

Направленной на расширение генетического разнообразия баклажана при селекции на качество
На правах рукописи
ВЕРБА Вадим Михайлович
УДК 635.646 : 631.524.7
РАЗРАБОТКА ЭЛЕМЕНТОВ ТЕХНОЛОГИИ, НАПРАВЛЕННОЙ НА РАСШИРЕНИЕ ГЕНЕТИЧЕСКОГО РАЗНООБРАЗИЯ БАКЛАЖАНА ПРИ СЕЛЕКЦИИ НА КАЧЕСТВО
Специальности: 06.01.05 – селекция и
семеноводство сельскохозяйственных растений
03.00.23 – биотехнология
Автореферат
диссертации на соискание ученой степени
кандидата сельскохозяйственных наук
Москва – 2011
Диссертационная работа выполнена в лаборатории селекции и семеноводства пасленовых культур Всероссийского научно-исследовательского института селекции и семеноводства овощных культур в 2009-2011 годах.
| Научные руководители: доктор сельскохозяйственных наук, профессор | Мамедов Мубариз Иса оглы |
| доктор сельскохозяйственных наук, старший научный сотрудник | Шмыкова Наталья Анатольевна |
| Официальные оппоненты: доктор сельскохозяйственных наук, старший научный сотрудник | Бухаров Александр Фёдорович |
| кандидат сельскохозяйственных наук, старший научный сотрудник | Кан Людмила Юрьевна |
Ведущая организация: Российский государственный аграрный университет - МСХА им. К.А. Тимирязева
Защита диссертации состоится «16» июня 2011 года в «12» часов на заседании совета по защите докторских и кандидатских диссертаций Д 220. 019. 01 при ВНИИ селекции и семеноводства овощных культур (143080, Московская область, Одинцовский район, п/о Лесной городок, пос. ВНИИССОК).
Факс: (495) 599-22-77 e-mail: [email protected]
С диссертацией можно ознакомиться в библиотеке ВНИИССОК
Автореферат разослан «13» мая 2011 года
Ученый секретарь совета
по защите докторских и
кандидатских диссертаций
Д 220. 019. 01
доктор сельскохозяйственных наук,
старший научный сотрудник Пышная О.Н.
Общая характеристика работы
Актуальность темы. В настоящее время проблема селекции сортов и гибридов F1 баклажана c различными параметрами, удовлетворяющих требования производства и потребителей, стоит очень остро. Динамичное развитие общества приводит к увеличению требований потребителей как к качеству и разнообразию продукции, так и к ее объемам. В связи с этим у производителей сельскохозяйственной продукции возникают соответствующие требования к селекционным достижениям, в первую очередь направленные на повышение урожайности и качества продукции (в т.ч. высокое содержание биологически активных веществ (БАВ)). В условиях современного рынка потребительский спрос постоянно растет и меняется, это создает необходимость в наличии у селекционера генотипически разнообразного исходного материала, наличие которого позволит быстро удовлетворять требования современного производства в короткие сроки и создавать линии с высокой комбинационной способностью для селекции на гетерозис.
В связи с этим создание генотипически разнообразного исходного материала с использованием межвидовой гибридизации и разработка методов биотехнологии, способствующих решению ряда проблем при его создании, являются актуальными. Это позволит расширить спектр поиска ценных селекционных форм, повысить эффективность выявления линий с высокими комбинационной способностью и содержанием БАВ для селекции высокоурожайных сортов и гибридов F1 баклажана с широкой нормой реакции к условиям различных культивационных сооружений и открытого грунта.
Цель и задачи и исследований. Цель работы - расширение генотипического спектра изменчивости на основе внутривидовых и межвидовых скрещиваний баклажана для создания разнокачественного исходного материала и отбор перспективных образцов для дальнейшей селекции.
Для реализации данной цели исследований были поставлены следующие задачи:
1.Изучить комбинационную способность и определить эффект гетерозиса по комплексу хозяйственно ценных признаков линий западноазиатского подвида S. melongena L. sp. occidentale Har.
2. Разработать основные элементы методики клонального микроразмножения баклажана (состав питательных сред, типы эксплантов).
3. Провести межвидовые скрещивания и разработать основные элементы методики эмбриокультуры in vitro для зародышей межвидовых гибридов.
4. Оценить полученные межвидовые гибриды по комплексу морфологических, биометрических и биохимических показателей.
Научная новизна работы. Впервые выявлены различия по содержанию фенольных соединений (антоцианы, флавоноиды, фенолкарбоновые кислоты) и пектиновых веществ в отечественных сортах и гибридах, полученных на основе внутри- и межвидовой гибридизации. Установлено, что основным антоцианом кожуры плодов баклажана является дельфинидин-(6-кумароил)-3-глюкозид. Показано, что образцы S.macrocarpon, S.aethiopicum превосходят по содержанию флавоноидов и фенолкарбоновых кислот сортообразцы S.melongena.
Разработана методика клонального микроразмножения отечественных сортов баклажана.
Впервые разработана методика эмбриокультуры с использованием тидиазурона и получены новые генотипы растений на основе межвидовой гибридизации, расширяющие спектр генотипической изменчивости: S.melongena х S.macrocarpon, S.melongena х S.aethiopicum, S.macrocarpon х S.melongena, S.aethiopicum х S.melongena, S.aethiopicum х S.macrocarpon.
Установлены различия в динамике развития зародышей у разных видов баклажана и их гибридов.
Практическая значимость. Выделены селекционно ценные линии с высокой комбинационной способностью и содержанием фенольных соединений. Полученный в результате исследований исходный селекционный материал может быть использован для создания сортов и гибридов F1 с высоким содержанием БАВ.
Подобраны питательные среды для клонального микроразмножения баклажана и эмбриокультуры, определены оптимальные сроки получения полноценных растений-регенерантов из незрелых зародышей.
Основные положения диссертации, выносимые на защиту:
- оценка комбинационной способности линий баклажана по комплексу хозяйственно ценных признаков;
- межвидовая гибридизация баклажана для расширения спектра генотипической изменчивости;
- разработка биотехнологических средств для селекции баклажана.
Апробация работы. Материалы диссертации были доложены на VII Международном симпозиуме по фенольным соединениям (Москва, 2009 год), на Международной ежегодной научно-практической конференции РГАУ-МСХА им. К.А. Тимирязева (2009 - 2010 гг).
Публикации результатов исследований. По материалам диссертации опубликовано 7 печатных работ, 2 из которых в журналах «Вестник РАСХН», «Сельскохозяйственная биология», входящих в перечень рецензируемых научных изданий.
Объем и структура диссертации. Диссертационная работа изложена на __ страницах, состоит из введения, __ глав, выводов, предложений для использования в селекционной практике, списка использованной литературы, содержащего __ наименований, в том числе __ иностранных авторов, приложения, иллюстрирована __ таблицами, __ рисунками.
Материал и методика проведения исследований
Материалом исследований служили селекционные линии S. melongena L. (Л-Беглец, Л-Солярис, Л-Бак, Л-Алмаз, Л-Багратион, Л-Бриллиант) и дикорастущие виды S.aethiopicum L. и S.macrocarpon L. из генофонда лаборатории селекции и семеноводства пасленовых культур ВНИИССОК. Скрещивания проводили по полной диаллельной схеме (рис.1).

Рис 1. Схема диаллельного скрещивания видов рода Solanum L.
Учитывая, что баклажан является факультативным самоопылителем, все скрещивания проводили с кастрацией в фазе хорошо развитых бутонов за день до раскрытия цветка. Пыльцу для опыления собирали с бутонов через день после их раскрытия и до растрескивания пыльников. Опыление выполняли в день кастрации в утренние часы (с 10 до 12) свежесобранной пыльцой с последующей изоляцией опыленных цветков. Завязавшиеся на момент скрещивания плоды и раскрытые цветки удаляли.
Линии, гибриды и межвидовые гибридные комбинации выращивали в вегетационной камере с досвечиванием, пленочной теплице и в зимних обогреваемых теплицах с малообъемной технологией возделывания по общепринятой агротехнике выращивания селекционного материала.
Разработку элементов методик клонального микроразмножения и эмбриокультуры проводили по общепринятым методам работы с культурами растительных тканей (Бутенко, 1984).
Изучение качественного состава антоцианов в кожице плодов баклажана проводили на кафедре химии МГУ и в лаборатории химии пищевых продуктов Института питания РАМН.
Количественное содержание антоцианов определяли в лаборатории биотехнологии и лаборатории применения агрохимических средств в семеноводстве овощных культур ВНИИССОК в соответствии с Руководством по методам контроля качества и безопасности биологически активных добавок к пище (2004), флавоноидов – в соответствии с Gajula et al. (2009), фенолкарбоновых кислот (ФКК) в мякоти – в соответствии с методикой Ларькиной М.С. и др. (2008)
При определении содержания нерастворимых пектинов в мякоти плодов использовали объемный метод, предложенный Раик С.Я. (Волобуева, Шатилова, 2008), в лаборатории токсикологии Института питания РАМН.
Комбинационную способность рассчитывали при помощи пакета статистической обработки результатов диаллельного скрещивания «Агро-Calk». Расчет комбинационной способности проводился по I методу Б. Гриффинга.
Статистическую обработку данных проводили по Доспехову Б.А. (1985) с использованием пакета прикладных программ MICROSOFT EXCEL 7,0.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
1. Оценка эффекта гетерозиса у гибридов F1 баклажана по
количественным признакам
В 2009 – 2010 гг. проведена оценка гибридов F1, полученных при скрещивании 5 линий баклажана. Изучение родительских линий показало, что они отличались разнообразием по всем признакам, кроме раннеспелости. У гибридов F1 разнообразие было отмечено по всем учитываемым признакам, в том числе и по раннеспелости – ряд гибридов превосходили родительские линии по раннеспелости на 10-20 суток.
По признаку «ранняя урожайность» эффект гетерозиса варьировал в пределах 101,9 – 391,3%, по признаку «общая урожайность» – в пределах 108,2 – 295,1%. Таким образом, проявление эффекта гетерозиса по ранней урожайности, в наших исследованиях, было выше, чем по общей (табл. 1).
Лучшим по ранней урожайности оказался гибрид F1 Л-Беглец х Л-Бак - 6,8 кг/м2 (эффект гетерозиса - 164,9%). Обе линии имели одинаково высокие показатели по ранней урожайности. Гибридная комбинация F1 Л-Алмаз х Л-Багратион, имевшая высокий эффект гетерозиса - 189,5%, была получена при скрещивании линий контрастных по ранней урожайности.
В целом, при анализе других гибридных комбинаций четкой закономерности в проявлении эффекта гетерозиса в зависимости от показателей родительских форм выявить не удалось. Гетерозис проявлялся как в гибридах с участием родительских линий с резко контрастными, так и с близкими по величине показателями.
Таблица 1
Эффект гетерозиса по ранней и общей урожайности у гибридов F1
баклажана (2009-2010 годы)
| № № п/п | Линии и гибридные комбинации | Ранняя урожайность | Общая урожайность | ||
| кг/м2 | гетерозис,% | кг/м2 | гетерозис,% | ||
| 1 | Л-Багратион | 2,8 | - | 11,0 | - |
| 2 | Л-Бак | 4,0 | - | 14,7 | - |
| 3 | Л-Алмаз | 0,7 | - | 4,1 | - |
| 4 | Л-Солярис | 0,7 | - | 2,7 | - |
| 5 | Л-Беглец | 4,1 | - | 8,9 | - |
| 6 | F1 Л-Багратион х Л-Бак | 4,3 | 107,8 | 15,9 | 108,2 |
| 7 | F1 Л-Багратион х Л-Алмаз | 4,6 | 166,7 | 15,8 | 143,6 |
| 8 | F1 Л-Багратион х Л-Солярис | 3,4 | 123,9 | 12,5 | 113,6 |
| 9 | F1 Л-Багратион х Л-Беглец | 6,1 | 147,0 | 17,7 | 160,9 |
| 10 | F1 Л-Бак х Л-Багратион | 3,9 | 99,5 | 14,1 | 95,9 |
| 11 | F1 Л-Бак х Л-Алмаз | 4,2 | 105,8 | 14,7 | 100,0 |
| 12 | F1 Л-Бак х Л-Солярис | 3,1 | 78,0 | 12,2 | 83,0 |
| 13 | F1 Л-Бак х Л-Беглец | 6,4 | 153,8 | 17,0 | 115,6 |
| 14 | F1 Л-Алмаз х Л-Багратион | 5,2 | 189,5 | 17,0 | 154,5 |
| 15 | F1 Л-Алмаз х Л-Бак | 1,1 | 26,8 | 6,7 | 45,6 |
| 16 | F1 Л-Алмаз х Л-Солярис | 2,0 | 289,9 | 12,1 | 295,1 |
| 17 | F1 Л-Алмаз х Л-Беглец | 4,2 | 101,9 | 10,9 | 122,5 |
| 18 | F1 Л-Солярис х Л-Багратион | 3,5 | 128,3 | 12,1 | 110,0 |
| 19 | F1 Л-Солярис х Л-Бак | 4,5 | 114,9 | 13,3 | 90,5 |
| 20 | F1 Л-Солярис х Л-Алмаз | 2,7 | 391,3 | 9,8 | 239,0 |
| 21 | F1 Л-Солярис х Л-Беглец | 3,0 | 71,7 | 12,0 | 134,8 |
| 22 | F1 Л-Беглец х Л-Багратион | 6,0 | 144,8 | 19,1 | 173,6 |
| 23 | F1 Л-Беглец х Л-Бак | 6,8 | 164,9 | 16,9 | 115,0 |
| 24 | F1 Л-Беглец х Л-Алмаз | 5,1 | 124,2 | 16,1 | 180,9 |
| 25 | F1 Л-Беглец х Л-Солярис | 4,2 | 102,2 | 8,9 | 100,0 |
По признаку «общая урожайность» наиболее высокий эффект гетерозиса был у гибридов с участием тех же линий - Л-Солярис и Л-Алмаз. Однако по абсолютному показателю они так же уступали гибридам, у которых эффект гетерозиса был ниже. В целом, 70% гибридов показали превышение по этому признаку над лучшей родительской линией.
Наиболее высокоурожайные гибриды были получены при скрещивании линий с высокими и средними показателями: F1 Л-Багратион х Л-Беглец, F1 Л-Бак х Л-Беглец, F1 Л-Беглец х Л-Багратион, F1 Л-Беглец х Л-Бак. Урожайность их составила 16,9 – 19,1 кг/м2. Эффект гетерозиса при этом был 115,0 -160,9%.
По признаку «число плодов на растении» гетерозис был выраженным. Варьирование эффекта гетерозиса по признакам «общее» и «товарное число плодов на растении» составило 101,0 – 160,1% и 102,2 – 246,9%, соответственно.
Таким образом, максимальные абсолютные значения, средний и высокий уровень эффекта гетерозиса проявлялись в скрещиваниях, соответствующих схеме высокий х средний или средний х средний уровень признака у родительских линий. Резко контрастные линии в скрещиваниях реже показывали высокий гетерозис и абсолютные значения признака. Однако, четких закономерностей проявления эффекта гетерозиса не наблюдалось.
По признаку «продолжительность периода «всходы-техническая спелость» набор родительских форм был достаточно однообразным. Линии, в результате скрещивания которых были получены более раннеспелые формы, не обладали существенно короткими межфазными периодами «всходы-цветение» и «цветение-созревание». Например, у наиболее раннеспелого гибрида F1 Л-Алмаз х Л-Солярис родительские линии проходили эти фазы за примерно одинаковый срок.
2. Комбинационная способность линий баклажана по количественным признакам
Оценка линий проведена по признакам урожайности (ранняя и общая), числу плодов на растении (общее и товарное) и продолжительности периода «всходы-техническая спелость».
При дисперсионном анализе оценка по критерию F показала, что общая комбинационная способность (ОКС) оказывала существенное влияние на все признаки, но в значительной степени на раннюю и общую урожайность и число плодов на растении (общее и товарное); в меньшей степени на раннеспелость. Специфическая комбинационная способность (СКС) также оказывала существенное влияние на все изучаемые признаки, но в большей степени на число товарных плодов и общую урожайность. Кроме того, дисперсионный анализ выявил высокую долю эффекта ОКС, которая превосходила эффект СКС в 2-4 раза по всем изучаемым признакам, кроме признака «продолжительность периода «всходы-техническая спелость», где эффект СКС был выше эффекта ОКС почти в два раза.
Сравнение эффектов ОКС (gi) показало, что они существенны по всем признакам. Л-Багратион и Л-Бак обладали существенной ОКС по таким важным признакам как «общая урожайность» и «число товарных плодов на растении». Л-Алмаз, обладая высокой ОКС по признаку «продолжительность периода «всходы-техническая спелость», отличалась существенно низкой ОКС по всем остальным признакам. Л-Беглец, обладая наибольшей ОКС по признакам «ранняя урожайность» и «общее число плодов на растении», уступала Л-Багратион и Л-Бак по признакам «общая урожайность» и «число товарных плодов на растении».
Не установлено четкой связи между уровнем ОКС и средним значением признака у исходных линий. Наблюдалось некоторое сходство между максимальными и минимальными значениями признака с уровнем ОКС по всем признакам, кроме признака «продолжительность периода «всходы-техническая спелость». Изучение варианс специфической комбинационной способности показало, что различия СКС по признаку «ранняя урожайность» несущественны. Наибольшей специфической комбинационной способностью по этому признаку обладала линия Л-Алмаз.
По признаку «общая урожайность» существенной максимальной СКС обладала только линия Л-Алмаз. Несколько меньше она была у Л-Багратион. У остальных изученных линий СКС по этому признаку была несущественной.
Вариансы СКС более контрастны по признаку «общее число плодов» и «число товарных плодов». Существенно высокой СКС по этим признакам обладала Л-Багратион, несколько меньше она была у Л-Алмаз.
По признаку «продолжительность периода «всходы-техническая спелость» существенно высокой СКС обладали две линии Л-Алмаз и Л-Солярис.
Анализ величин общей и специфической комбинационной способности показал, что ни одна линия не обладает существенными эффектами СКС и ОКС по всем изученным признакам (табл. 2).
Таблица 2
Сочетание высоких ОКС и СКС у линий баклажана по количественным
признакам (2009-2010 годы)
| Название линий | Наличие (+) высокой ОКС и СКС по признакам | |||||||||
| Урожайность | Число плодов на растении | Продолжительность периода «всходы-техническая спелость» | ||||||||
| ранняя | общая | общее | товарных | |||||||
| ОКС | СКС | ОКС | СКС | ОКС | СКС | ОКС | СКС | ОКС | СКС | |
| Л-Багратион | + | + | + | + | + | + | ||||
| Л-Бак | + | + | + | + | + | |||||
| Л-Алмаз | + | + | + | + | + | + | ||||
| Л-Солярис | + | |||||||||
| Л-Беглец | + | + | + | + | ||||||
Сочетание в Л-Алмаз эффектов ОКС и СКС по признаку «продолжительность периода «всходы-техническая спелость» существенно сказывалось на раннеспелости гибридов с участием этой линии. Сочетание высоких эффектов СКС в Л-Алмаз и Л-Солярис позволило получить гибрид со значительной раннеспелостью – 80-85 суток. Эффект СКС по этому признаку проявляется только в том случае, если Л-Алмаз выступал в качестве материнского компонента в скрещиваниях. В то же время Л-Солярис, не обладая значительной ОКС по этому признаку, во всех остальных гибридных комбинациях не влияла на раннеспелость гибридов.
По другим признакам наблюдалась аналогичная закономерность. Так Л-Беглец, Л-Бак и Л-Багратион, обладая высокой ОКС по признакам «ранняя» и «общая урожайность» в гибридных комбинациях, показали положительные результаты – большинство высокоурожайных гибридов F1 были получены с участием этих линий. Особенно сильно действие ОКС проявилось в гибриде F1 Л-Беглец х Л-Багратион.
Результаты анализа линий по ОКС и СКС позволили отобрать линии с высокой комбинационной способностью, которые в дальнейшем могут послужить для создания высокогетерозисных гибридов F1: Л-Багратион, Л-Бак могут использоваться для получения более высокоурожайных гибридов. Линия Л-Алмаз, обладая высокой ОКС и СКС по признаку «продолжительность периода «всходы-техническая спелость», может быть использована для получения более раннеспелых гибридов, либо для создания популяций при селекции на раннеспелость.
3. Биохимический состав плодов баклажана
Оценка линий и гибридов F1 баклажана вида S.melongena по содержанию фенольных соединений (флавоноиды и фенолкарбоновые кислоты) в мякоти плодов показала, что их суммарное содержание варьирует в пределах от 1,7 до 2,4% (табл. 3). У линий суммарное содержание фенольных соединений изменялось в пределах от 1,8 до 2,2%. Среди гибридов F1 наибольшее содержание суммарного количества фенольных соединений было у следующих гибридов: F1 Л-Беглец х Л-Солярис – 2,3%, F1 Л-Беглец х Л-Алмаз – 2,3%, F1 Л-Алмаз х Л-Беглец – 2,3%. У дикорастущих видов баклажана S.aethiopicum и S.macrocarpon общее содержание исследуемых фенольных соединений было в 1,5 раза выше, чем у S. melongenа - до 2,9 – 3%.
Содержание флавоноидов в исследованных нами гибридах F1 и линиях баклажана варьировало в пределах 0,7 – 1,1%. Виды S.macrocarpon и S.aethiopicum содержали большее количество флавоноидов: S.aethiopicum – до 1,4%, S.macrocarpon – до 1,2%.
Общее содержание фенолкарбоновых кислот в линиях и гибридах F1 вида S.melongena было 1,0-1,5%, S.aethiopicum - 1,5%, S.macrocarpon – 1,8%.
Содержание нерастворимых пектинов в линиях и гибридах F1 S.melongena составило 0,27 – 0,63%, в S.aethiopicum и S.macrocarpon оно было незначительно выше, чем у S.melongena и составило 0,51 – 0,64%.
Хроматографический анализ качественного состава антоцианов показал, что для всех исследуемых линий и гибридов F1 характерно наличие одинакового качественного состава антоцианов в кожице плодов (рис. 2). Все пики имели одинаковое время удерживания и различались только площадью.
Определение химической структуры антоцианов в кожуре плодов линий и гибридов F1 показало, что основным антоцианом является дельфинидин-(6-кумароил)-3-глюкозид (рис. 3).
Таблица 3
Содержание фенольных соединений в мякоти плодов различных видов
баклажана (S. melongena, S.macrocarpon, S. aethiopicum) (2010 – 2011 годы)
| №№ п/п | Линия, гибридная комбинация | Cодержание фенольных соединений,% | Содержание нерастворимых пектинов,% | |||
| Содержание антоцианов в кожуре плодов | Содержание ФКК в мякоти плодов | Содержание флавоноидов в мякоти плодов | Общее (4+5) | |||
| 2 | 3 | 4 | 5 | 6 | 7 | |
| Линии и гибриды F1 S.melongena | ||||||
| 1 | F1 Л-Багратион х Л-Бак | 0,19 ± 0,03 | 1,4 ± 0,0 | 0,8 ± 0,0 | 2,2 | 0,48 |
| 2 | F1 Л-Багратион х Л-Алмаз | 0,17 ± 0,02 | 1,2 ± 0,0 | 0,7 ± 0,1 | 1,9 | 0,45 |
| 3 | F1 Л- Багратион х Л-Солярис | 0,14 ± 0,02 | 1,3 ± 0,0 | 0,9 ± 0,1 | 2,2 | 0,30 |
| 4 | F1 Л-Багратион х Л-Беглец | 0,12 ± 0,02 | 1,2 ± 0,0 | 1,0 ± 0,1 | 2,2 | 0,32 |
| 5 | F1 Л-Бак х Л-Багратион | 0,15 ± 0,06 | 1,4 ± 0,0 | 0,7 ± 0,1 | 2,1 | 0,35 |
| 6 | F1 Л-Бак х Л-Алмаз | 0,25 ± 0,03 | 1,4 ± 0,0 | 0,7 ± 0,0 | 2,1 | 0,32 |
| 7 | F1 Л-Бак х Л-Солярис | 0,17 ± 0,02 | 1,4 ± 0,0 | 0,7 ± 0,0 | 2,1 | 0,46 |
| 8 | F1 Л-Бак х Л-Беглец | 0,13 ± 0,01 | 1,0 ± 0,0 | 0,7 ± 0,1 | 1,7 | 0,42 |
| 9 | F1 Л-Алмаз х Л-Багратион | 0,12 ± 0,02 | 1,0 ± 0,0 | 0,8 ± 0,1 | 1,8 | 0,63 |
| 10 | F1 Л-Алмаз х Л-Бак | 0,19 ± 0,04 | 1,2 ± 0,0 | 0,7 ± 0,1 | 1,9 | 0,38 |
| 11 | F1 Л-Алмаз х Л-Солярис | 0,33 ± 0,05 | 1,4 ± 0,0 | 0,8 ± 0,1 | 2,2 | 0,35 |
| 12 | F1 Л-Алмаз х Л-Беглец | 0,14 ± 0,02 | 1,2 ± 0,0 | 1,1 ± 0,0 | 2,3 | 0,45 |
| 13 | F1 Л-Солярис х Л-Багратион | 0,16 ± 0,00 | 1,4 ± 0,0 | 0,8 ± 0,0 | 2,2 | 0,33 |
| 14 | F1 Л-Солярис х Л-Бак | 0,17 ± 0,03 | 1,1 ± 0,0 | 1,0 ± 0,1 | 2,1 | 0,44 |
| 15 | F1 Л-Солярис х Л-Алмаз | 0,22 ± 0,06 | 1,3 ± 0,0 | 0,7 ± 0,1 | 2,0 | 0,43 |
| 16 | F1 Л-Солярис х Л-Беглец | 0,09 ± 0,02 | 1,5 ± 0,0 | 0,7 ± 0,1 | 2,2 | 0,40 |
| 17 | F1 Л-Беглец х Л-Багратион | 0,11 ± 0,01 | 1,1 ± 0,0 | 0,8 ± 0,0 | 1,9 | 0,40 |
| 18 | F1 Л-Беглец х Л-Бак | 0,12 ± 0,03 | 1,0 ± 0,0 | 0,7 ± 0,0 | 1,7 | 0,27 |
| 19 | F1 Л-Беглец х Л-Алмаз | 0,18 ± 0,02 | 1,4 ± 0,0 | 0,9 ± 0,0 | 2,3 | 0,37 |
| 20 | F1 Л-Беглец х Л-Солярис | 0,22 ± 0,03 | 1,3 ± 0,0 | 1,0 ± 0,0 | 2,3 | 0,40 |
| 21 | Л-Беглец | 0,11 ± 0,02 | 1,1 ± 0,0 | 0,7 ± 0,0 | 1,7 | 0,44 |
| 22 | Л-Багратион | 0,18 ± 0,02 | 1,3 ± 0,0 | 0,9 ± 0,1 | 2,2 | 0,46 |
| 23 | Л-Бак | 0,16 ± 0,05 | 1,0 ± 0,0 | 0,8 ± 0,1 | 1,8 | 0,37 |
| 24 | Л-Алмаз | 0,12 ± 0,05 | 1,0 ± 0,0 | 0,8 ± 0,0 | 1,8 | 0,48 |
| 25 | Л-Солярис | 0,11 ± 0,01 | 1,3 ± 0,0 | 0,8 ± 0,1 | 2,1 | 0,47 |
| 26 | Л-Бриллиант | 0,13 ± 0,04 | 1,0 ± 0,0 | 0,8 ± 0,1 | 1,8 | 0,58 |
| Дикорастущие виды | ||||||
| 27 | S.aethiopicum | - | 1,5 ± 0,0 | 1,4 ± 0,0 | 2,9 | 0,64 |
| 28 | S.macrocarpon | - | 1,8 ± 0,0 | 1,2 ± 0,1 | 3,0 | 0,51 |

Рис. 2. Разделение смеси антоцианов, содержащихся в кожуре плодов баклажана при 520 нм (анализ 23.06.10 г) (снизу вверх): F1 Багира, Л-Багратион, Л-Бак, Л-Алмаз, Л-Солярис, Л-Беглец.

Рис. 3. Результаты определения химической структуры антоциана, содержащего в кожуре плодов S.melongena, на масс-детекторе (TOF-MS) (520 нм).
Количество антоцианов в кожуре плодов гибридов F1 и линий варьировало в пределах 0,09 - 0,33% (табл. 3).
В пересчете на 1 кг плодов содержание антоцианов в линиях и гибридах F1 составило в среднем 0,12 – 0,31 г/кг. 60% гибридов F1 по этому признаку не превышали родительские линии. У 40% гибридов наблюдался эффект гетерозиса по данному признаку 105,6 – 193,8%.
Более высокое содержание флавоноидов и фенолкарбоновых кислот в дикорастущих видах баклажана говорит о необходимости вовлечения их в селекционный процесс, для повышения содержания их в селекционных образцах.
4. Разработка методики клонального микроразмножения
Одним из методов преодоления межвидовых барьеров является эмбриокультура, однако в основе всех биотехнологических методов лежит клональное микроразмножение.
При подборе оптимальных сред для инициации каллусообразования использовали 7 вариантов сред. В качестве основной среды использовали среду Мурасиге и Скуга (МС) (Murashige, Skoog, 1962) с добавлением регуляторов роста: 0,2 мг/л ТДZ; 0,02 мг/л ТДZ; 0,2 мг/л БАП; 2,0 мг/л БАП; 2,0 мг/л НУК; 0,2 мг/л кинетина; 2,0 мг/л кинетина.
Через 3-4 недели экспланты наиболее интенсивно развивались на среде с добавлением 0,02 мг/л и 0,2 мг/л ТДZ. У 100% эксплантов семядолей формировался плотный темно- и светло-зеленый каллус с множеством почек (3-30 шт./эксплант). У 80-90% эксплантов начинали развиваться микророзетки. Для 90-100% эксплантов гипокотиля было характерно образование белого рыхлого каллуса с почками, побеги чаще формировались в верхней части гипокотиля. Незначительное число почек и побегов развилось в вариантах с регулятором роста БАП (0,2 и 2,0 мг/л) и НУК (2,0 мг/л). Единичные побеги образовывались на среде с 0,2 мг/л кинетина на эксплантах гипокотиля. Экспланты не развивались на средах с 2,0 мг/л кинетина.
Подсчет числа почек и побегов у эксплантов сорта Снежный через 15 суток после введения в культуру in vitro показал, что наибольшее их число образовалось на каллусе семядолей в варианте с 0,02 мг/л ТДZ – 39,9 шт./эксплант и 13,9 шт./эксплант, соответственно. В варианте с 0,2 мг/л ТДZ образовывалось 17,2 шт./эксплант почек и 9,1 шт./эксплант побегов.
В связи с тем, что большинство почек не тронулись в рост даже через 28 суток после начала культивирования, возникла необходимость в переносе почек с каллусом на регенерационные среды. Для этих целей использовали среду МС с пониженной концентрацией ТДZ в комбинации с БАП, так как ТДZ в высоких концентрациях вызывает витрификацию и значительно снижает процент укоренившихся побегов (Magioli et al., 1998a, Franklin et al., 2004).
Через 15 суток наибольшее число почек каллуса семядолей развивалось при переносе с индукционной среды, содержащей 0,2 мг/л ТДZ, на регенерационные среды, содержащие 0,01 мг/л ТДZ + 0,01 мг/л БАП и 0,01 мг/л ТДZ, - 39,2 шт./эксплант и 38,8 шт./эксплант, соответственно (табл. 4, рис. 4а). При переносе с индукционной среды, содержащей 0,02 мг/л ТДZ, развивалось в среднем 17-17,6 шт./эксплант. В остальных вариантах число побегов было незначительное.
Таблица 4
Число побегов (шт./эксплант) баклажана (S.melongena, сорт Снежный),
развившихся на каллусе семядолей на регенерационных средах
(через 15 суток), (2009 год)
| Регулятор роста, мг/л в регенерационной среде | Индукционная среда (регулятор роста, мг/л) | |||
| 0,2 ТДZ | 0,02 ТДZ | 0,2 БАП | 2,0 НУК | |
| 0,01 ТДZ + 0,01БАП | 39,2±3,6 | 17,6±2,1 | 1,7±2,8 | 0,2±0,4 |
| 0,01 ТДZ | 38,8±2,8 | 17,0±3,8 | 5,1±2,6 | 0,0±0,0 |
| Безгормональная | 10,0±6,3 | 5,8±2,9 | 1,8±1,6 | 0,0±0,0 |

Рис. 4. Образование почек и развитие побегов баклажана (S. melongena – сорт Снежный) на различных эксплантах, культивируемых на среде МС с добавлением 0,02 мг/л тидиазурона; а - образование почек и развитие побегов на семядольном экспланте, б - побегообразование из участка гипокотиля.
Важным этапом работы был подбор среды для укоренения полученных побегов. Так как в эксперименте для регенерации использовались сниженные концентрации тидиазурона, побеги, развившиеся из почек каллуса семядолей, укоренялись с частотой 85% после двукратной пересадки на среду МС половинным содержанием основных компонентов ( МС) без добавления регуляторов роста с интервалом полторы-две недели. На среде МС без добавления регуляторов роста укоренение шло с частотой 76%.
Почки и микророзетки из почек рыхлого каллуса гипокотиля, через две недели культивирования на регенерационных средах были слабо развиты, поэтому их отделяли от каллуса и повторно высаживали на аналогичные среды (0,01 мг/л ТДZ и 0,01 мг/л ТДZ + 0,01 мг/л БАП) для пролонгации. Через две недели наблюдалось развитие побегов 0,5 – 2,0 см длиной (рис. 4б).
Для укоренения и снижения витрификации полученные побеги отделяли от каллуса и дважды (с интервалом 14 суток) пересаживали на среду МС и МС без добавления регуляторов роста. Исключение регуляторов роста на этом этапе позволило снизить степень эндогенных цитокининов и, тем самым, уменьшить процент витрифицированных растений. Через 30 суток на среде МС побеги укоренялись с частотой 78%, на среде МС – 66% (табл. 5, рис. 5а, б).
После акклиматизации полученные растения не имели отклонений по фенотипу и аномалий развития.
Таблица 5
Результаты укоренения побегов баклажана (S.melongena, сорт Солярис), полученных из каллуса гипокотиля, на индукционных средах с добавлением 0,2 и 0,02 мг/л ТДZ (2009 год)
| Сутки | Число розеток, шт. | ||||||
| общее | погибших | укоренив- шихся | общее | погибших | укоренив- шихся | ||
| MS | MS | ||||||
| 15 | Всего | 50 | 2 | 12 | 50 | 5 | 26 |
| % | 4 | 24 | 10 | 52 | |||
| 30 | Всего | 50 | 5 | 33 | 50 | 9 | 39 |
| % | 10 | 66 | 18 | 78 | |||
Таким образом, лучшей для введения эксплантов сортообразцов Солярис и Снежный в культуру in vitro была среда Мурасиге и Скуга с добавлением ТДZ концентраций 0,02 и 0,2 мг/л. Для регенерации побегов оптимальными были среды с уменьшенной концентрацией тидиазурона (0,01 мг/л ТДZ и 0,01 мг/л ТДZ +0,01 мг/л БАП). Укоренение побегов происходило лучше на среде МС.

Рис. 5. Укоренение побегов баклажана (S. melongena – сорт Солярис) на среде МС с половинной концентрацией компонентов без регуляторов роста: 1 – начало укоренения, 2 – укоренившиеся побеги, готовые к посадке in vivo.
Полученные данные позволяют говорить о том, что тидиазурон является перспективным регулятором роста при использовании его для культуры тканей баклажана.
5. Разработка методики культивирования in vitro зародышей межвидовых гибридов
После определения регулятора роста, на который ткани баклажана имели хороший отклик, была начата разработка методики эмбриокультуры. Важным при этом является знание стадий и динамики развития исходных видов и межвидовых гибридов. В исследовании учитывали 5 стадий развития: глобулярный эмбриоид, сердечковидный эмбриоид, торпедовидный эмбриоид, почти сформировавшийся зародыш, сформировавшийся зародыш (рис. 6).

Рис. 6. Стадии развития зародышей представителей рода Solanum L (S.melongena, S.aethiopicum, S.macrocarpon): 1 – глобулярный эмбриоид; 1а - глобулярный эмбриоид с зародышевым мешком; 2 – сердечковидный эмбриоид; 3 – торпедовидный эмбриоид; 4 – почти сформировавшийся зародыш; 5 – сформировавшийся зародыш.
Наиболее быстро развитие зародыша шло у вида S.aethiopicum – зародыш формировался на 30 сутки после опыления, а все учитываемые стадии развития проходили за 10-12 суток. У S.melongena зародыш полностью формировался на 40, а у S.macrocarpon – на 45 сутки после опыления (табл. 6).
Таблица 6
Динамика и стадии развития зародышей у представителей рода Solanum L. (S.melongena, S.aethiopicum, S.macrocarpon) (2009 год)
| Вид | Время после опыления, сутки | Размер зародышей (длина), мм | Стадия развития зародыша |
| S.melongena (Л-Алмаз, Л-Бриллиант) | 20 | 0,032±0,001 | глобулярный эмбриоид |
| 23 | 0,10±0,01 | сердечковидный эмбриоид | |
| 27 | 0,60±0,06 | торпедовидный эмбриоид | |
| 31 | 1,20±0,06 | торпедовидный эмбриоид | |
| 35 | 3,80±0,09 | почти сформировавшийся зародыш | |
| 40 | 4,30±0,09 | сформировавшийся зародыш | |
| S. aethiopicum | 20 | 0,080±0,007 | глобулярный – сердечковидный эмбриоид |
| 25 | 2,2±0,3 | торпедовидный эмбриоид - почти сформировавшийся зародыш | |
| 30 | 4,10±0,05 | сформировавшийся зародыш | |
| S.macrocarpon | 20 | 0,070±0,005 | глобулярный эмбриоид |
| 25 | 0,15±0,03 | сердечковидный эмбриоид | |
| 30 | 1,1±0,1 | торпедовидный эмбриоид | |
| 35 | 1,8±0,2 | торпедовидный эмбриоид | |
| 40 | 3,9±0,2 | почти сформировавшийся зародыш | |
| 45 | 4,5±0,1 | сформировавшийся зародыш |
Скорость прохождения учитываемых фаз у межвидовых гибридов отличалась от скорости развития зародышей у исходных видов. В гибридах с S.aethiopicum (в качестве отцовского компонента), зародыши развивались быстрее, а у гибридов с S.melongena (в качестве отцовского компонента) развитие зародышей шло более медленно. В гибридах с S.macrocarpon в качестве отцовского компонента такого эффекта не наблюдалось. Размеры зародышей у межвидовых гибридов не имели значительных отличий от размеров у родительских линий.
Для культивирования недозрелых зародышей основополагающим является состав питательной среды для индукции развития зародышей после изоляции. При подборе среды использовалась среда МС с добавлением двух регуляторов роста: тидиазурона (0,1 мг/л) и НУК (10 мг/л).
Развитие зародышей после высадки на среды имело общие закономерности. У всех трех видов и межвидовых гибридов видов развитие у 20-суточных зародышей не наблюдалось. 25–30-суточные зародыши на среде с 0,1 мг/л ТДZ развивались в нормальные растения только у вида S. aethiopicum и межвидовых гибридов с его участием. У видов с S. melongena и S.macrocarpon и полученных с их участием межвидовых гибридов – начиная с 35-суточного возраста. На среде с 0,1 мг/л ТДZ зародыши через 1,5-2 недели развивали нормальные семядоли (рис. 7).

Рис. 7. Развитие зародышей (возраст 35 суток) гибрида S. melongena (Л-Бриллиант) х S. aethiopicum после посадки на индукционную среду МС + 0,1 мг/л ТДZ.
На среде с добавлением 10 мг/л НУК зародыши в возрасте 20-25 суток не развивались. 30-40-суточные зародыши в большинстве комбинаций развивались очень слабо – на них образовывался белый рыхлый каллус и ненормально развитые семядоли, без точки роста и корней или, наоборот, с мощным главным корнем без придаточных корешков (рис. 8а, б).

Рис. 8. Развитие зародышей (возраст – 35 суток) гибрида S. melongena (Л-Бриллиант) х S. aethiopicum после посадки на индукционную среду МС + 10 мг/л НУК: а – через 4-5 суток после начала культивирования, б – через 10-14 суток.
Зародыши комбинации S.macrocarpon х S. aethiopicum для культивирования получить не удалось. Плоды не завязывались или формировались очень мелкие и без семян.
Учет выживаемости показал, что в большинстве комбинаций у зародышей в возрасте 20-25 суток (глобулярный - торпедовидный эмбриоид) она была низкой (0-10%) на обеих средах (табл. 7). У зародышей возрастом 30-40 суток (торпедовидный эмбриоид – полностью сформировавшийся зародыш) частота выживаемости была на уровне 50-100%.
Через 1,5-2 недели зародыши с развившимися семядольными листочками пересаживали на среду МС для укоренения.
Таблица 7
Выживаемость зародышей межвидовых гибридных комбинаций баклажана на индукционной среде через 2 недели после введения в культуру in vitro (среда МС с добавлением регуляторов роста) (2009 год)
| Гибрид | Сутки после опыления | Выживаемость,% | |
| 0,1 мг/л ТДZ | 10 мг/л НУК | ||
| S.macrocarpon х S.melongena (Л-Бриллиант) | 25 | 0 | 0 |
| 30 | 100 | 50 | |
| 35 | 100 | 100 | |
| 40 | 50 | 100 | |
| S.aethiopicum х S.melongena (Л-Бриллиант) | 20 | 10 | 0 |
| 25 | 90 | 0 | |
| 30 | 100 | 100 | |
| 35 | 60 | 50 | |
| S.melongena (Л-Бриллиант) х S.aethiopicum | 20 | 0 | 0 |
| 25 | 0 | 0 | |
| 30 | 100 | 100 | |
| 35 | 100 | 100 | |
| S.melongena (Л-Бриллиант) х S.macrocarpon | 25 | 0 | 0 |
| 30 | 100 | 100 | |
| 35 | 100 | 100 | |
| 40 | 100 | 100 | |
| S.aethiopicum х S.macrocarpon | 20 | 0 | 0 |
| 25 | 0 | 0 | |
| 30 | 60 | 60 | |
| 40 | 80 | 100 | |
| Среднее | 57,5 | 53 | |
В среднем процент укоренившихся и пригодных к пересадке в почву растений составил 69% для зародышей, развивавшихся на среде с добавлением 0,1 мг/л ТДZ и 9,6% на среде с добавлением 10 мг/л НУК. Процент укоренения зародышей возрастом 35 и более суток в большинстве комбинаций был выше 80%.
6. Оценка полученных родительских форм и межвидовых гибридов по морфологическим признакам, биометрическим и биохимическим показателям
В 2009-2010 годах проведено изучение полученных межвидовых гибридов баклажана по комплексу биометрических, морфологических показателей и содержанию фенольных соединений (табл.8, 9).
Таблица 8
Биометрическая характеристика видов и межвидовых гибридов баклажана рода Solanum L.(2009-2010 годы)
| №№№ п/п | Вид, гибридная комбинация | Длина междоузлия, см | Параметры листа | Параметры плода | |||
| Длина, см | Ширина, см | Длина черешка, см | Диаметр, см | Масса, г | |||
| 1 | S.melongena | 13,2 ± 1,3 | 36,6 ± 1,5 | 27,3 ± 1,4 | 13,8 ± 0,6 | 7,3±0,9 | 174,6±5,8 |
| 2 | S.macrocarpon | 6,5 ± 0,8 | 38,3 ± 1,7 | 28 ± 1,5 | 3,9 ± 0,4 | 3,2±0,2 | 26,9±2,5 |
| 3 | S.aethiopicum | 11,0 ± 0,9 | 35,3 ± 1,3 | 26,4 ± 1,2 | 12,5 ± 0,4 | 3,7±0,3 | 44,4±3,6 |
| 4 | S.macrocarpon x S.melongena (Л-Бриллиант) | 2,3 ± 0,6 | 16,6 ± 2,4 | 9,2 ± 1,6 | 6,0 ± 0,5 | 2,7±0,3 | 25-35 |
| 5 | S.melongena (Л-Бриллиант) х S.macrocarpon | 1,7 ± 0,3 | 14,4 ± 0,9 | 7,5 ± 0,4 | 5,8 ± 0,3 | 3,2±0,2 | 25-35 |
| 6 | S.melongena (Л-Бриллиант) х S.aethiopicum | 8,5 ± 0,9 | 32,0 ± 2,7 | 27,2 ± 3,4 | 12,4 ± 0,9 | 2,6±0,1 | 25-30 |
| 7 | S.aethiopicum х S.melongena (Л-Бриллиант) | 7,9 ± 0,5 | 39,4 ± 2,0 | 33,1 ± 2,3 | 13,4 ± 0,5 | 4,1±0,1 | 30-50 |
| 8 | S.aethiopicum х S.macrocarpon | 2,5 ± 0,2 | 11,4 ± 1,3 | 6,9 ± 0,9 | 2,8 ± 0,2 | 1,7±0,1 | 4 - 6 |
Установлено, что гибриды первого поколения в основном наследуют признаки дикорастущих видов по таким признакам как форма, размер, масса плода. У ряда комбинаций проявлялись шипы, которые отсутствовали у родительских форм.
По таким признакам как габитус растения, форма листа и опушенность, наследование зависело от гибридной комбинации. Закономерностью является лишь то, что как прямые, так и обратные гибриды F1 диких видов с S.melongena по основным морфологическим признакам и биометрическим показателям являются довольно схожими, что, предположительно, говорит о более сильной экспрессии генов диких видов в межвидовых гибридах.
Фертильность пыльцы межвидовых гибридов была на уровне 23 – 33%. Наиболее высокой она была у межвидового гибрида F1 S.macrocarpon x S.melongena (Л-Бриллиант) – 32,9%.
Таблица 9
Морфологические признаки различных видов и межвидовых гибридов баклажана рода Solanum L. (2009 – 2010 годы)
| №№ п/п | Вид, гибридная комбинация | Высота растения, см | Окраска цветка | Характеристики плода | ||
| Форма | Окраска в технической спелости | Окраска в биологической спелости | ||||
| 1 | S.melongena | 250-300 | сиреневый | цилиндрический | темно-фиолетовая | светло-фиолетовая |
| 2 | S.macrocarpon | 160-220 | темно-сиреневый | округлый | желтая | желтая |
| 3 | S.aethiopicum | 200-270 | белый | овальный | светло-красная | красная |
| 4 | S.macrocarpon x S.melongena (Л-Бриллиант) | 16 - 20 | темно-сиреневый | округлая, слегка удлиненная | желто-зеленая | желтая |
| 5 | S.melongena (Л-Бриллиант) х S.macrocarpon | 13 - 18 | темно-сиреневый | округлая, слегка удлиненная | светло-серая, лиловая | желтая |
| 6 | S.melongena (Л-Бриллиант) х S.aethiopicum | 180 - 190 | белый | округлая, овальная | желтая | оранжевая |
| 7 | S.aethiopicum х S.melongena (Л-Бриллиант) | 30 - 35 | белый | овальная, яйцевидная | желтая | оранжевая |
| 8 | S.aethiopicum х S.macrocarpon | 30 - 55 | светло-сирененвый | округлая | светло-оранжевая | оранжевая и красно-оранжевая |
Гибридность полученных в ходе межвидовых скрещиваний растений баклажана была также подтверждена с помощью ПЦР-анализа с использованием ISSR и IRAP праймеров. В спектрах амплифицированных ISSR- и IRAP-фрагментов индивидуальных растений F1 были выявлены как отцовские, так и материнские компоненты (рис. 9).
По результатам проведенного молекулярного анализа трех гибридных комбинаций была подтверждена гибридная природа всех индивидуальных растений F1, полученных с использованием метода эмбриокультуры in vitrо.
Биохимический анализ мякоти плодов межвидовых гибридов баклажана показал, что суммарное содержание флавоноидов и фенолкарбоновых кислот составило 2,6 - 2,7%, что в 1,5 раза выше по сравнению с родительским видом S.melongena (Л-Бриллиант). В межвидовых гибридах количество флавоноидов было в 1,3 раза выше по сравнению с родительским видом S.melongena (Л-Бриллиант) (табл. 10).
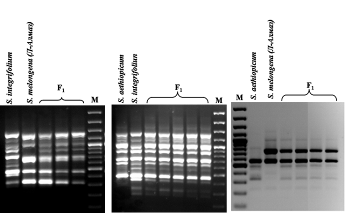
Рис 9. Электрофореграмма продуктов ПЦР-амплификации с помощью ISSR (А и Б) и IRAP (В) праймеров у трех родительских форм баклажана (S. melongena, S. aethiopicum и S. integrifolium) и их межвидовых гибридов F1. М – маркер молекулярных масс (GeneRuler, 100 bp DNA Ladder, "Fermentas", Литва).
Таблица 10
Содержание фенольных соединений в мякоти плодов межвидовых гибридов баклажана (2010 – 2011 годы)
| Линия, гибридная комбинация | Cодержание фенольных соединений,% | Содержание нерастворимых пектинов,% | |||
| Содержание антоцианов в кожуре плодов | Содержание ФКК в мякоти плодов | Содержание флавоноидов в мякоти плодов | Общее (4+5) | ||
| 2 | 3 | 4 | 5 | 6 | 7 |
| S.aethiopicum х S.melongena (Л-Бриллиант) | - | 1,7 ± 0,0 | 0,9 ± 0,0 | 2,6 | 0,57 |
| S.aethiopicum х S.melongena (Л-Алмаз) | - | 1,6 ± 0,0 | 1,0 ± 0,0 | 2,7 | 0,56 |
| S.melongena (Л-Бриллиант) x S.macrocarpon | - | 1,6 ± 0,0 | 1,1 ± 0,0 | 2,7 | 0,58 |
Содержание фенолкарбоновых кислот в межвидовых гибридах F1 составило 1,6 – 1,7%, что в 1,6-1,7 раза больше по сравнению с родительским видом S.melongena (Л-Бриллиант). По содержанию пектинов межвидовые гибриды незначительно отличались от исходных родительских видов и их содержание было промежуточным по отношению к ним.
Выводы
1. Показано, что раннеспелость гибридных комбинаций (80-85 суток) зависит от сочетаний высоких эффектов ОКС и СКС у родительских линий.
По количественным признакам, определяющим урожайность гибридных комбинаций F1 по проявлению эффекта гетерозиса, закономерности не наблюдается.
2. Основным антоцианом линий и гибридных комбинаций F1 селекции ВНИИССОК является дельфинидин-(6-кумароил)-3-глюкозид, который ранее не был выявлен в баклажане. Содержание антоцианов в плодах родительских линий составляет 0,12-0,18 г/кг плодов, в гибридных комбинациях – 0,14 – 0,31 г/кг. По данному признаку у 40% гибридных комбинаций F1 наблюдается эффект гетерозиса 105,6 - 193,8%.
3. Изучение морфогенетических реакций in vitro разных типов эксплантов показало, что наибольшей регенерационной активностью обладают семядольные экспланты, которые могут найти широкое применение для массового получения регенерантов баклажана на среде Мурасиге и Скуга с добавлением низких концентраций тидиазурона (0,02 - 0,2 мг/л ТДZ).
4. На основе изучения динамики развития зародышей видов S.melongena L., S.macrocarpon L., S.aethiopicum L. и их гибридов установлено, что зародыши проходят основные стадии развития (от глобулярного эмбриоида до полностью сформировавшегося зародыша) в промежутке 20 – 45 суток после опыления. Наиболее быстрое развитие зародыша наблюдается у вида S.aethiopicum – за 30 суток, у S.melongena – за 40 суток, у S.macrocarpon – за 45 суток. Скорость развития зародышей межвидовых гибридов незначительно отличается от скорости развития зародышей родительских видов.
5. Предложены элементы технологии получения межвидовых гибридов баклажана методом эмбриокультуры, состоящие из условий стерилизации семян, стадий развития зародышей, составов питательных сред для индукции, регенерации побегов, укоренения растений-регенерантов.
6. В межвидовых гибридных комбинациях баклажана в большей степени проявляются морфологические и биометрические признаки генеративных органов дикого вида. Показано, что прямые и обратные комбинации являются схожими по основным признакам (габитус растения, форма, размер и масса цветка и плода).
Фертильность пыльцы всех межвидовых гибридов, полученных при скрещивании S.melongena L., S.macrocarpon L. и S.aethiopicum L. не превышает 30-35%. Все плоды партенокарпические. Семена образуются только при опылении пыльцой культурного вида S. melongena.
7. Содержание флавоноидов в межвидовых гибридных комбинациях в 1,3 раза, фенолкарбоновых кислот – в 1,6-1,7 раза выше, чем у вида S.melongena.
Рекомендации к практическому использованию в селекции
1. Для получения перспективных гибридов F1, отличающихся высокой урожайностью, можно использовать линии Л-Багратион, Л-Бак и Л-Беглец, раннеспелостью – линию Л-Алмаз.
3. Для клонального микроразмножения селекционно ценных форм баклажана отечественных сортов использовать среду Мурасиге и Скуга с добавлением 0,02 и 0,2 мг/л ТДZ. Для регенерации побегов рекомендуется использовать среды с уменьшенной концентрацией тидиазурона (0,01 мг/л ТДZ и 0,01 мг/л ТДZ + 0,01 мг/л БАП). Для укоренения побегов, развившихся из почек каллуса семядолей и гипокотиля – среду Мурасиге и Скуга с половинным содержанием основных компонентов без добавления регуляторов роста.
4. Для получения межвидовых гибридов баклажана культивировать незрелые зародыши на стадии торпедовидного эмбриоида (> 2 мм) и более поздних (возраст > 30 суток с момента опыления) на среде Мурасиге и Скуга с добавлением 0,1 мг/л ТДZ. Срок развития зародышей на индукционной среде не должен превышать 7-14 суток. Для укоренения оптимальна среда Мурасиге и Скуга с половинным содержанием основных компонентов без добавления регуляторов роста.
5. Линии Л-Алмаз и Л-Солярис можно использовать как исходный материал для получения форм с высоким содержанием антоцианов в кожуре плодов баклажана.
6. Для получения форм с высоким содержанием фенольных соединений в мякоти плодов использовать межвидовые гибридные комбинации S.aethiopicum х S.melongena (Л-Бриллиант), S.aethiopicum х S.melongena (Л-Алмаз), S.melongena (Л-Бриллиант) x S.macrocarpon.
Список научных публикаций
1. Верба В.М. Антоцианы в плодах баклажана селекции ВНИИССОК/ М.И. Мамедов, О.Н. Пышная, Н.А. Шмыкова, В.М. Верба, Передеряев О.И.// Фенольные соединения: фундаментальные и прикладные аспекты. – М.: Научный мир, 2010. – с. 135-144.
2 Верба В.М. Клональное микроразмножение баклажана / В.М. Верба, М.И. Мамедов, О.Н. Пышная, Н.А. Шмыкова // Доклады ТСХА. – М: РГАУ-МСХА. – 2010. – Вып. 282. – Ч.1. – с. 603 – 607.
3. Верба В.М. Содержание антоцианов в плодах линий баклажана селекции ВНИИССОК в условиях малообъемной гидропоники / В.М. Верба, М.И. Мамедов, О.Н. Пышная, Н.А. Шмыкова // Овощи России. – 2009. – № 3. – с.37-42.
4. Верба В.М. Культурный баклажан – происхождение, задачи селекции и генетические источники / М.И. Мамедов, О.Н. Пышная, В.М. Верба // Овощи России. – 2009. – № 4 (6). – 2010. – № 1 (7). – с.27 – 33.
5. Верба В.М. Получение межвидовых гибридов баклажана методом эмбриокультуры / В.М. Верба, М.И. Мамедов, О.Н. Пышная, Н.А. Шмыкова // Сельскохозяйственная биология. – 2010. – №5. – с.66-71.
6. Верба В.М. Клональное микроразмножение баклажана путем органогенеза / В.М. Верба, М.И. Мамедов, О.Н. Пышная, Н.А. Шмыкова // Вестник РАСХН. – 2010. – № 6. – с.57-59.
7. Верба В.М. Определение количественного состава антоцианов в плодах баклажана (S. melongena) селекции ВНИИССОК / В.М. Верба, М.И. Мамедов, О.Н. Пышная, Н.А. Шмыкова // Доклады ТСХА. - М: РГАУ-МСХА, 2011 (в печати).