораторных популяций. Использовано по 200 имаго каждого подвида, с которыми поставлено по 2 варианта опыта. Опыт продолжался 30 дней. За этот период температура в толще снега менялась от 0 до -10,5 С.
За весь опыт доля погибших C. tesquorum altaicus составила 19 %, C. tesquorum sungaris – 63 %, F. luculenta luculenta – 39 %. Таким образом, C. tesquorum altaicus лучше переживали низкие температуры, чем C. tesquorum sungaris (t=6,33; P<0,001) и F. luculenta luculenta (t=3,11; P<0,01).
Способность к переживанию неблагоприятных условий изучали у инфицированных чумным микробом C. tesquorum двух подвидов (табл. 7).
Таблица 7
Переживание низкой температуры зараженными чумным микробом блохами
| Подвид блох | Количество блох | Количество выживших блох | ||||||
| t между | ||||||||
| абс. | % | абс. | % | и | подвидами | |||
| C. tesquorum altaicus | 48 | 17 | 43 | 89,6 | 14 | 82,4 | 0,78 | 3,66* |
| C. tesquorum sungaris | 44 | 17 | 29 | 65,9 | 7 | 41,2 | 1,76 | |
Низкотемпературный период (18 дней) без подкормок лучше пережили C. tesquorum altaicus, чем C. tesquorum sungaris. Достоверных различий по этому показателю между самками и самцами за этот период не отмечено.
Более длительные сроки выживания установлены для инфицированных имаго C. tesquorum altaicus, зимовавших в искусственных нежилых «гнездах» суслика. До весны и лета следующего года выживало 69,9-87,6 % из зараженных осенью самок и 48,1-60,9 % самцов. Из блох, инфицированных летом, дожили до теплого периода следующего года 29,3 % особей, причем только самки (Базанова, Маевский, 1996). Таким образом, значительно успешнее доживали до нового эпизоотического сезона имаго, инфицированные осенью (по сравнению с летом) предыдущего года (без учета пола t=13,35; P<0,001), причем как самки (t=11,49, P<0,001), так и самцы (t=8,40, P<0,001). Самки C. tesquorum altaicus могут переживать не один, а два зимних периода и сохранять чумной микроб в течение 21-22 месяцев. Укусы таких особей вызывали иммуноаллергическую перестройку в организме сусликов, что выявлялось положительным ППН через 642-646 суток после заражения блох.
Поскольку бактериальный «блок» преджелудка у блох рассматривается как обязательное условие специфического способа передачи чумного микроба от грызуна к грызуну (Бибикова, Классовский, 1974; Hinnebuch et al., 1998), большое значение может иметь продолжительность жизни таких блох. Установлено, что длительность жизни «блокированных» C. tesquorum altaicus при +8 С составляла от двух до 20 суток. Половых различий в частоте гибели насекомых за этот период не выявлено, но большинство самцов погибло на вторые сутки, а самок – на шестые. Высокая продолжительность жизни (до 219 суток) отмечена у «блокированных» C. tesquorum altaicus, содержащихся при более низких температурах (от +6 до -6 С) в условиях искусственного гнезда суслика. Такие блохи могут длительно сохранять чумной микроб, о чем свидетельствуют результаты бактериологического исследования оставшихся в живых к концу апреля самок с сохранившимся «блоком». Но передачи микроба зверькам при подкормке этих блох не произошло, видимо в связи с низкой чувствительностью суслика к возбудителю весной.
Зимующие в состоянии оцепенения N. mana также могут длительно сохранять чумной микроб (348 суток) и передавать его зверькам летом следующего после их заражения года (Базанова, Хабаров, 1993).
Состояние агрегированности чумного микроба в организме переносчика и его значение для сохранения микроорганизма. Фенотипические, в том числе морфологические, изменения чумного микроба возникают в организме имаго блох при резком изменении интенсивности обменных процессов: нарушении питания, холодовом оцепенении, повышении температуры (Бибикова, Классовский, 1968, 1974; Акиев и др., 1976; Воронова, Феоктистов, 1978; Бейер, 1979; Козлов, 1979; Hinnebusch et al., 1998).
Изучена морфологическая изменчивость и способность к агрегированию чумного микроба в организме C. tesquorum altaicus при неблагоприятных условиях существования переносчика (длительное голодание, холодный период года). Установленные факты свидетельствуют, что именно в агрегированном состоянии микроб находится в имаго зимний период. Результаты весеннего (апрель) опыта с «перезимовавшими» блохами, по доле особей с агрегированным микробом при первых подкормках наиболее близки данным, полученным к концу осеннего (сентябрь) эксперимента. Так, осенью за три последние подкормки выявлено 45,8±1,17 % особей с конгломератами микроба, весной за три первых подкормки «перезимовавших» насекомых – 49,1±15,39. Для сравнения отметим, что весной средняя доля «молодых» особей с конгломератами за три первые подкормки составила 0,7 % при их 100 % зараженности. У «молодых» насекомых этот показатель при трех первых подкормках существенно ниже (t=13,02, P<0,001), чем у «перезимовавших».
Доля «перезимовавших» имаго с агрегированным микробом за первые три подкормки была стабильно высокой как весной, так и летом. Весной отмечено 45,9±4,03 % особей с конгломератами, летом – 42,3±2,45 %. Зараженность «перезимовавших» блох составила 64,3 и 50 % соответственно.
Состояние агрегированности чумного микроба усиливается при голодании насекомых. На это указывают данные опыта с двумя группами «молодых» имаго в летний период. Подкормки одной группы начали на третьи сутки после инфицирования, второй – на 16. В первой группе появление особей с агрегатами микроба зарегистрировано на восьмые сутки (3 подкормка) после заражающего кормления имаго. Средняя доля таких блох по трем первым подкормкам составила 1,8 %, что достоверно меньше (t=5,63, P<0,001), чем в летних опытах с «перезимовавшими» имаго (в среднем 44,1±3,24).
Во второй группе после первой подкормки (16 сутки от заражения насекомых), выявлено 23 особи (12,2 %) с конгломератами возбудителя. При этом средняя доля таких имаго составила за три первые подкормки 9,7 %. Различия между группами насекомых, которых начали подкармливать на третьи и 16 сутки после заражения, достоверны (t=5,89, P<0,001). Длительное голодание привело к увеличению числа особей с агрегированным микробом при первом кормлении. В дальнейшем при регулярном кормлении блох межгрупповые различия, выявленные по трем первым подкормкам, нивелировались. По доле особей, с конгломератами микроба, между «молодыми», голодавшими после заражения в течение 16 суток, и «перезимовавшими» имаго, достоверных различий не установлено, что указывает на изменчивость микроба и повышенную агрегацию в любых стрессовых условиях.
К неоптимальным условиям существования чумного микроба в организме блохи относится высокая температура, лежащая вне температурного оптимума для имаго данного вида. В наших экспериментах «молодых» насекомых содержали в опытах весной при 22-27° С, летом – при 8-10° С. В первом случае особи с агрегированным микробом выявлены на третьи сутки после инфицирования насекомых (1 подкормка), во втором – на восьмые (3 подкормка). Очевидно, быстрый переход возбудителя в состояние агрегированности в организме блохи явился ответной реакцией на неоптимальную температуру окружающей среды для C. tesquorum altaicus и самого микроба.
Изучение морфологии чумного микроба у «перезимовавших» C. tesquorum altaicus с «глыбками» показало наличие палочковидных и шаровидных форм в соотношении 2:1. У «молодых» блох преобладали шаровидные формы и редко встречались палочки с невыраженной биполярностью, у «блокированных» («молодых» и «перезимовавших») – только шаровидные очень мелкие (0,2-0,5 мкм), бледно окрашенные формы. На ультратонких срезах желудочно-кишечного тракта «молодых» особей в первые сутки после заражения микроб представлен палочковидными и шаровидными формами, по мере голодания блох в основной массе отмечали округлившиеся бактериальные клетки, иногда увеличенные в размерах. В «перезимовавших» насекомых чумной микроб претерпевает более глубокие изменения, что, особенно, выражено в ультраструктуре самих микроорганизмов, которая включает дополнительные мембранные структуры (Маевский и др., 1994).
Таким образом, к изменению морфологии и агрегированию чумного микроба в блохах приводят особенности физиологического состояния насекомых при их подготовке к зимней диапаузе, длительном голодании и изменении температуры окружающей среды, что обеспечивает его сохранение.
Изучены особенности переживания чумным микробом холодного времени года в самках и самцах C. tesquorum altaicus. Блох инфицировали в сентябре. До зимовки одну группу подкармливали на сусликах периодически (1 вариант), другую содержали на хозяине постоянно (2 вариант). В 1 варианте опыта за последнюю подкормку (4 октября) конгломераты микроба отметили у 67,9 % самок и 48,2 % самцов (t=3,12; P<0,01). Средняя за опыт доля таких имаго (без учета пола) равнялась 18,9 %. В двух группах 2 варианта опыта перед помещением в «гнездо» суслика доля блох с агрегированным микробом составила 2,4 и 4,7 %, среди них преобладали самки. Представляет интерес факт, что во 2 варианте доля имаго с конгломератами была в 10-20 раз ниже, чем в 1, а различия в уровне блокообразования между ними не превышали 2-3 кратных. Вероятно, повышенное агрегирование микроба в организме блохи не обязательно приводит к увеличению уровня блокообразования, во всяком случае, осенью, при подготовке имаго к зимней диапаузе.
Имаго 1 варианта опыта изъяли из «гнезда» в апреле. Выжило 69,4 % самок и 46,9 % самцов. После первой подкормки конгломераты микроба отмечены у 50,0 % самок и 23,7 % самцов, различия достоверны (t=3,5; P<0,001).
Блохи 2 варианта опыта извлечены из «гнезда» в июне. В одной группе выжило 64,9 % самок и 40,0 % самцов, в другой – 52,6 % и 33,3 % соответственно. Выживаемость имаго в 1 и 2 вариантах опыта достоверно не различалась и была значимо выше у самок (t1=4,2, t2=4,1; P<0,001 для обеих групп 2 варианта). Летом после первой подкормки в одной группе доля самок с конгломератами микроба составила 42,5, самцов – 26,1 % (t=2,0; P<0,05), в другой – 52,6 и 33,3 % (t=2,5; P<0,05) соответственно.
Результаты проведенного анализа указывают на преобладающее значение самок C. tesquorum altaicus в сохранении чумного микроба и раскрывают механизмы его переживания в организме блох в холодное время года. Микроб в агрегированном состоянии накапливается осенью преимущественно в желудочно-кишечном тракте самок основного переносчика. В период зимовки насекомых продолжается процесс изменения морфологии микроорганизма и агрегирования отдельных клеток в конгломераты, в таком состоянии чумной микроб может сохраняться в организме блох длительное время (Маевский и др., 1994; Базанова и др., 2007).
Особенности скрытой (латентной) фазы эпизоотического процесса в Тувинском природном очаге чумы. Течение эпизоотического процесса в Тувинском очаге разделяют на две фазы – скрытую (латентную), протекающую в период спячки суслика, и активную – во время бодрствования животного (Равдоникас, 1985). Отмечено по результатам зимних раскопок, что в зимний период гнезда летнего типа остаются свободными от зверьков, где и сохраняется значительная часть имаго C. tesquorum altaicus и чумной микроб в их организме зимой (Крюков, 1984; Вержуцкий и др., 2003).
Как и многие природные процессы, эпизоотический недоступен для прямого наблюдения за его протеканием (Солдаткин, Фенюк, 1968). Нами предпринята попытка изучения в эксперименте отдельных элементов латентной фазы эпизоотического процесса для уточнения механизмов сохранения чумного микроба в зимний период.
Изучена восприимчивость и чувствительность суслика к чумному микробу перед зимней спячкой, а также возможность инфицирования зимующих совместно с хозяином блох. Установлены факты инфицирования сусликов чумой осенью через укусы блох. Весной передачи возбудителя зверькам, вышедшим из спячки, не отмечено (Маевский и др., 1999). Выживаемость эктопаразитов совместно с хозяином в среднем составила 38,5 %. Из 400 интактных блох, помещенных на сусликов осенью, весной собрано 154 живых и 20 мертвых. Чумной микроб от эктопаразитов выделен в одном случае. Результаты опыта указывают на низкую выживаемость C. tesquorum altaicus совместно с хозяином и слабую возможность заражения на суслике зимой.
В серии экспериментов продолжительностью более 600 суток изучена выживаемость зараженных блох, зимующих отдельно от хозяина. Доля выживших C. tesquorum altaicus, инфицированных осенью, составляла в весенне-летний период следующего года 63,5-77,3 %. Чумной микроб отмечен весной у 64,3 %, летом – у 50 % таких насекомых. Отдельные особи (самки) сохраняли микроб 21,5-22 месяца, при этом зарегистрирован факт формирования «блока» на 411 сутки после заражения блох (Базанова, Маевский, 1996).
В рассматриваемых опытах различия в выживаемости имаго, зимующих отдельно или совместно с прокормителем, высоко достоверны (t=5,129, P<0,001). При этом самки во всех опытах пережили зимний период лучше, чем самцы (для первого опыта t=3,421, для второго t=5,370, для третьего t=4,406; для всех опытов P<0,001). Полученные данные свидетельствуют, что C. tesquorum altaicus гораздо лучше переживают зимний период отдельно от хозяина. «Перезимовавшие» насекомые весной слабо осуществляли передачу чумного микроба суслику, но летом этот процесс активизировался.
Рассмотрим, как изменялась доля особей с чумным микробом в агрегированном состоянии осенью у блох выплода текущего года, а затем весной и в начале лета у перезимовавших отдельно от хозяина имаго.
Осенью доля «молодых» особей с конгломератами микроба (до 70 %) возросла к концу эксперимента (3 и 4 октября). По сравнению со средним за осенний опыт показателем (15,8 %) доля блох с агрегированным микробом к началу октября увеличилась более чем в четыре раза. В этот период имаго заканчивают физиологическую перестройку для переживания зимнего периода в состоянии оцепенения, что, видимо, приводит к изменению взаимодействия микроба с организмом переносчика, возрастанию его способности к агрегации и сохранению в таком состоянии в C. tesquorum altaicus зимой.
Весной среди «перезимовавших» насекомых при первых трех подкормках (на 228-233 сутки после заражающего кормления) отмечено 44,3-48,0 % особей с конгломератами микроба. Кроме того, сразу же после первой подкормки выявлено пять блох с «блоками». При дальнейших подкормках доля особей с «глыбками» постепенно уменьшалась. Весной доля блох с конгломератами в среднем составила 31,5 %, что вдвое выше, чем осенью. Необходимо указать, что передача микроба произошла только в одном случае из 12 при кормлении 83 блох без видимого «блока» на 20 сутки от начала подкормок (19 мая), без генерализации инфекционного процесса у суслика. Факт передачи установлен на основании положительных результатов исследования на наличие антител к чумному микробу (РПГА – 1:320 и РНАг – 1:2560).
Летом периодические подкормки «перезимовавших» насекомых проводили с 17 по 27 июня. После первой-третьей подкормок (на 275-280 сутки от инфицирования блох) зарегистрировано от 37,9 до 51,0 % особей с агрегированным микробом. «Блокированные» блохи (4 экз.) отмечены после первой подкормки, что указывает на преимущественное значение «перезимовавших» блох как вектора в июне, по сравнению с имаго выплода текущего года. Средняя доля насекомых с агрегированным микробом составила 23,5 %, несколько снизившись по сравнению с весной. Следует отметить, во-первых, что эффективная передача микроба сусликам в летний период произошла уже при трех первых подкормках «перезимовавших» насекомых. Во-вторых, у всех трех заболевших чумой зверьков отмечено наличие возбудителя в крови, что способствует дальнейшей трансмиссии микроба. В-третьих, летом выше соотношение числа блох с «блоком» к их количеству с конгломератами. Осенью при более высокой, чем летом, доле таких насекомых (до 70 %), увеличения числа особей с «блоком» не наблюдали.
Результаты рассмотренных опытов показывают, что в Тувинском природном очаге основным местом сохранения чумного микроба в холодное время года является организм имаго основного переносчика, которые переживают зимний период в нежилых норах длиннохвостого суслика. Экспериментально установлено, что для осеннего периода в очаге характерно накопление возбудителя в организме блох в агрегированном состоянии, которое проявляется весной в исходно высокой концентрации конгломератов бактерий у перезимовавших имаго. При этом чумной микроб глубоко изменяет свою морфологию и ультраструктуру, а также культуральные свойства.
У блох с агрегированным микробом образование чумного «блока» возможно уже в первые дни питания на сусликах. В летний период возрастает вероятность передачи инфекции грызунам во время первого кровососания на них перезимовавших в состоянии оцепенения эктопаразитов. Перезимовавшие и сохранившие в своем организме вирулентного возбудителя чумы насекомые могут обеспечить начало следующей активной фазы эпизоотического процесса в Тувинском природном очаге, являясь основным фактором, обеспечивающим трансмиссию микроба в мае-июне. В июне-июле инфекциозность укусов «перезимовавших» особей возрастает и не уступает таковой у имаго выплода текущего года (Базанова и др., 2004). Чумной микроб, сохранившись в конгломератах в зимовавших в состоянии оцепенения блохах, под влиянием внешних условий (в том числе повышения температуры воздуха) вновь восстанавливает способность к трансмиссии. Появившиеся летом суслики нового поколения активно посещают старые норы, входят в контакт с пережившими зиму имаго (чаще самками), способными к эффективному заражению животных возбудителем чумы. Вследствие этого возобновляется циркуляция чумного микроба в Тувинском природном очаге.
ВЫВОДЫ
1. Во взаимоотношениях чумного микроба с блохами наблюдается выраженная сезонная динамика, установленная по следующим показателям: частота и сроки блокообразования, инфекциозность укусов «блокированных» особей, частота передачи инфекции и генерализации инфекционного процесса у заболевших носителей. Максимальные значения этих показателей в экспериментах, проведенных на сусликах, совпадают по времени (середина лета) с пиком активизации эпизоотического процесса в природном очаге.
2. Возрастание значений исследованных показателей от весны к лету проявляется также при питании эктопаразитов на лабораторных животных, не имеющих сезонных колебаний чувствительности к возбудителю чумы. Этот феномен может свидетельствовать о том, что сезонность взаимодействия чумного микроба и блох обусловлена их коадаптацией, прочно вошла в годовой цикл жизнедеятельности каждого и имеет место независимо от питания насекомых на специфичном или неспецифичном хозяине.
3. Половые различия у блох оказывают существенное влияние на их взаимоотношения с чумным микробом. «Блок» преджелудка у самцов формировался в большинстве случаев чаще и в более короткие сроки после заражающего кормления. Отмечается большее число передач чумного микроба группами блох, сформированных из самцов. Самцы играют значительную роль в распространении возбудителя в период начала выплода нового поколения блох, когда в популяции преобладают особи этого пола. Длительное сохранение чумного микроба происходит в основном в организме самок, что обусловлено их большей устойчивостью к неблагоприятным условиям окружающей среды и продолжительностью жизни.
4. Установлено, что оптимальной для образования «блока» у С. tesquorum altaicus – основного переносчика чумы в Тувинском природном очаге – и передачи блохами чумного микроба является температура от 9-10 С до 16-18 С. Более высокие или низкие ее параметры снижают векторную способность блохи. Высокая амплитуда колебаний атмосферного давления повышает смертность инфицированных блох обоего пола что, в конечном итоге, отражается на инфекционном потенциале переносчика.
5. «Неблокированные» инфицированные блохи имеют равнозначное значение с «блокированными» в активизации эпизоотического процесса. При этом «неблокированные» особи, особенно самки, способны длительно (до двух лет) сохранять чумной микроб и осуществлять его передачу зверькам. Трансмиссия микроба насекомыми без видимого «блока» отмечена при кормлении на животных разных по численности групп эктопаразитов (от единичных особей до 150-200). Чувствительным к возбудителю чумы зверькам эффективная передача возможна при кровососании на них небольших (до 10 особей) групп «неблокированных» блох.
6. Существенную роль в активизации и распространении эпизоотического процесса в природном очаге чумы играют второстепенные способы передачи микроба блохами, такие как алиментарный и через поврежденную кожу. Передача чумного микроба «блокированными» насекомыми, алиментарным путем и внесением через поврежденную кожу зверька при расчесах одинаково часто заканчивается генерализацией инфекции у заболевших сусликов, что увеличивает вероятность заражения блох при кровососании на таких зверьках и обеспечивает дальнейшую трансмиссию возбудителя.
7. Эпизоотологическая роль C. tesquorum altaicus как основного переносчика в Тувинском природном очаге определяется биологическими особенностями вида (максимальной продолжительностью жизни, активностью кровососания, устойчивостью к неблагоприятным условиям окружающей среды, высокой способностью к переживанию низких температур). Блохи, у которых активно формировались «глыбки» микроба (N. mana и F. elatoides elatoides), уступали C. tesquorum altaicus либо в алиментарной активности (N. mana), либо имели более высокую смертность (F. elatoides elatoides).
8. Установлены межпопуляционные различия у блох из Тувинского природного очага по ряду морфологических показателей, уровню флуктуирующей асимметрии и особенностям взаимоотношений с чумным микробом. У C. tesquorum altaicus и R. li transbaikalica из двух популяций более активно формируются конгломераты при инфицировании вариантом микроба, аналогичным циркулирующему на том участке, откуда происходят насекомые, что может указывать на специфичность исторически складывающихся взаимоотношений блох и чумного микроба с одного участка очаговости.
9. Установлена возможность формирования устойчивых функциональных связей между географически разобщенными (из разных очагов) чумным микробом и насекомыми, а также микробом и исторически не контактировавшими с возбудителем блохами (гибридами и насекомыми с неочаговых по чуме территорий). Данный феномен позволяет предположить возможность укоренения чумного микроба на территориях, где ранее инфекция не присутствовала, и активизацию эпизоотического процесса в местах потенциально «пригодных» для гибридизации насекомых.
10. Состояние агрегированности чумного микроба в организме блохи обусловлено изменением его фенотипа и связано с биологическими особенностями насекомого. Такое состояние обеспечивает микробу возможность длительного пребывания в организме переносчика, защищая его от действия неблагоприятных факторов. Перезимовавшие в холодовом оцепенении имаго, сохранившие чумной микроб, обеспечивают активизацию эпизоотического процесса весной и ранним летом. В этот период такие блохи имеют важное эпизоотологическое значение, поскольку образование «блока» и передача микроба грызунам возможны уже при первом кровососании на них «перезимовавших» эктопаразитов.
11. Устойчивость сибирских природных очагов чумы основана на исторически сложившейся, специфичной для каждого очага системе взаимоотношений теплокровных животных, блох и чумного микроба. Значительное влияние на взаимоотношения чумного микроба и блох оказывают сезон года, популяционные особенности и пол насекомых, а также стабильность онтогенетического развития особей.
Основные работы, опубликованные по теме диссертации
Статьи в рецензируемых журналах, рекомендованных ВАК
1. Сезонная динамика блокообразования у блохи Citellophilus tesquorum altaicus из Тувинского природного очага чумы / Л.П. Базанова, И.Ф. Жовтый, М.П. Маевский и др. // Мед. паразитол. – 1991. – № 1. – С. 24 – 26.
2. Изменчивость Yersinia pestis в организме блохи / М.П. Маевский, Л.П. Базанова, Н.П. Конов и др. // Журн. микробиол. – 1994. – № 3. – С. 16-21.
3. Экспериментальное изучение гибридов от скрещивания блохи Citellophilus tesquorum двух подвидов передавать возбудителя чумы / А.Я. Никитин, Л.П. Базанова, Л.К. Нечаева и др. // Мед. паразитол. – 1995. – № 4. – С. 14-17.
4. Базанова Л.П. Длительность сохранения возбудителя чумы в организме блохи Citellophilus tesquorum altaicus / Л.П. Базанова, М.П. Маевский // Мед. паразитол. – 1996. – № 1. – С. 45-48.
5. Базанова Л.П. Экспериментальное изучение возможности сохранения возбудителя чумы в субстрате гнезда длиннохвостого суслика / Л.П. Базанова, М.П. Маевский, А.В. Хабаров // Мед. паразитол. – 1997. –№ 4. – С. 37 -39.
6. Заражающая способность блох Citellophilus tesquorum altaicus в различные месяцы эпизоотического сезона / Л.П. Базанова, А.Ф. Попков, А.В. Хабаров, А.Г. Вылков // Пробл. особо опасных инфекций. – Саратов. – 1999. – Вып. 79. – С. 202 – 203.
7. Маевский М.П. О сохранении возбудителя чумы у длиннохвостого суслика из Тувинского природного очага в зимний период / М.П. Маевский, Л.П. Базанова, А.Ф. Попков // Мед. паразитол. – 1999. – № 1. – С. 55-57.
8. Базанова Л.П. Различия в образовании блока преджелудка у самцов и самок Xenopsylla cheopis (Siphonaptera: Pulicidae) / Л.П. Базанова, Г.А. Воронова, Е.Г. Токмакова // Паразитология. – 2000. – Т. 34, вып. 1. – С. 56-59.
9. Базанова Л.П. О блокообразовании у блохи Citellophilus tesquorum altaicus Ioff, 1936 в зависимости от пола насекомого / Л.П. Базанова, А.В. Хабаров // Мед. паразитол. – 2000. – № 1. – С. 42-44.
10. К использованию полимеразной цепной реакции для детекции чумного микроба в блохах / С.В. Балахонов, Г.А.Воронова, М.Ю. Шестопалов, Л.П. Базанова, Е.Г. Токмакова // Мед. паразитол. – 2000. – № 3. – С. 29-31.
11. Совершенствование эпизоотологического мониторинга в природных очагах чумы Сибири / Т.И. Иннокентьева, А.Ф. Попков, Д.Б. Вержуцкий, Л.П. Базанова и др. // Пробл. особо опасных инфекций. – Саратов, 2002. – Вып.1 (83). – С. 52-58.
12. Базанова Л.П. Влияние температурного фактора на векторную способность блохи Citellophilus tesquorum altaicus / Л.П. Базанова, А.Я. Никитин, Н.Ф. Галацевич // Мед. паразитол. – 2003. – № 3. – С. 24 – 27.
13. Базанова Л.П. Значение блокированных и неблокированных блох Citellophilus tesquorum altaicus (Ioff, 1936) в передаче чумной инфекции / Л.П. Базанова, Е.Г. Токмакова, М.П. Маевский // Пробл. особо опасных инфекций. – Саратов, 2003. – Вып. 86. – С. 14-20.
14. Воронова Г.А. Значение блох (Siphonaptera) разных видов в поддержании эпизоотий чумы в сибирских природных очагах / Г.А. Воронова, Л.П. Базанова // Бюл. ВСНЦ СО РАМН. – Иркутск, 2004. – Т. 2, № 1. – С. 58-65.
15. Базанова Л.П. Влияние сезонных особенностей и половых различий на передачу чумного микроба блохами Citellophilus tesquorum altaicus (Ioff, 1936) и эпизоотический процесс / Л.П. Базанова, Г.А. Воронова, Е.Г. Токмакова // Бюл. ВСНЦ СО РАМН. – Иркутск, 2004. – Т. 2, № 1. – С. 28-34.
16. Сравнительная информативность полимеразной цепной реакции и традиционных методов детекции чумного микроба в блохах и лабораторных животных / С.В. Балахонов, Г.А. Воронова, В.В. Синьков, Л.П. Базанова и др. // Бюл. ВСНЦ СО РАМН. – Иркутск, 2004. – Т. 2, № 1. – С. 39-44.
17. Качественная неоднородность блохи Citellophilus tesquorum по способности к блокообразованию / В.М. Корзун, А.Я. Никитин, Л.П. Базанова и др. // Бюл. ВСНЦ СО РАМН. – Иркутск, 2004. – Т. 2, № 1. – С. 89-94.
18. Базанова Л.П. Сезонная динамика заражающей способности блохи Citellophilus tesquorum altaicus в Тувинском природном очаге чумы / Л.П. Базанова, А.Ф. Попков, Н.Ф. Галацевич // Паразитология. – 2004. – Т. 38, вып. 1. – С. 46-52.
19. Некоторые особенности взаимоотношений Citellophilus tesquorum altaicus (Siphonaptera: Ceratophyllidae) с возбудителем чумы / Л.П. Базанова, Г.А. Воронова, Е.Г.Токмакова, В.В. Синьков // Паразитология. – 2004. – Вып. 38, № 2. – С. 139-149.
20. Различия между двумя популяциями Citellophilus tesquorum altaicus из Тувинского природного очага чумы / Л.П. Базанова, Д.Б. Вержуцкий, А.Я. Никитин и др. // Мед. паразитол. – 2004. – № 1. – С. 37-39.
21. Базанова Л.П. Сезонные особенности динамики агрегированного состояния возбудителя чумы в организме Citellophilus tesquorum altaicus (Siphonaptera) / Л.П. Базанова, А.Я. Никитин, М.П. Маевский // Мед. паразитол. – 2004. – № 2. – С. 3-6.
22. Базанова Л.П. Эколого-физиологические аспекты взаимоотношений сочленов паразитарной триады в Тувинском природном очаге чумы (по экспериментальным исследованиям) / Л.П. Базанова, М.П. Маевский, А.Ф. Попков // Мед. паразитол. – 2004. – № 4. – С. 36-42.
23. Изменчивость и агрегированность возбудителя чумы как способ его сохранения в организме Citellophilus tesquorum altaicus (Siphonaptera) / Л.П. Базанова, А.Я. Никитин, М.П. Маевский, Ю.М. Капустин // Пробл. особо опасных инфекций. – Саратов, 2004. – Вып. 88, № 2. – С. 29-33.
24. Токмакова Е.Г. Образование блока преджелудка, алиментарная активность и смертность у блох Amphipsylla primaris primaris, инфицированных возбудителем чумы / Е.Г. Токмакова, Д.Б. Вержуцкий, Л.П. Базанова // Паразитология. – 2006. – Т. 40, вып. 3. – С. 215-224.
25. Особенности взаимоотношений возбудителя чумы и блох с различных участков Тувинского природного очага / Л.П. Базанова, Д.Б. Вержуцкий, А.Я. Никитин и др. // Мед. паразитол. – 2006. – № 3. – С. 35-38.
26. Эпидемиологическое обоснование санитарной охраны территории Сибири от заноса чумы из Монголии / А.С. Марамович, С.А. Косилко, Г.А. Воронова, Т.И. Иннокентьева, Л.П. Базанова и др. // Пробл. особо опасных инфекций. – 2007. – Вып. 1 (93). – С. 38-43.
27. Базанова Л.П. Трансмиссия возбудителя чумы блохами Frontopsylla luculenta luculenta (Jordan et Rothschild, 1923), их алиментарная активность и смертность / Л.П. Базанова, Г.А. Воронова, Е.Г. Токмакова // Пробл. особо опасных инфекций. – 2007. – Вып. 2 (94). – С. 16-20.
28. Базанова Л.П. Сохранение возбудителя чумы в зимний период самками и самцами Citellophilus tesquorum altaicus / Л.П. Базанова, А.Я. Никитин, М.П. Маевский // Мед. паразитол. – 2007. – № 4. – С. 34-36.
29. Сезонные особенности трансмиссии возбудителя чумы длиннохвостому суслику блохами Citellophilus tesquorum в Туве / Л.П. Базанова, А.Я. Никитин, А.Ф. Попков, М.П. Маевский // Зоол. журн. – 2007. – Т. 86, № 7 – С. – 846-852.
30. Чума в Китае. Опасность заноса в регионы Сибири и Дальнего Востока / А.С. Марамович, С.А. Косилко, Т.И. Иннокентьева, Г.А. Воронова, Л.П. Базанова и др. // Журн. микробиол. – 2008. – № 1. – С. 95-99.
31. Базанова Л.П. О роли блох – основных и второстепенных переносчиков чумы – в циркуляции возбудителя в сибирских природных очагах (обзор) / Л.П. Базанова, Т.И. Инокентьева // Мед. паразитол. – 2008. – № 3. – С. 54-59.
32. Базанова Л.П. К оценке эпизоотологической роли блох длиннохвостого суслика в Тувинском природном очаге чумы / Л.П. Базанова, В.Т. Климов // Мед. паразитол. – 2008. – № 4. – С. 55-59.
Прочие научные статьи и тезисы
33. Базанова Л.П. Взаимоотношения блох монгольской пищухи Тувы с возбудителем чумы основного и алтайского подвидов / Л.П. Базанова, В.Т. Климов // Успехи медицинской энтомологии и акарологии: Матер. Х съезда ВЭО. – Л., 1990. – С.3-4.
34. О переживании зимнего периода блохами Citellophilus tesquorum altaicus инфицированными возбудителем чумы в эксперименте / Л.П. Базанова, М.П. Маевский, И.Ф. Жовтый, А.Г. Вылков, Н.Ф. Галацевич // Матер. регионал. совещания противочумн. учрежд. по эпидемиол., эпизоотол. и профилактике особо опас. инф. (19-20 декабря 1989 г., г. Уральск). – Куйбышев, 1990. – С. 29-30.
35. Изучение возможности инфицирования длиннохвостого суслика возбудителем чумы через поврежденную кожу в эксперименте / Л.П. Базанова, С.В. Сахаров, А.В. Хабаров, С.В. Федоров // Организация эпиднадзора при чуме и меры ее профилактики: Матер. межгосудар. науч.- практ. конф. – Алма-Ата, 1992. – Ч.1. – С. 77-79.
36. Базанова Л.П. Эпизоотологическая оценка блохи Neopsylla mana Wagn. в Тувинском природном очаге чумы по результатам экспериментов / Л.П. Базанова, А.В. Хабаров // Проблемы природно-очаговых и зоонозных инфекций в Сибири и на Дальнем Востоке: Матер. регионал. науч.-практ. конф. – Чита, 1993. – С.11-13.
37. Феномен увеличения способности к трансмиссии у возбудителя чумы при заражении им внутривидовых гибридов Citellophilus tesquorum / А.Я. Никитин, Л.П. Базанова, Л.К. Нечаева и др. // Актуальные проблемы профилактики природно-очаговых инф. болезней: Тез. докл. науч. конф., посвящ. 60-летию Иркут. противочумн. ин-та (октябрь, 1994 г.). – Иркутск, 1994. – С.119-120.
38. Воронова Г.А. О возможном участии триптофанзависимых штаммов чумного микроба в поддержании эпизоотий / Г.А. Воронова, Л.П. Базанова // Журн. инфекц. патологии. – Иркутск, 1998. – Т.5, № 4. – С. 37-38.
39. Базанова Л.П. Способы передачи возбудителя чумы блохой Citellophilus tesquorum altaicus Ioff, 1936 / Л.П. Базанова, М.П. Маевский // Chinese Journal of Control of Endemic Disease. – 1999. – V. 14. – P. 183-185.
40. Оценка эпизоотологической роли блохи Citellophilus tesquorum altaicus в Тувинском природном очаге чумы / Л.П. Базанова, М.П. Маевский, А.Ф. Попков, Н.Ф. Галацевич // Sciertific Journal Centre for Control and Research of Natural Infecticus Diseases. – Ulaanbaatar, 2000. – № 8. – P. 67-73.
41. Базанова Л.П. Межпопуляционные различия во взаимоотношениях с возбудителем чумы двух массовых видов блох длиннохвостого суслика из Тувы / Л.П. Базанова, Д.Б. Вержуцкий, А.В. Хабаров // Карантинные и зоонозные инфекции в Казахстане. – Алматы, 2000. – Вып. 2. – С. 48-52.
42. Базанова Л.П. К оценке эпизоотологической роли блохи Rhadinopsylla li transbaikcalica Ioff et Tifl. (1947) в Тувинском природном очаге чумы / Л.П. Базанова, Д.Б. Вержуцкий // Актуальные проблемы инфектологии и паразитологии: Матер. Первой междунар. юбил. конф. (2-5 апреля 2001 г., г. Томск). – Томск, 2001. – С. 42.
43. Токмакова Е.Г. О влиянии атмосферного давления на алиментарную активность и смертность блох / Е.Г. Токмакова, Л.П. Базанова, Г.А. Воронова // Актуальные проблемы эпидемической безопасности: Матер. юбил. науч.-практ. конф., посвящ. 50-летию Ставропольского противочумн. ин-та (15-16 окт. 2006 г., Ставрополь). – Ставрополь, 2002. – С. 281-282.
44. Базанова Л.П. Половые различия в образовании чумного блока у блох – переносчиков возбудителя чумы / Л.П. Базанова, Е.Г. Токмакова // Проблемы инфекции в клинической медицине: Матер. науч. конф. и VIII съезда Итало-Российского общ-ва по инфекционным болезням (5-6 декабря 2002 г.). – Санкт-Петербург, 2002. – С. 32.
45. Базанова Л.П. Способность блохи Citellophilus tesquorum с неочаговой территории к передаче возбудителя чумы / Л.П. Базанова, А.Я. Никитин // Проблемы современной паразитологии: Матер. междунар. конф. и III съезда Паразитол. об-ва при РАН. СПб.: РАН, 2003. – Ч. I.- С. 58-59.
46. Никитин А.Я. Исследование возможности образования эффективных ксенопар между Yersinia pestis и блохой Citellophilus tesquorum / А.Я. Никитин, Л.П. Базанова // Бюл. ВСНЦ СО РАМН. – Иркутск, 2003. – № 3. – С. 152 – 155.
47. Токмакова Е.Г. Особенности размножения возбудителя чумы в блохах Amphipsylla primaris primaris Jordan et Rothschild, 1915 в различные сезоны года / Е.Г. Токмакова, Л.П. Базанова, Д.Б. Вержуцкий // Журн. инфекц. патологии. – Иркутск, 2003. – Т. 10. – № 4. – С. 106-107.
48. О возможных путях эволюции взаимоотношений блох с возбудителем чумы (на примере сибирских горных очагов) / А.Я. Никитин, Т.И. Иннокентьева, Л.П. Базанова, И.К. Машковский // Противочумн. учрежд. России и их роль в обеспечении эпидем. благополучия населения страны. – М., 2004. – С. 71-75.
49. Изучение скрытой (латентной) фазы эпизоотического процесса в Тувинском природном очаге чумы в эксперименте / Л.П. Базанова, А.Я. Никитин, А.Ф. Попков, М.П. Маевский // Противочумн. учрежд. России и их роль в обеспечении эпидем. благополучия населения страны. – М., 2004. – С. 79-84.
50. Флуктуирующая асимметрия признаков хетотаксии у представителей отряда Siphonaptera / А.Я. Никитин, В.М. Корзун, Е.Г. Токмакова, Л.П. Базанова, Д.Б. Вержуцкий // Успехи современного естествознания. – 2005. – № 3. – С. 56-57.
51. Bazanova L.P. Role of fleas Frontopsylla luculenta luculenta (Jordan et Rothschild, 1923) from the Transbaikalian natural plague focus in the epizootic process / L.P. Bazanova, G.A. Voronova, E.G. Tokmakova // Scientific journal Center for infectious diseases with natural foci. – Ulaanbaatar, 2006. – № 14. – P. 66-74.
52. Некоторые особенности взаимоотношений Xenopsylla cheopis (Rothschild, 1903) и возбудителя чумы с различным плазмидным составом / Е.Г. Токмакова, Г.А. Воронова, Л.П. Базанова, С.В. Балахонов // Матер. I Всеросс. Совещ. по кровосос. насекомым (Санкт-Петербург, 24-27 октября 2006 г.) – Санкт-Петербург, 2006. – С. 197-199.