

Популяционные особенности ядовитого секрета обыкновенной гадюки vipera berus ( linnaeus, 1758) в волжском бассейне
На правах рукописи
ЗАЙЦЕВА ОЛЬГА ВЛАДИМИРОВНА
ПОПУЛЯЦИОННЫЕ ОСОБЕННОСТИ
ЯДОВИТОГО СЕКРЕТА ОБЫКНОВЕННОЙ ГАДЮКИ
Vipera berus (Linnaeus, 1758) В ВОЛЖСКОМ БАССЕЙНЕ
Специальность 03.02.08 – экология (биология)
Автореферат
диссертации на соискание ученой степени
кандидата биологических наук
Тольятти – 2011
Работа выполнена в лаборатории герпетологии и токсинологии
Института экологии Волжского бассейна РАН
Научный руководитель: кандидат биологических наук
Маленев Андрей Львович
Официальные оппоненты: доктор биологических наук, профессор
Гелашвили Давид Бежанович;
доктор биологических наук
Розенцвет Ольга Анатольевна
Ведущая организация: Самарский государственный университет
Защита диссертации состоится 25 ноября 2011 г. в 1000 часов на заседании диссертационного совета Д 002.251.01 при Институте экологии Волжского бассейна РАН по адресу: 445003, Самарская обл., г. Тольятти, ул. Комзина, 10.
Тел. 8 (8482) 489977; факс 8 (8482) 489504; E-mail: [email protected]
С диссертацией можно ознакомиться в библиотеке Института экологии Волжского бассейна РАН, с авторефератом – в сети Интернет на сайте ИЭВБ РАН по адресу: http://www.ievbras.ru
Автореферат разослан «____» октября 2011 г.
Ученый секретарь
![]()
диссертационного совета,
кандидат биологических наук А.Л. Маленёв
Общая характеристика работы
Актуальность темы исследований. Гадюки, обитающие в популяциях Волжского бассейна, являются неотъемлемым компонентом экосистем и одним из звеньев трофической цепи. Обыкновенную гадюку Vipera berus (Linnaeus, 1758) можно рассматривать и как хозяйственно ценный вид – продуцент ядовитого секрета, который служит сырьем при производстве ряда лекарственных препаратов и сыворотки «Антигадюка». Разработка и производство новых лекарств на основе ядов гадюк невозможны без учета биохимических свойств сырья – ядовитого секрета. Для создания более эффективных сывороток против укуса гадюк необходимо учитывать ряд внутривидовых и межпопуляционных особенностей белкового состава яда.
В Волжском бассейне распространены два подвида обыкновенной гадюки – номинативный V. b. berus (Linnaeus, 1758) и лесостепной, или гадюка Никольского V. b. nikolskii Vedmederja, Grubant et Rudaeva, 1986. При этом значительную часть Волжского бассейна охватывает зона интерградации этих подвидов (Зиненко, 2003; Milto, Zinenko, 2005; Бакиев и др., 2008, 2009). Ядовитый секрет гадюк в поволжских популяциях имеет свои биохимические особенности (различающиеся активности ферментов и пептидный состав), которые также можно использовать в качестве дополнительных критериев для таксономической дифференциации вышеуказанных подвидов гадюк. Все вышеперечисленные причины и определяют актуальность популяционных исследований свойств ядовитого секрета обыкновенных гадюк в Волжском бассейне.
Цель исследования: анализ подвидовых, меж- и внутрипопуляционных особенностей ядовитого секрета обыкновенных гадюк, встречающихся на территории Волжского бассейна. Для достижения поставленной цели необходимо было решить следующие задачи:
1) определить токсичность (среднесмертельную дозу ЛД50) ядовитого секрета обыкновенных гадюк из разных популяций на территории Волжского бассейна;
2) проанализировать различия в активности протеолитических ферментов и оксидазы L-аминокислот яда обыкновенных гадюк из различных точек ареала в пределах Волжского бассейна;
3) выявить подвидовые и межпопуляционные особенности пептидного состава ядовитого секрета гадюк в бассейне Волги;
4) определить различия активности оксидазы L-аминокислот и пептидного состава в образцах яда разных цветовых вариаций;
5) установить направленность изменений свойств ядовитого секрета обыкновенных гадюк в популяциях на территории Волжского бассейна.
Научная новизна. Впервые определены значения ЛД50 яда обыкновенных гадюк из ряда поволжских популяций. Впервые выявлены и проанализированы меж- и внутрипопуляционные различия активности ферментов яда гадюк в Волжском бассейне и отмечены особенности пептидного состава яда, обусловленные подвидовой принадлежностью гадюк. Охарактеризованы различия активности оксидазы L-аминокислот и пептидного состава в образцах яда разного цвета. Впервые отмечена тенденция изменений свойств яда гадюк, направленная от популяций с признаками номинативного подвида к популяциям с признаками гадюки Никольского, т.е. от Волжского бассейна к Донскому.
Теоретическое значение результатов. Результаты исследований, изложенные в диссертации, дополняют сведения о популяционной изменчивости свойств змеиных ядов, расширяют диапазон диагностических признаков в таксономии гадюк и вносят вклад в популяционную экологию и прикладную герпетологию.
Практическая значимость результатов. Результаты исследования внутривидовой изменчивости свойств яда обыкновенной гадюки могут быть использованы при разработке современной нормативно-технической документации на змеиные яды, при производстве более эффективной сыворотки против укусов гадюк на территории Российской Федерации, а также при решении спорных вопросов подвидовой принадлежности обыкновенных гадюк.
Связь темы диссертации с плановыми исследованиями. Исследования по теме входят в план фундаментальных исследований ИЭВБ по направлениям «Экология организмов и сообществ» (6.2. «Закономерности природной и антропогенной трансформации экосистем различного типа и уровня организации бассейна Средней и Нижней Волги») и «Биологическое разнообразие» (6.3. «Видовое и экосистемное биоразнообразие в бассейне Средней и Нижней Волги»).
Апробация работы. Результаты диссертационного исследования докладывались и обсуждались на научных конференциях «Татищевские чтения: актуальные проблемы науки и практики» (Тольятти, 2007, 2010), Четвертой конференции герпетологов Поволжья (Тольятти, 2007), Второй международной научной конференции «Актуальные проблемы экологической физиологии, биохимии и генетики животных» (Саранск, 2009), Чтениях памяти проф. В.А. Попова (Казань, 2009), Молодежной научной конференции «Актуальные проблемы экологии Волжского бассейна» (Тольятти, 2009, 2011), Международной молодежной конференции (Уфа, 2009), IV съезде Герпетологического общества им. А.М. Никольского (Казань, 2009).
Публикация результатов исследования. По теме диссертации автором опубликовано 19 печатных работ, в том числе 5 публикаций в ведущих рецензируемых изданиях, рекомендованных ВАК, и одна коллективная монография.
Декларация личного участия автора. Автор лично принимал участие в полевых исследованиях, провел большую часть лабораторных экспериментов и статистическую обработку результатов. Текст диссертации написан автором по плану, согласованному с научным руководителем. Доля участия автора в совместных публикациях пропорциональна числу соавторов.
Основные положения, выносимые на защиту.
1. Токсичность (среднесмертельная доза ЛД50) ядовитого секрета обыкновенных гадюк в Волжском бассейне не зависит от географического местоположения популяции и половой принадлежности гадюк.
2. В активности ферментов яда обыкновенных гадюк из поволжских популяций имеются внутривидовые, меж- и внутрипопуляционные различия.
3. Два подвида обыкновенных гадюк в Волжском бассейне различаются пептидным составом ядовитого секрета, активностью ферментов нем и, как правило, цветом яда.
4. Изменения свойств ядовитого секрета (снижение активности ферментов и интенсивности желтой окраски, изменение пептидного состава) в пределах Волжского бассейна направлены от популяций с преобладающими признаками номинативного подвида V. b. berus к популяциям с превалирующими признаками гадюки Никольского V. b. nikolskii, т.е. от Волжского бассейна к Донскому.
Объем и структура диссертации. Диссертация состоит из введения, 7 глав, выводов и списка использованной литературы. Общий объем диссертации составляет 107 страниц. Работа содержит 10 таблиц и 16 рисунков. Список литературы включает 209 источников, в том числе 86 на иностранных языках.
Благодарности. Автор благодарит за помощь в сборе и обработке материалов: н.с. ИЭВБ РАН, к.б.н. Г.В. Епланову, аспиранта ИЭВБ РАН И.В. Шуршину, с.н.с. ИЭВБ РАН, д.б.н. В.К. Шитикова, в.н.с. Музея природы Харьковского национального университета им. В.Н. Каразина (г. Харьков, Украина), к.б.н. А.И. Зиненко. Особую признательность автор выражает научному руководителю – заведующему лабораторией герпетологии и токсинологии ИЭВБ РАН, к.б.н. А.Л. Маленеву и с.н.с. ИЭВБ РАН, к.б.н., доценту А.Г. Бакиеву за внимание и помощь на всех этапах работы.
ОСНОВНОЕ Содержание работы
Глава 1. Обзор литературы
В главе приведены современные литературные данные по систематике, биологии и экологии обыкновенной гадюки Vipera berus на территории Волжского бассейна, а также сведения о составе и свойствах ее ядовитого секрета.
Глава 2. Материалы и методы
Обыкновенных гадюк отлавливали во время экспедиций с апреля по сентябрь 2005–2011 гг. в различных областях Волжского бассейна и содержали в лаборатории герпетологии и токсинологии ИЭВБ РАН. После отбора яда и морфологических измерений змей по возможности выпускали в места отлова. Точки отлова гадюк приведены на рис. 1.
Отбор яда проводили механическим способом в чашки Петри, массируя пальцами ядовитые железы. Образцы высушивали до постоянной массы в эксикаторе над хлористым кальцием в течение 14–15 суток при комнатной температуре и хранили при температуре +5°С в полипропиленовых микропробирках. Каждый образец представлял собой кристаллизованный секрет ядовитых желез, полученный за одно ядовзятие от одной гадюки («индивидуальный» образец) или от нескольких гадюк из одной популяции («объединенный» образец). Ряд образцов ядовитого секрета гадюк был любезно предоставлен коллегами – В.Г. Старковым, А.И. Зиненко, Н.А. Литвиновым, Н.А. Наумкиной, М.В. Ушаковым, Д.Н. Ползиковым.
Токсичность яда обыкновенной гадюки. Эксперименты по токсикометрии яда обыкновенной гадюки проводили на белых мышах-самцах массой 20±1 г. Инъекции растворов яда мышам проводили подкожно, и через 24 часа фиксировали количество умерших и выживших экспериментальных животных. Среднесмертельную дозу яда (ЛД50) рассчитывали методом модифицированного пробит-анализа (Безруков и др., 1995). Для определения ЛД50 использовали «объединенные» образцы яда обыкновенных гадюк, обитающих в разных популяциях Волжского бассейна.
Протеолитическую активность в яде обыкновенной гадюки устанавливали по гидролизу казеината натрия (Murata et al., 1963), поэтому в дальнейшем речь пойдет о казеинолитической составляющей протеаз.
Активность оксидазы L-аминокислот измеряли спектрофотометрически с использованием в качестве субстрата L-фенилаланина (Wellner, Liсhtenberg, 1971).
Концентрацию белка в растворах ядовитого секрета определяли по методу Лоури (Lowry et al., 1951).
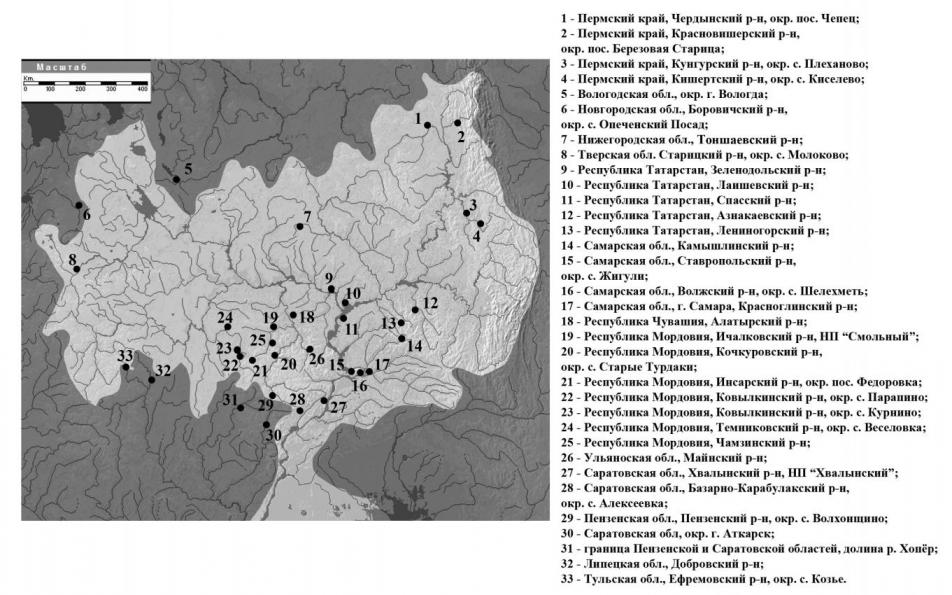
Рис. 1. Места отлова гадюк
Обозначения: 1 – Пермский край, Чердынский р-н, окр. пос. Чепец; 2 – Пермский край, Красновишерский р-н, окр. пос. Березовая Старица; 3 – Пермский край, Кунгурский р-н, окр. с. Плеханово; 4 – Пермский край, Кишертский р-н, окр. с. Киселёво; 5 – Вологодская обл., окр. г. Вологда; 6 – Новгородская обл., Боровичский р-н, окр. с. Опеченский Посад; 7 – Нижегородская обл., Тоншаевский р-н; 8 – Тверская обл., Старицкий р-н, окр. с. Молоково; 9 – Республика Татарстан, Зеленодольский р-н; 10 – Республика Татарстан, Лаишевский р-н; 11 – Республика Татарстан, Спасский р-н; 12 – Республика Татарстан, Азнакаевский р-н; 13 – Республика Татарстан, Лениногорский р-н; 14 – Самарская обл., Камышлинский р-н, окр. с. Татарский Байтуган; 15 – Самарская обл., Ставропольский р-н, окр. с. Жигули; 16 – Самарская обл., Волжский р-н, окр. с. Шелехметь; 17 – Самарская обл., г. Самара, Красноглинский р-н, ур. Горелый Хутор; 18 – Республика Чувашия, Алатырский р-н; 19 – Республика Мордовия, Ичалковский р-н, НП «Смольный»; 20 – Республика Мордовия, Кочкуровский р-н, окр. с. Старые Турдаки; 21 – Республика Мордовия, Инсарский р-н, окр. пос. Федоровка; 22 – Республика Мордовия, Ковылкинский р-н, окр. с. Парапино; 23 – Республика Мордовия, Ковылкинский р-н, окр. с. Курнино; 24 – Республика Мордовия, Темниковский р-н, окр. с. Веселовка; 25 – Республика Мордовия, Чамзинский р-н; 26 – Ульяновская обл., Майнский р-н; 27 – Саратовская обл., Хвалынский р-н, НП «Хвалынский»; 28 – Саратовская обл., Базарно-Карабулакский р-н, окр. с. Алексеевка; 29 – Пензенская обл., Пензенский р-н, окр. с. Волхонщино; 30 – Саратовская обл., Аткарский р-н, окр. г. Аткарск; 31 – граница Пензенской и Саратовской областей, долина р. Хопёр; 32 – Липецкая обл., Добровский р-н; 33 – Тульская обл., Ефремовский р-н, окр. с. Козье.
Электрофорез образцов ядовитого секрета. Электрофоретическое разделение пептидов яда проводили по методу Лэммли (Laemmli, 1970) в вертикальных пластинах полиакриламидного геля (толщина пластины – 0,75 мм, размер – 80140 мм) с использованием 0,025 М Трис-глицинового электродного буфера (рН = 8,3) в присутствии 0,1% SDS и 3 мМ ЭДТА. Концентрация акриламида в разделяющем геле составляла 12,5%, в концентрирующем – 5%. Определение молекулярной массы исследуемых пептидов и расчет их процентного содержания в образце осуществляли с помощью программы Gel Pro Analyzer.
Морфология. У обыкновенных гадюк отмечали следующие признаки фолидоза: количество брюшных щитков, не считая анального (Ventr.), количество пар подхвостовых (S.cd.) щитков, количество рядов чешуй вокруг середины тела (Sq.), количество верхнегубных (Lab.) и нижнегубных (S.lab.) щитков, количество чешуй вокруг глаза, не считая надглазничного (C.oc.), количество мелких щитков между глазом и верхнегубными щитками (S.oc.), количество скуловых щитков (Lor.), количество интеркантальных чешуй (Ic.), число парафронтальных щитков (Pf.). В работе использовали не только живых змей, но и экземпляры обыкновенных гадюк из коллекции рептилий Института экологии Волжского бассейна РАН.
Объем исследованного материала. Для морфологических исследований использовано более 250 экземпляров обыкновенной гадюки, проанализировано на активности ферментов более 50 «объединенных» и более 250 «индивидуальных» образцов ядовитого секрета, проведено электрофоретическое разделение в ПААГ более чем 150 образцов.
Статистическую обработку экспериментальных данных проводили с помощью общепринятых методов биометрии (Лакин, 1980) и пакета прикладных программ Excel, PAST и РС-ORD.
Глава 3. Токсичность ядовитого секрета
В экспериментах по токсикометрии образцы ядовитого секрета подобраны таким образом, чтобы можно было сравнить значения яда гадюк из географически удаленных друг от друга популяций и ЛД50 яда гадюк различной подвидовой принадлежности (табл. 1). При попарном сравнении ЛД50 образцов яда гадюк из разных местообитаний статистически значимых межпопуляционных различий не выявлено (P>0,05), даже при сравнении крайних значений ЛД50: минимального – 2,9±0,52 мг/кг и максимального – 4,7±1,09 мг/кг.
Большая часть образцов яда была получена от гадюк с морфологическими признаками номинативного подвида, а образцы яда из Пензенской, Саратовской областей и районов на их границе получены от гадюк, по морфологическим признакам определенным как гадюка Никольского. Сравнение значений ЛД50 образцов яда гадюк разных подвидов не выявило статистически значимых подвидовых различий в токсичности ядовитого секрета обыкновенных гадюк.
На примере гадюк, обитающих в черте г. Самара, статистически значимых половых различий в токсичности ядовитого секрета нам также обнаружить не удалось, что говорит в пользу отсутствия у самок или самцов каких-либо преимуществ в умерщвлении потенциальных пищевых объектов – мышевидных грызунов.
Таблица 1
ЛД50 образцов яда обыкновенных гадюк из различных точек ареала
| Область сбора образцов | ЛД50 (мг/кг) М±m | |
| Самарская | самцы | 3,9±0,52 |
| самки | 3,7±0,50 | |
| Нижегородская | 3,2±0,36 | |
| Ульяновская | 3,9±0,65 | |
| Республика Татарстан | 3,6±0,66 | |
| Республика Мордовия | 3,0±0,67 | |
| Республика Чувашия | 4,7±1,09 | |
| Пермская | 3,2±0,72 | |
| Пензенская* | 4,1±0,73 | |
| Граница Пензенской и Саратовской областей* | 2,9±0,52 | |
| Саратовская* | 3,6±0,51 | |
| Харьковская* | 3,0±0,60 | |
Примечание: * – образцы ядовитого секрета обыкновенных гадюк, по внешним морфологическим признакам относящихся к подвиду V. b. nikolskii.
Глава 4. Активности ферментов ядовитого секрета
4.1. Межпопуляционные и подвидовые различия
Выбор ферментов обусловлен тем, что протеолитические ферменты являются определяющим звеном в картине отравления ядом гадюк (Орлов и др., 1990; Gutierrez et al., 2010), а оксидаза L-аминокислот, действуя совместно с протеазами, определяет цвет ядовитого секрета (Iwanaga, Suzuki, 1979; Tan, Fung, 2010).
Эксперименты по определению межпопуляционных различий в активности ферментов яда гадюк были проведены на «объединенных» образцах ядовитого секрета, где собран ядовитый секрет от гадюк разного размера, возраста, пола, но из одной популяции. Здесь параметры ядовитого секрета усреднены уже во время ядовзятия, и отражают лишь общий уровень активности того или иного фермента яда гадюк в популяции. При сравнении активности ферментов яда гадюк из разных популяций можно говорить лишь о межпопуляционных различиях без учета многих других факторов, могущих влиять на активность ферментов ядовитого секрета.
Усредненные значения удельной активности ферментов ядовитого секрета обыкновенных гадюк, встречающихся в поволжских популяциях, полученные при анализе «объединенных» образцов яда, представлены в табл. 2.
Таблица 2
Активности ферментов в «объединенных» образцах
ядовитого секрета обыкновенных гадюк
| Район отлова | Координаты точки отлова | Протеаза, мкг тирозина/мг белка мин | Оксидаза L-аминокислот, Е/мг белка мин | ||||
| n | M±m | lim | n | M±m | lim | ||
| Нижегородская обл., Тоншаевский р-н | 5783.607’ с.ш. 4733.769’ в.д. | 9 | 18,6±0,48 | 16,4–20,7 | 6 | 25,5±2,01 | 20,6–34,6 |
| Респ. Татарстан, Зеленодольский р-н | 5590.419’ с.ш. 4879.611’ в.д. | 6 | 15,6±0,46 | 14,3–17,2 | 11 | 25,2±0,73 | 21,3–29,3 |
| Респ. Чувашия Алатырский р-н | 5501.831’с.ш. 4649.142’ в.д. | 6 | 22,4±0,64 | 19,7–23,9 | 3 | 25,3±0,33 | 24,7–25,8 |
| Респ. Татарстан, Лениногорский р-н | 5472.918’ с.ш. 5229.354’ в.д. | 11 | 14,2±0,56 | 11,3–17,9 | 17 | 21,9±0,66 | 17,0–26,7 |
| Респ. Мордовия, Ичалковский р-н, НП «Смольный» | 5450.110’ с.ш. 4522.641’ в.д. | 9 | 13,3±0,55 | 6,7–19,7 | 3 | 20,6±0,29 | 20,9–20,0 |
| Респ. Мордовия, Темниковский р-н, окр. с. Веселовка | 5433.009’ с.ш. 4258.579’ в.д. | 5 | 17,9±1,56 | 14,5–23,4 | 3 | 27,1±1,60 | 24,9–30,3 |
| Ульяновская обл., Майнский р-н | 5405.319’ с.ш. 4773.937’ в.д. | 6 | 25,8±1,44 | 19,6–31,3 | 3 | 25,6±1,48 | 23,7–28,5 |
| Респ. Мордовия, Ковылкинский р-н, окр. с. Парапино | 5359.278’ с.ш. 4357.059’ в.д. | 6 | 16,6±0,76 | 14,4–18,9 | 3 | 25,8±1,10 | 23,6–27,0 |
| г. Самара, Красноглинский р-н, Горелый Хутор | 5322.842’ с.ш. 4918.552’ в.д. | 15 | 20,2±0,85 | 14,6–26,1 | 6 | 20,9±0,69 | 18,5–23,5 |
| Саратовская обл., Хвалынский р-н, НП «Хвалынский» | 5248.789’ с.ш. 4804.766’ в.д. | 5 | 8,9±1,52 | 4,9–14,3 | 3 | 6,4±0,64 | 5,2–7,5 |
| Граница Пензенской и Саратовской обл., пойма р. Хопёр* | 5237.429’ с.ш. 4347.633’ в.д. | 5 | 8,2±059 | 6,1–9,4 | 3 | 1,4±0,21 | 1,0–1,8 |
| Саратовская обл., Аткарский р-н, пой-ма р. Медведица* | 5150.280’ с.ш. 4459.381’ в.д. | 5 | 9,21±1,44 | 7,4–15,0 | 3 | 0,9±0,06 | 0,9–1,0 |
| Украина, Харьковская обл., Змиевский р-н * | 4960.703’ с.ш. 3633.796’ в.д. | 3 | 2,6±0,49 | 2,1–3,0 | 3 | 5,7±0,77 | 4,1–6,6 |
Примечания: * – образцы ядовитого секрета обыкновенных гадюк, по внешним морфологическим признакам относящихся к подвиду V. b. nikolskii;
n – количество определений активности ферментов в «объединенном» образце.
Анализ экспериментальных данных позволяет сделать некоторые выводы. Во-первых, средние величины активности протеаз и оксидазы L-аминокислот яда гадюк в разных популяциях Волжского бассейна различны. Так, значения активности протеолитических ферментов в яде изменяются от максимальных значений в Ульяновской области (25,8±1,44) до минимальных, наблюдаемых в популяции на границе Пензенской и Саратовской областей (8,2±059).
Максимальная удельная активность оксидазы L-аминокислот обнаружена нами в популяции гадюк из Республики Мордовия (27,1±1,60), минимальная – отмечена в популяции из Саратовской области (0,9±0,06). При этом различия средних значений оказались в высшей степени статистически значимыми и для протеолитических ферментов (Р<0,001), и для оксидазы L-аминокислот (Р<0,001). Надо подчеркнуть, что максимальные значения активности обоих ферментов отмечены в популяциях гадюк номинативного подвида, а минимальные – в популяциях гадюки Никольского. При попарном сравнении средних значений активности ферментов яда в популяциях гадюк одного подвида в ряде случаев также наблюдаются статистически значимые различия. Например, при сравнении протеолитической активности ядов гадюк из популяции Ульяновской области и Татарстана (Р<0,001) или оксидазы L-аминокислот яда гадюк, встречающихся в Самарской области и Республике Мордовия (Р<0,01).
Во-вторых, в яде гадюк с хорошо выраженными признаками гадюки Никольского V. b. nikolskii (популяции из Саратовской области и на ее границе с Пензенской) обнаруживается более низкая, чем в яде гадюк номинативного подвида V. b. berus, активность ферментов: протеаз – в 3–8 раз, оксидазы L-аминокислот – в 4–20 раз. При попарном сравнении активности ферментов яда из любой популяции гадюк номинативного подвида V. b. berus с таковой яда гадюк Никольского V. b. nikolskii различия между средними значениями в яде гадюк разных подвидов также оказываются статистически значимыми.
В-третьих, анализ изменений уровня активности ферментов показывает общую тенденцию уменьшения средних значений активностей исследуемых ферментов, направленную от популяций с превалирующими признаками номинативного подвида V. b. berus в северных и центральных районах Волжского бассейна к популяциям с преобладающими признаками гадюки Никольского V. b. nikolskii на границе бассейнов Волги и Дона.
4.2. Активность оксидазы L-аминокислот и цвет ядовитого секрета
Из литературных данных известно, что желтый цвет яда обусловлен содержанием оксидазы L-аминокислот и наличием кофермента ФАД (Iwanaga, Suzuki, 1977). Считается также, что для номинативного подвида обыкновенной гадюки более характерен желтый яд, а для гадюки Никольского – бесцветный (Milto, Zinenko, 2005). В ходе исследований мы обнаружили, что обыкновенные гадюки в популяциях Волжского бассейна продуцируют ядовитый секрет разной окраски: ярко-желтый, желтоватый и бесцветный. Кроме того, нами была обнаружена популяция в Пензенском районе Пензенской области, где одни особи продуцируют бесцветный, другие – бледно-желтый ядовитый секрет. Мы определили активность этого фермента в образцах яда обыкновенной гадюки, имеющих разный цвет, из ряда поволжских популяций (табл. 3).
Таблица 3
Активность оксидазы L-аминокислот в образцах яда разного цвета
| Место отлова | Цвет яда | n | Удельная активность оксидазы L-аминокислот, Е/мг белка мин | |
| M±m | lim | |||
| Республика Мордовия, Ковылкинский р-н | ярко-желтый | 3 | 25,8±1,10 | 23,6–27,0 |
| Нижегородская обл., Тоншаевский р-н | ярко-желтый | 6 | 25,5±2,01 | 20,6–34,6 |
| Республика Татарстан, Лениногорский р-н | ярко-желтый | 17 | 21,9±0,66 | 17,0–26,7 |
| Самарская обл., НП «Самарская Лука» | ярко-желтый | 7 | 20,9±0,69 | 18,45–23,45 |
| Саратовская обл., Хвалынский р-н | желтоватый | 3 | 6,4±0,64 | 5,2–7,5 |
| Пензенская обл., Пензенский р-н* | желтоватый | 5 | 6,1±0,14 | 5,36–6,98 |
| бесцветный | 6 | 0,5±0,14 | 0,02–0,85 | |
| Граница Пензенской и Саратовской обл.* | желтоватый | 3 | 1,4±0,21 | 1,0–1,8 |
| Саратовская обл., Аткарский р-н* | бесцветный | 3 | 0,9±0,06 | 0,9–1,0 |
Примечания: * – образцы ядовитого секрета обыкновенных гадюк, по внешним морфологическим признакам относящихся к подвиду V. b. nikolskii;
n – количество определений активности ферментов в «объединенном» образце.
Активность оксидазы L-аминокислот меняется в образцах от средних значений менее 1,0 Е/мг белка в мин (бесцветный яд гадюк из Саратовской и Пензенской областей) до 25,8 единиц (ярко-желтый яд гадюк из Республики Мордовия). При попарном сравнении средних значений активности фермента в образцах, относящихся к разным популяциям, обнаруживаются статистически значимые различия. К примеру, различия между выборочными средними образцов яда гадюк из Самарской и Пензенской областей в высшей степени достоверны (Р<0,001). В то же время, сравнение между собой образцов ядовитого секрета желтого цвета из популяций номинативного подвида (Самарская и Нижегородская области и Республики Мордовия и Татарстан) статистически значимых различий не выявило (Р>0,05). Можно заметить, что образцы яда гадюк из популяций, морфологически близких к гадюке Никольского, были либо бесцветными, либо бледно-желтого цвета. Активность оксидазы L-аминокислот в этих образцах значительно ниже таковой в яде гадюк номинативного подвида – при сравнении средних значений здесь также наблюдаются статистически значимые подвидовые различия. Кроме того, яд светло-желтой окраски с промежуточными значениями активности оксидазы L-аминокислот характерен для гадюк, популяции которых относятся к зоне интерградации, где у гадюк отмечаются признаки обоих подвидов.
4.3. Внутрипопуляционная изменчивость ферментов яда
Анализ активности ферментов ядовитого секрета гадюк с использованием «индивидуальных» образцов яда позволяет выявить особенности ядовитого секрета особей внутри популяций. А отлов гадюк в одном месте и в одно время позволяет исключить географический, сезонный и микроклиматический факторы, которые могут влиять на значения активности ферментов в яде. Для анализа использовали данные из семи популяций обыкновенных гадюк (табл. 4).
Таблица 4
Активности ферментов в «индивидуальных» образцах
ядовитого секрета обыкновенных гадюк
| Место отлова | Пол | n | Протеаза, мкг тирозина/мг белка в мин | n | Оксидаза L-аминокислот, Е/мг белка в мин | ||
| M±m | lim | M±m | lim | ||||
| Пермский край, Чердынский р-н, окр. пос. Чепец | самцы | 8 | 21,4±2,35 | 11,5–32,8 | 8 | 24,6±4,60 | 10,1–47,3 |
| самки | 32 | 18,5±1,05 | 9,8–34,0 | 32 | 24,5±1,37 | 12,2–45,0 | |
| Вологодская обл., окр. г. Вологда | самцы | 4 | 22,5±3,92 | 12,5–31,1 | 4 | 18,5±2,19 | 12,0–21,7 |
| самки | 5 | 21,2±3,15 | 14,6–32,2 | 5 | 25,1±2,05 | 16,9–27,6 | |
| г. Самара, Красноглинский р-н, ур. Горелый Хутор | самцы | 29 | 20,1±1,42 | 8,4–37,9 | 11 | 21,4±2,69 | 1,2–32,0 |
| самки | 15 | 20,6±1,97 | 9,8–34,6 | 11 | 26,2±2,23 | 16,2–38,1 | |
| Самарская обл., Волжский р-н, окр. с. Шелехметь | самцы | 18 | 15,4±1,47 | 8,3–28,9 | 18 | 24,6±1,64 | 10,5–35,1 |
| самки | 6 | 17,1±4,21 | 2,5–32,9 | 6 | 20,4±3,07 | 13,7–34,6 | |
| Самарская обл., Ставропольский р-н, окр. с. Жигули | самцы | 21 | 10,7±0,67 | 4,3–14,8 | 17 | 19,3±1,21 | 11,7–28,8 |
| самки | 9 | 12,9±1,38 | 8,1–22,8 | 9 | 24,4±1,75 | 18,3–34,2 | |
| Пензенская обл., Пензенский р-н, окр. с. Волхонщино* | самцы | 26 | 9,8±0,49 | 4,9–15,3 | 26 | 3,4±0,88 | 0–13,9 |
| самки | 17 | 8,8±0,46 | 6,4–12,2 | 17 | 1,4±0,70 | 0–9,7 | |
| Липецкая обл., Добровский р-н* | самцы | 5 | 5,6±0,39 | 4,6–6,6 | 5 | 0,8±0,66 | 0–3,4 |
| самки | 10 | 6,5±0,44 | 4,1–8,4 | 10 | 0,8±0,32 | 0–3,5 | |
Примечания: * – образцы ядовитого секрета обыкновенных гадюк, по внешним морфологическим признакам относящихся к подвиду V. b. nikolskii;
n – количество проанализированных «индивидуальных « образцов.
Во всех исследованных популяциях обыкновенной гадюки статистически значимых половых различий в ферментативной активности яда не обнаружено, что говорит в пользу равных условий у самцов и самок в умерщвлении добычи и переваривании пищи.
В яде обыкновенных гадюк из исследованных популяций наблюдается высокая внутрипопуляционная изменчивость активности ферментов. Минимальные и максимальные значения активности протеаз внутри каждой популяции различаются в 3 и более раз. Еще большую изменчивость проявляет оксидаза L-аминокислот. Это обусловлено наличием в выборке образцов «бесцветного» яда с нулевыми значениями активности оксидазы L-аминокислот, и, вероятно, объясняется большей генетической гетерогенностью особей, входящих в состав этих популяций. Основной причиной различий свойств ядовитого секрета гадюк внутри отдельных популяций является, по мнению ряда исследователей, различный генетический статус особей. Считается, что изменчивость свойств ядовитого секрета находится под генетическим контролем и служит основой микроэволюционного процесса (Chippaux et al., 1991; Mebs, 1999).
Глава 5. Пептидный состав ядовитого секрета
5.1. Подвидовые особенности пептидного состава яда
Для определения подвидовых различий пептидного состава ядовитого секрета гадюк из популяций Волжского бассейна «объединенные» образцы яда были проанализированы методом электрофореза в ПААГ. На рис. 2 и 3 представлены типичные картины распределения основных пептидных компонентов в яде гадюк двух подвидов. Пептидный состав яда гадюк номинативного подвида V. b. berus с большой территории Волжского бассейна однороден, и образцы имеют лишь незначительные различия в количествах некоторых пептидных компонентов (рис. 2, образцы 1 – 8).
Сравнение между собой образцов яда V. b. nikolskii из разных популяций также говорит о схожести их пептидного состава (рис. 3: образцы 3–9). Но между ядовитым секретом V. b. berus и V. b. nikolskii видны качественные различия: это касается пары низкомолекулярных белков массой 16,8 и 14,7 кДа, которые в образцах V. b. nikolskii имеют другую массу. Для ядовитого секрета гадюк номинативного подвида характерен пептид массой 66 кДа, который не отмечен в яде гадюки Никольского. Минорный компонент с массой 19,8 кДа обнаружен лишь в образцах яда гадюк из Саратовской и Харьковской областей, т.е. у гадюки Никольского.
 Рис. 2. Электрофореграмма ядовитого секрета V. b. berus из разных популяций Обозначения: 1 – Ульяновская область, 2 – Республика Чувашия, 3 – Нижегородская область, 4 – Самарская область, 5 – Республика Татарстан, 6 – Республика Мордовия, 7 – Киевская область, 8 – Черниговская область, 9* – Харьковская область, 10* – Хвалынский р-н Саратовской области, * – приведенные для сравнения образцы яда V. b. nikolskii. Рис. 2. Электрофореграмма ядовитого секрета V. b. berus из разных популяций Обозначения: 1 – Ульяновская область, 2 – Республика Чувашия, 3 – Нижегородская область, 4 – Самарская область, 5 – Республика Татарстан, 6 – Республика Мордовия, 7 – Киевская область, 8 – Черниговская область, 9* – Харьковская область, 10* – Хвалынский р-н Саратовской области, * – приведенные для сравнения образцы яда V. b. nikolskii. |  Рис. 3. Электрофореграмма ядовитого секрета V. b. nikolskii из разных популяций Обозначения: 1* – Нижегородская область; 2* – Самарская область; 3 – Пензенская область (желтый яд); 4 – Пензенская область (бесцветный яд); 5 – граница Пензенской и Саратовской обл.; 6 – Аткарский р-н Саратовской области; 7 – Кировоградская область; 8 – Харьковская область; 9 – Хвалынский р-н Саратовской области, * – приведенные для сравнения образцы яда V. b. berus. Рис. 3. Электрофореграмма ядовитого секрета V. b. nikolskii из разных популяций Обозначения: 1* – Нижегородская область; 2* – Самарская область; 3 – Пензенская область (желтый яд); 4 – Пензенская область (бесцветный яд); 5 – граница Пензенской и Саратовской обл.; 6 – Аткарский р-н Саратовской области; 7 – Кировоградская область; 8 – Харьковская область; 9 – Хвалынский р-н Саратовской области, * – приведенные для сравнения образцы яда V. b. berus. |
| В правой колонке указаны молекулярные веса маркерных белков, в левой – молекулярные массы исследуемых пептидов. | |
5.2. Внутрипопуляционные особенности пептидного состава яда
Внутрипопуляционные различия полипептидного состава яда исследовали также электрофорезом в ПААГ, но на «индивидуальных» образцах ядовитого секрета гадюк (рис. 4). На примере яда гадюк из г. Самары заметно, что качественный состав пептидов у самок и самцов в популяциях одинаков, наблюдаются лишь некоторые количественные различия в содержании отдельных пептидов. Аналогичные результаты были получены на образцах яда гадюк из других исследованных популяций. По-видимому, различия в количественном содержании отдельных пептидов определяют различную активность ферментов в яде и обусловлены генетической гетерогенностью особей в популяции.
В ядовитом секрете гадюки Никольского (на примере популяции гадюк в Пензенской области) половые различия в пептидном составе также не обнаруживаются, но заметны различия между образцами яда разного цвета (рис. 5). В ядовитом секрете желтой окраски присутствует пептид массой 58,8 кДа, который не обнаруживается в образце бесцветного яда.
 Рис. 4. Электрофореграмма ядовитого секрета обыкновенных гадюк из г. Самары (образцы 1–3 – самки, 4–7 – самцы). Рис. 4. Электрофореграмма ядовитого секрета обыкновенных гадюк из г. Самары (образцы 1–3 – самки, 4–7 – самцы). |  Рис. 5. Электрофореграмма ядовитого секрета обыкновенных гадюк из Пензенской области Обозначения: ж – яд желтого цвета у самца; ж – яд желтого цвета у самки; б – бесцветный яд у самца; б – бесцветный яд у самки; Пж – «объединенный» образец яда желтого цвета; Пб – «объединенный» образец бесцветного яда. Рис. 5. Электрофореграмма ядовитого секрета обыкновенных гадюк из Пензенской области Обозначения: ж – яд желтого цвета у самца; ж – яд желтого цвета у самки; б – бесцветный яд у самца; б – бесцветный яд у самки; Пж – «объединенный» образец яда желтого цвета; Пб – «объединенный» образец бесцветного яда. |
| В правой колонке указаны молекулярные веса маркерных белков, в левой – молекулярные массы исследуемых пептидов. | |
5.3. Различия пептидного состава в образцах яда разного цвета
Ранее уже упоминалось, что обыкновенные гадюки в популяциях Волжского бассейна продуцируют ядовитый секрет разной интенсивности желтой окраски: ярко-желтый, желтоватый и бесцветный (см. раздел 4.2.). Различия в пептидном составе яда разных цветовых вариаций также исследовали методом электрофореза в ПААГ. Образцы ядовитого секрета 1–9 на электрофореграмме (рис. 6) расположены слева направо по мере снижения активности оксидазы L-аминокислот и уменьшения интенсивности желтой окраски. Снижение интенсивности желтой окраски и уменьшение активности оксидазы L-аминокислот яда сопровождается уменьшением количества пептида массой 58,8 кДа (рис. 7). Его максимум (13–18%) отмечен в образцах 1–3 (яд желтого цвета с максимальной активностью оксидазы L-аминокислот, полученный от гадюк номинативного подвида из Пермского края, Нижегородской и Самарской областей). В образцах 7–9 (бесцветный яд гадюки Никольского из популяций Пензенской, Липецкой и Саратовской областей), где активность оксидазы L-аминокислот не обнаруживается, пептид 58,8 кДа полностью отсутствует. В образцах 4–6 (яд желтоватого цвета с промежуточными значениями активности оксидазы L-аминокислот из популяций гадюк на границе Волжского и Донского бассейнов) его количество (4,4–6,5%) также занимает промежуточные значения.
 Рис. 6. Электрофореграмма образцов ядовитого секрета разной окраски В правой колонке приведены молекулярные массы маркерных белков. Рис. 6. Электрофореграмма образцов ядовитого секрета разной окраски В правой колонке приведены молекулярные массы маркерных белков. |  Рис. 7. Зависимость активности оксидазы L-аминокислот от содержания пептида 58,8 кДа в образцах яда обыкновенных гадюк. Рис. 7. Зависимость активности оксидазы L-аминокислот от содержания пептида 58,8 кДа в образцах яда обыкновенных гадюк. |
| Обозначения образцов: 1 – Пермский край; 2 – Нижегородская область; 3 – Самарская область; 4 – Пензенская область; 5 – Хвалынский р-н Саратовской области; 6 – граница Пензенской и Саратовской областей; 7 – Аткарский р-н Саратовской области; 8 – Липецкая область; 9 – Пензенская область. | |
Молекулярная масса, количество данного пептида в ядах гадюк и его пропорциональная зависимость от активности оксидазы L-аминокислот позволяют предположить, что пептид 58,8 кДа может являться компонентом комплекса «оксидаза L-аминокислот – ФАД», а именно – мономером оксидазы L-аминокислот (Samel et al., 2006).
Глава 6. Направленность изменений свойств ядовитого секрета
в пределах Волжского бассейна
На основании анализа свойств ядовитого секрета гадюк из популяций Волжского бассейна (табл. 3 и 4, рис. 2–6), можно выделить три группы образцов ядовитого секрета со схожими свойствами (рис. 8).
1. Яд с высокой активностью ферментов (активность протеаз более 15 мкг тирозина/мг белка в минуту, активность оксидазы L-аминокислот более 17 Е/мг белка мин), ярко-желтого цвета, с характерным набором пептидов, который продуцируют гадюки с морфологическими признаками номинативного подвида V. b. berus, встречающиеся на большей части Волжского бассейна.
2. Ядовитый секрет бледно-желтого цвета с промежуточными значениями ферментативных активностей (активность протеаз от 9 до 12 мкг тирозина/мг белка мин, активность оксидазы L-аминокислот от 2 до 10 Е/мг белка мин), который продуцируют гадюки с морфологическими признаками, характерными для обоих подвидов. Обнаружение в поволжских популяциях данной группы гадюк с промежуточными свойствами ядовитого секрета подтверждает отсутствие четкой границы распространения указанных подвидов и свидетельствует в пользу существования зоны их интерградации.
3. Бесцветный ядовитый секрет с низкими значениями активности ферментов (активность протеаз менее 10 мкг тирозина/мг белка в минуту, активность оксидазы L-аминокислот менее 1 Е/мг белка мин), который продуцируют гадюки с морфологическими признаками V. b. nikolskii в популяциях на границе бассейна Волги с бассейном Дона.

Рис. 8. Группы популяций обыкновенных гадюк
со схожими свойствами ядовитого секрета
Таким образом, изменения свойств ядовитого секрета обыкновенных гадюк в Волжском бассейне сводятся к уменьшению активности исследуемых ферментов, снижению интенсивности желтой окраски и изменению пептидного состава. Эти изменения носят постепенный характер и направлены от популяций с признаками номинативного подвида к популяциям с признаками гадюки Никольского. Наиболее выраженные морфологические признаки гадюки Никольского нами обнаружены в популяциях на водоразделе Волги и Дона. Изменения свойств ядовитого секрета в популяциях гадюк Волжского бассейна также направлены в сторону Донского бассейна, где сохранились наиболее «чистые» популяции гадюки Никольского. Причины этого явления, скорее всего, генетические и определяются сложными путями расселения и обмена генетическим материалом у обыкновенных гадюк в бассейне Волги.
Глава 7. Многомерный статистический анализ региональной
изменчивости признаков в поволжских популяциях гадюк
Поскольку наряду с изменчивостью свойств ядовитого секрета на значительной части Волжского бассейна в популяциях обыкновенных гадюк наблюдается смешение и перекрывание морфологических признаков, мы сталкиваемся со сложной проблемой оценки подвидовой принадлежности гадюк и выделения границ распространения подвидов.
Для оценки региональной составляющей изменчивости популяций мы проанализировали средние значения 9 морфологических признаков и двух показателей свойств яда методом главных компонент (рис. 9).

Рис. 9. Ординационная диаграмма морфологических признаков гадюк
и свойств их ядовитого секрета
Первая главная компонента охватывает 41% всего статистического разброса исходных показателей и связана с ведущими морфологическими признаками (C.oc., Ventr., Pf., Lor., S.lab.) и свойствами ядовитого секрета, определяющими различия подвидов V. b. berus и V.b. nikolskii. При этом вклад ферментативных активностей в распределение признаков оказался выше, чем таковой у морфологических признаков (коэффициент корреляции Пирсона r=0,936 для активности протеазы, r=0,919 для L-аминооксидазы, в то время как для морфологических признаков r=0,8 и ниже). Вторая компонента объясняет 24% вариации данных и уточняет региональную изменчивость других морфологических признаков (Ic., Lab., S.cd.). Кроме того, по совокупности морфологических признаков и свойств яда из всех анализированных популяций метод убедительно выделяет группу популяций гадюк с признаками гадюки Никольского (Липецкая, Пензенская и Саратовская области). Аналогичный результат получен при использовании метода «барьеров» (Manni et al., 2004), идентифицировавшего границы, относительно которых параметры изучаемых популяций претерпевают наиболее резкие характерные изменения. Эта граница также отделяет популяции гадюк с признаками гадюки Никольского от всех остальных.
Применение иерархической классификации по методу «ближайшего соседа» к анализу морфологических признаков гадюк и свойств их яда позволило выделить три группы популяций гадюк, соответствующие выделенным нами группам исследуемых образцов ядовитого секрета (рис. 10).

Рисунок 10. Результаты кластерного анализа морфологических признаков
и свойств ядовитого секрета гадюк в поволжских популяциях
Таким образом, независимые методы многомерной статистики указывают на то, что свойства ядовитого секрета обыкновенных гадюк в сочетании с морфологическими признаками являются более информативным показателем, чем только морфологические признаки, и свидетельствуют в пользу целесообразности использования свойств ядовитого секрета при установлении таксономической принадлежности обыкновенных гадюк в популяциях на территории Волжского бассейна.
Выводы
1. Статистически значимых подвидовых, межпопуляционных и половых различий в значениях среднесмертельной дозы ЛД50 яда обыкновенных гадюк в Волжском бассейне не выявлено.
2. Установлено, что удельная активность ферментов в яде гадюк различается как между популяциями, так и между особями внутри популяций. Статистически значимых половых различий в ферментативной активности яда во всех исследованных популяциях не обнаружено. Уровень активности протеолитических ферментов и оксидазы L-аминокислот в яде гадюк номинативного подвида статистически значимо выше, чем у гадюки Никольского.
3. Методом электрофореза в ПААГ обнаружены различия пептидного состава ядовитого секрета V. b. berus и V. b. nikolskii. Пептидный состав яда, характерный для номинативного подвида обыкновенной гадюки, отмечен на большей части Волжского бассейна, характерный для яда гадюки Никольского – на границе Волжского и Донского бассейнов и далее в бассейне Дона. Переходных форм пептидного состава ядовитого секрета не обнаружено.
4. Установлены различия активности оксидазы L-аминокислот и пептидного состава ядовитого секрета обыкновенных гадюк в образцах яда разного цвета. В образцах желтоокрашенного яда гадюк V. b. berus и V. b. nikolskii присутствует пептид массой 58,8 кДа, который не обнаруживается в образцах бесцветного яда V. b. nikolskii. Изменения цвета в образцах яда гадюк из разных популяций сопровождается изменением активности оксидазы L-аминокислот и количества пептида массой 58,8 кДа.
5. В поволжских популяциях обыкновенных гадюк отмечена общая тенденция изменения свойств ядовитого секрета – снижение активности протеолитических ферментов и оксидазы L-аминокислот, уменьшение интенсивности желтой окраски и изменение пептидного состава яда. Изменения носят плавный, постепенный характер, и эта тенденция направлена от популяций с признаками номинативного подвида к популяциям с признаками гадюки Никольского, т.е. от Волжского бассейна к Донскому.
Список работ, опубликованных по теме диссертации:
В рецензируемых изданиях, рекомендованных ВАК:
1) Маленев А. Л., Бакиев А. Г., Зайцева О. В., Шуршина И. В. Токсичность яда обыкновенных гадюк из различных пунктов ареала // Известия Самарского научного центра РАН, 2007. – Т. 9, № 1. – С. 259–261.
2) Маленев А.Л., Бакиев А.Г., Зайцева О.В., Шуршина И.В., Зиненко А.И. Протеолитическая активность яда обыкновенных гадюк из некоторых популяций России и Украины // Известия Самарского научного центра РАН, 2007. – Т. 9. – № 4. – С. 1056–1060.
3) Зайцева О.В., Маленев А.Л. Особенности биохимических свойств яда обыкновенных гадюк Vipera berus, обитающих на территории Волжского бассейна // Аграрная Россия, 2009. – Спец. выпуск. – С. 36–37.
4) Маленев А.Л., Зайцева О.В., Бакиев А.Г., Зиненко А.И. Обыкновенная гадюка на границе речных бассейнов Волги и Дона: особенности морфологии змей и свойств ядовитого секрета в популяции из Пензенской области // Современная герпетология. – 2010. Т. 10, вып. 3 / 4 – С. 115–120.
5) Павлов А.В, Зиненко А.И., Йогер У., Штумпель Н., Петрова И.В., Маленев А.Л., Зайцева О.В., Шуршина И.В., Бакиев А.Г. Естественная гибридизация гадюк восточной степной Vipera renardi и обыкновенной Vipera berus. // Известия Самарского научного центра Российской академии наук. – 2011. – Т. 13 (№ 5). – С. 172–176.
В монографиях:
6) Бакиев А.Г., Маленев А.Л., Зайцева О.В., Шуршина И.В. Змеи Самарской области. – Тольятти: ООО «Кассандра», 2009. – 170 С.
В сборниках научных трудов и докладов на научных конференциях:
7) Маленев А. Л., Зайцева О. В., Шуршина И. В., Наумкина Н. А., Павлов А. В. Токсичность и биохимические свойства яда обыкновенной гадюки Vipera berus из Республики Татарстан // Актуальные проблемы герпетологии и токсинологии: Сб. науч. тр. Вып. 9. – Тольятти, 2006. – С. 123–126.
8) Маленев А.Л., Бакиев А.Г., Шуршина И.В., Зайцева О.В. Протеолитическая активность нативного и сухого яда самцов и самок обыкновенной гадюки // Актуальные проблемы герпетологии и токсинологии: Сб. науч. тр. Вып. 9. – Тольятти, 2006. – С. 118–122.
9) Зайцева О.В., Шуршина И.В., Маленев А.Л., Бакиев А.Г. Ферментативные активности яда обыкновенной гадюки из двух популяций Среднего Поволжья // Материалы науч. конф. «Татищевские чтения: актуальные проблемы науки и практики». – Тольятти: ВУиТ, 2007. – С. 98–103.
10) Маленев А.Л., Бакиев А.Г., Зайцева О.В., Шуршина И.В. Протеолитическая активность и пептидный состав яда самцов и самок обыкновенной гадюки из Самарской области // Актуальные проблемы герпетологии и токсинологии: Сб. науч. тр. Вып. 10. – Тольятти, 2007. – С. 94–100.
11) Бакиев А.Г., Маленев А.Л., Четанов Н.А., Зайцева О.В., Песков А.Н. Обыкновенная гадюка Vipera berus (Reptilia, Viperidae) в Волжском бассейне: материалы по биологии, экологии и токсинологии // Бюлл. Самарская Лука, 2008. – Т. 17, № 4(26). – С. 759–816.
12) Маленев А.Л., Зайцева О.В., Бакиев А.Г., Ручин А.Б., Рыжов М.К. Активность ферментов и пептидный состав ядовитого секрета обыкновенных гадюк, обитающих в Республике Мордовия // Актуальные проблемы экологической физиологии, биохимии и генетики животных: Материалы II Междунар. науч. конф. – Саранск: Типография ООО «Мордовия-ЭКСПО», 2009. – С. 104–106.
13) Зайцева О.В. Полипептидный состав ядовитого секрета некоторых видов гадюк // Экологический сборник 2: Труды молодых ученых Поволжья. – Тольятти: ИЭВБ РАН, Изд-во «Кассандра», 2009. Стр. 226-229.
14) Зайцева О.В., Маленев А.Л., Бакиев А.Г., Ушаков М. В., Ползиков Д. Н. Особенности морфологии обыкновенных гадюк и биохимических свойств их яда у границ бассейна Верхней Волги // Материалы VII Международной науч.-пр. конф. «Татищевские чтения: актуальные проблемы науки и практики». – Тольятти: ВУиТ, 2010. – С. 61–67.
15) Шуршина И.В., Маленев А.Л., Зайцева О.В., Бакиев А.Г. Сравнительный анализ свойств ядов гадюк Волжского бассейна // Вопросы герпетологии. Материалы IV съезда Герпетологического общества им. А.М. Никольского. – СПб: Русская коллекция, 2011. – С. 315–318.
16) Зайцева О.В. Сравнение морфологии обыкновенных гадюк и биохимических свойств их яда из популяций Право- и Левобережья Самарской области // Экологический сборник 3: Труды молодых ученых Поволжья / Под ред. проф. С.В.Саксонова. – Тольятти: Кассандра, 2011. – С. 68–72.
17) Маленев А.Л., Зайцева О.В., Бакиев А.Г. Предварительные данные о свойствах ядовитого секрета обыкновенных гадюк Vipera berus из Национального парка «Хвалынский» // Труды НП «Хвалынский», 2010.- Вып 2. – С. 52–57.
18) Зайцева О.В., Маленев А.Л., Бакиев А.Г. Установленные лимиты значений активности протеаз и оксидазы L-аминокислот яда обыкновенной гадюки Vipera berus из популяций Среднего Поволжья // Труды Мордовского государственного природного заповедника им. П.Г. Смидовича. – Вып. IX. – Саранск; Пушта, 2011. – С. 283–286.
19) Зайцева О.В., Маленев А.Л., Бакиев А.Г. Исследования свойств яда обыкновенной гадюки в Волжском бассейне: практическое значение полученных результатов // Бюллетень «Самарская Лука: проблемы региональной и глобальной экологии». – 2011. – Т. 20, № 1. – С. 180–184.