

Эколого-биологические особенности представителей сем. onagraceae (кипрейные) в мурманской области
На правах рукописи
Асминг Светлана Викторовна
ЭКОЛОГО-БИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ
ПРЕДСТАВИТЕЛЕЙ СЕМ. ONAGRACEAE (КИПРЕЙНЫЕ)
В МУРМАНСКОЙ ОБЛАСТИ
Специальности 03.00.16 – экология
03.00.05 – ботаника
АВТОРЕФЕРАТ
диссертации на соискание учёной степени
кандидата биологических наук
Тольятти – 2009
Работа выполнена на кафедре биологии Кольского филиала
Петрозаводского государственного университета
Научный руководитель: кандидат биологических наук, доцент
Похилько Анна Антоновна
Официальные оппоненты: доктор биологических наук, профессор
Кавеленова Людмила Михайловна
кандидат биологических наук,
доцент, Дьячкова Тамара Юрьевна
Ведущая организация: Санкт-Петербургский
государственный университет
Защита состоится 22 декабря 2009 г. в 1230 на заседании диссертационного совета Д 002.251.01 при Институте экологии Волжского бассейна РАН по адресу: 445003, г. Тольятти, ул. Комзина, 10.
Тел. (8482) 48-99-77, факс (8482) 48-95-04; E-mail: [email protected]
С диссертацией можно ознакомиться в библиотеке Института экологии Волжского бассейна РАН, с авторефератом – в сети Интернет на сайте ИЭВБ РАН по адресу: http//www.ievbran.itlt.ru
Автореферат разослан «20» ноября 2009 г.
Ученый секретарь
![]()
диссертационного совета,
кандидат биологических наук А.Л. Маленев
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность исследования. Из 36 родов сем. Onagraceae в Мурманской области произрастают 3 – Chamaenerion (Rafin.) Rafin., Circaea L. и Epilobium L..
На территории Кольского полуострова встречается 1 вид рода Chamaenerion – Ch. angustifolium (L.) Scop., 1 вид рода Circaea – С. alpina L. и 8 видов рода Epilobium – E. adenocaulon Hausskn., E. alpinum L., E. alsinifolium Vill., E. collinum
C. C. Gmel., E. davuricum Fisch. ex Hornem., E. hornemannii Reichenb.,
E. lactiflorum Hausskn. и E. palustre L.
Наиболее детально изучен Chamaenerion angustifolium. Исследованием этого вида занимались многие отечественные и зарубежные ученые (Водолазский, 1982; Забелкин, Уланова, 1995; Moss, 1936; Myerscough, 1980). В отличие от Chamaenerion два других рода изучены недостаточно полно. В частности, не исследовались онтогенез, возрастные спектры, жизненность популяций и экология этих растений.
Род Circaea не включён в состав Арктической флоры СССР, поэтому изучение популяций Circaea alpina преставляет большой ботанико-географический интерес. Во Флоре северо-востока европейской части СССР А.К. Скворцов для района исследований данный вид также не приводит (1976). Кроме того, данный вид представлен изолированной популяцией и удалён от основного ареала в восточной Европе приблизительно на 1000 км.
Circaea alpina и 3 вида рода Epilobium (E. alsinifolium, E. davuricum и
E. lactiflorum) являются редкими и нуждающимися в охране на территории Мурманской области, занесёнными в региональные Красные книги. Некоторые виды рода Epilobium и C. alpina обладают полезными свойствами, являются лекарственными, кормовыми и декоративными (Растительные ресурсы СССР, 1987).
Circaea alpina, Epilobium alsinifolium и E. davuricum – кальцефильные виды, указывающие на наличие пород с присутствием извести (Kytvuori, 1962; Раменская, 1983), могут служить индикаторами плодородия почв, способствовать выявлению ценных сообществ и мест концентрации редких видов.
Цель и задачи исследования. Целью работы является детальное изучение эколого-биологических особенностей Circaea alpina и 8 видов рода Epilobium,
как в естественных местах произрастания на территории региона, так и в условиях культуры. Особое внимание уделено C. alpina на северном пределе распространения этого чрезвычайно трудно культивируемого вида.
Для достижения этой цели были сформулированы следующие задачи:
1) изучить онтогенез, возрастной состав и жизненность особей в ценопопуляциях, способы возобновления, всхожесть семян Circaea alpina и видов рода Epilobium;
2) выявить лимитирующие факторы, определить жизненные стратегии, изучить адаптированность видов на границах естественного распространения и дать прогноз дальнейшему существованию их популяций; 3) определить диагностические признаки для разграничения видов рода Epilobium.
Основные положения диссертации, выносимые на защиту:
– особенности онтогенетического развития и способов возобновления определяют возможности видов рода Epilobium и Circaea alpina к существованию
в условиях Крайнего Севера и расселению;
– адаптированность к условиям существования и распространение представителей сем. Onagraceae в Мурманской области обусловлены
как комплексом условий среды обитания, так и эколого-биологическими особенностями видов.
Научная новизна. Впервые изучены онтогенез, возрастные спектры, экологические особенности, способы возобновления, всхожесть семян видов
рода Epilobium и Circaea alpina, произрастающих на территории Мурманской области. Определено современное состояние ценопопуляций изучаемых видов,
дана оценка жизнеспособности особей в ценопопуляциях.
Практическая ценность. Изучение диагностических признаков, онтогенеза, адаптационных механизмов, возрастных спектров и жизненности ценопопуляций исследуемых видов вносит определённый вклад в систематику, биологию
и экологию растений. Дихотомический ключ для определения видов рода Epilobium представляет интерес для ботаников.
Исследование современного состояния ценопопуляций представителей
сем. Onagraceae имеет большое значение для прогнозирования развития ценопопуляций и возможности их сохранения. Сведения о географическом распространении изученных растений могут быть внесены в базы данных по редким видам. Материалы работы могут быть использованы Комитетом по природным ресурсам и охране окружающей среды Мурманской области при проектировании особо охраняемых природных территорий и при определении допустимых видов деятельности в районах произрастания.
Апробация работы. Материалы диссертации доложены и обсуждены
на конференции молодых учёных «Сбалансированное природопользование
на примере освоения минеральных ресурсов» в 2004 г. (г. Апатиты), VII научной конференции Кольского филиала Петрозаводского государственного университета
в 2004 г. (г. Апатиты), на Международной конференции Петрозаводского государственного университета «Структурно-функциональные особенности биосистем Севера» в 2005 г. (г. Петрозаводск).
Благодарности. Выражаю благодарность за руководство – к.б.н., доценту кафедры биологии КФ ПетрГУ А.А. Похилько, за консультативную помощь -
д.б.н., зав. лабораторией микробиологии Института проблем промышленной экологии Севера КНЦ РАН (ИППЭС) Г.А. Евдокимовой, научному сотруднику лаборатории наземных экосистем ИППЭС Петрову В.Н., научному сотруднику Института экологии Волжского бассейна РАН (ИЭВБ) Н.С. Ракову., научному сотруднику Полярно-альпийского ботанического сада-института им. Н.А. Аврорина КНЦ РАН (ПАБСИ) В.А. Костиной, к.б.н., старшему научному сотруднику ПАБСИ О.А. Белкиной, а также к.б.н., научному сотруднику ПАБСИ А.Ю. Лихачёву
за определение видов мохообразных.
Публикации. По теме диссертации опубликовано 8 работ, в том числе 1 –
в ведущем рецензируемом журнале, рекомендованном ВАК.
Структура и объём работы. Диссертация состоит из введения, 4 глав, выводов, библиографического списка и приложения, изложена на 161 странице машинописного текста, содержит 36 таблиц и 61 рисунок. Библиографический список включает 104 источника, в том числе 27 иностранных.
1. ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Объектами исследования являются 8 видов рода Epilobium (E. adenocaulon,
E. alpinum, E. alsinifolium, E. collinum, E. davuricum, E. hornemannii, E. lactiflorum
и E. palustre) и Circaea alpina (сем. Onagraceae).
Изучение эколого-биологических особенностей вышеназванных видов проводилась в течение 2002-2004 гг. Использовались различные методы: географический, систематический, онтогенетический, методы популяционного мониторинга, разработанные Л.Б. Заугольновой (1994), И.М. Ермаковой и др. (1987), Т.Г. Соколовой (1987) и Ю.А Злобиным (1989).
Материал собран в разных районах Мурманской области – на юго-западе (Кандалакшский район, окрестности пос. Вуориярви, ущелье Пюхякуру) и в центральной части (Кировский и Апатитский район). Кроме того, использовались материалы ПАБСИ, Ботанического Института им. В.Л. Комарова, сборы студентов
и аспирантов Кольского филиала Петрозаводского государственного университета
в окрестностях с. Умба и. Сосновка и др., собственные сборы из различных районов Мурманской области и литературные данные (Мишкин, 1953; Шляков, 1959; Раменская, Андреева, 1982; Раменская, 1983; Белкина и др., 1991).
Возрастные спектры ценопопуляций видов рода Epilobium и Circaea alpinа изучались на постоянных и полустационарных площадках. При исследовании жизненности ценопопуляций Circaea alpina использовались два метода: выделение трёх уровней жизненного состояния и многобалльная оценка жизненности (Ермакова и др., 1987). При изучении жизненности видов рода Epilobium применялся метод многобалльной оценки. Разработаны шкалы балльных оценок.
Учитывались параметры ценопопуляций – проективное покрытие (%),
обилие (по Друде), число особей разных возрастных групп (%), соотношение генеративных и вегетативных особей (%). Сделаны геоботанические описания
в местах произрастания исследуемых видов.
Материал для изучения онтогенеза видов рода Epilobium и Circaea alpina собирался методом эколого-популяционной выборки. Изучение онтогенеза проводилось в естественных местах обитания и в лабораторных условиях. Возрастные состояния выделялись по А.А. Уранову (1975). Для изучения первых этапов онтогенеза в лабораторных условиях проведены опыты по определению всхожести семян E. hornemannii и E. palustre, собранных в разных местах обитания.
Для изучения всхожести семян и первых этапов развития семена проращивали при комнатной температуре на свету, стратифицировали в течение 14 дней
при -1-0оС. Эксперимент длился 6 месяцев, проведено 3 варианта опытов
с интервалом в 2 месяца. Морфология семян, строение почек изучались
по общепринятой методике с использованием светового микроскопа МБС–10
при увеличении 4х10, 7х8.
2. СИСТЕМАТИЧЕСКИЙ ОБЗОР ПРЕДСТАВИТЕЛЕЙ СЕМ. ONAGRACEAE, ПРОИЗРАСТАЮЩИХ В МУРМАНСКОЙ ОБЛАСТИ
Сем. Onagraceae содержит около 500 видов, 36 родов, встречающихся
в умеренной зоне Нового Света. Только немногие из родов распространены
по всему свету, сюда относится в первую очередь pод Epilobium, содержащий около 250 видов. В пределах бывшего СССР он является наиболее распространённым –
53 вида (Штейнберг, 1949). Меньшее, чем Epilobium, распространение имеют: Chamaenerion Hill (7 видов) и Circaea L. (6) (Скворцов, 1989).
На территории Мурманской области произрастают представители 3-х родов: Chamaenerion, Circaea и Epilobium. Род Chamaenerion представлен 1 видом –
Ch. angustifolium. Из 7 видов рода Circaea, свойственных северной умеренной
и холодной зоне на территории региона встречается только 1 C. alpina (Шляков, 1959). В Мурманской области произрастают все перечисленные выше 6 видов рода Epilobium, редко встречаются заносный E. adenocaulon и более южный E. collinum.
В работе представлено подробное сравнение ключей разных авторов
для определения исследуемых видов. Проведён анализ морфологических и других диагностических признаков, характеризующих тот или иной вид рода Epilobium. Для определения видов рода Epilobium, произрастающих на территории Мурманской области, нами предложен дихотомический ключ, основанный
на ключах Р.Н. Шлякова (1953) и А.К. Скворцова (1980, 1991).
Виды рода Epilobium, произрастающие на территории Мурманской области, обладают основными и вспомогательными диагностическими признаками. Основными диагностическими признаками являются: органы возобновления (столоны, корневища), ветвление, длина междоузлий, форма листьев, длина
и ширина черешка, окраска и опушение коробочки и поверхность семян.
3. ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ, ФИТОЦЕНОЛОГИЯ
И ЭКОЛОГИЯ ПРЕДСТАВИТЕЛЕЙ СЕМ. ONAGRACEAE
3.1. ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ ПРЕДСТАВИТЕЛЕЙ СЕМ. ONAGRACEAE
Анализ литературных данных позволяет сделать вывод, что рассмотренные виды относятся к разным географическим элементам. Circaea alpina, Epilobium palustre и E. adenocaulon являются бореальными, E. davuricum, E. hornemannii
и E. alsinifolium гипоарктическими, E. lactiflorum арктическим, E. alpinum арктоальпийским и E. collinum плюризональным.
В Мурманской области встречаются как аборигенные представители
сем. Onagraceae, среди которых есть более южные виды, находящиеся на северном пределе ареала (Circaea alpina, Epilobium collinum), так и заносные (E. adenocaulon).
Circaea alpina имеет широкое распространение в умеренных широтах северного полушария. Характерное растение хвойных лесов. C. alpina является относительно древней (доледниковой), но в Скандинавии – представляет пришельцев, заселивших её в послеледниковое время и отчасти ещё находящихся
в процессе расселения (Толмачёв, 1974:163).
В Мурманской области известно только одно место произрастания Circaea alpina – на юго-западе Кандалакшского района. Это территория Имандровского флористического района, его юго-западного подрайона, где много богатых почвообразующих пород. Поэтому здесь встречаются арктические и бореальные представителей, требовательные к плодородию почв (Раменская, 1983). Для охраны редких видов создан комплексный региональный заказник «Кутса».
Как отмечает М.Л. Раменская (1983), 3 вида рода Epilobium (E. alsinifolium,
E. davuricum и E. hornemannii) ограничены севером региона, т.е. тундрой, лесотундрой и полосой редкостойных лесов; 2 вида (E. alpinum и E. lactiflorum) встречаются только в тундре или лесотундре, а также в горно-тундровом поясе.
E. palustre относится к видам, распространённым по всему региону,
т.е. встречающимся в естественных сообществах от тундровой зоны до подзоны средней тайги, E. collinum – к видам средней тайги.
В Мурманской области большинство видов рода Epilobium встречается
в центральной части, на юго-востоке и на севере. На востоке и северо-западе количество выявленных видов меньше, вероятно, из-за слабой изученности.
3.2. ЭКОЛОГИЯ И ФИТОЦЕНОЛОГИЯ ПРЕДСТАВИТЕЛЕЙ
СЕМ. ONAGRACEAE
3.2.1. ЭКОЛОГИЯ И ФИТОЦЕНОЛОГИЯ CIRCAEA ALPINA
По свидетельству ряда учёных (Толмачёв, 1978; Раменская, 1983)
Circaea alpina – лесной вид, характерный для более плодородных лесных почв
и хорошо развитого и поэтому довольно тенистого древесного яруса.
Это характерное растение хвойных лесов умеренных широт северного полушария.
Circaea alpina – мезофит, сциофит, кальцефил, вид ненарушенных местообитаний. Произрастает в лесах определённого типа (тенистых темнохвойных мшистых лесах), в разнотравно-моховых прирусловых ивняках по замшелым склонам, затенённым влажным расщелинам скал (Штейнберг, 1949; Скворцов, 1974; Воробьёв, 1981; Раменская, 1983). Этот вид предпочитает среднее и избыточное увлажнение, растёт на влажной перегнойной почве в тенистых сырых местах (Попов, 1957; Шляков, 1959; Рысин, Рысина, 1987; Власова, 1996).
Ущелье Пюхякуру, где была найдена популяция Circaea alpina располагается в широтном направлении и имеет склоны южной и северной экспозиции
и, следовательно, различается в экологическом, почвенном и фитоценотическом отношении. На большей части дна ущелья находится озеро. На склонах ущелья вдоль озера сформировался старовозрастный ельник разнотравно-папоротниково-зеленомошный. На склоне южной экспозиции для ценопопуляции C. alpina сложились более благоприятные условия. По опушке данного ельника сформировалась полоса, образованная ивами (Salix caprea и др.), переходящая
в сообщество прибрежно-водных видов (Caltha palustris, Filipendula ulmaria,
Galium uliginosum, Stellaria nemorum, виды рода Equisetum, Viola epipsila).
В данном фитоценозе зафиксировано 38 видов сосудистых растений.
Ярус подлеска, представленный Rubus idaeus, Ribes glabellum и R. nigrum достаточно разрежен. Из характерных спутников ели присутствуют Vaccinium myrtillus и V. vitis-idaea. Из стелющихся кустарничков следут назвать Linnaea borealis, а из плаунов Lycopodium annotinum. Травянистый ярус сложен преимущественно папоротниками: Matteuccia struthiopteris, Dryopteris carthusiana, Athyrium filix-femina, Gymnocarpium dryopteris, Phegopteris connectilis и др. В травянистом ярусе присутствует Maianthemum bifolium, Actaea erythrocarpa, Cirsium heterophyllum, Trientalis europea, из злаков и Poa nemoralis. Большинство из перечисленных видов данного ценоза обладают узкой экологической амплитудой, исчезают при вырубке леса
или нарушения древесного яруса. Это же относится и к Circaea apina. Своеобразие данного сообщества указывает на его самобытность, возраст, длительность существования, слабую нарушенность и хорошую сохранность всех его элементов.
Circaea alpina растёт на моховых подушках, в моховом покрове наблюдается разнообразие мхов (21 род, 25 видов). Мохообразные весьма гигроскопичны, являются накопителями влаги, что также создаёт благоприятные условия
для существования C. alpina. Проективное покрытие травяно-кустарничкового яруса составляет от 50 до 95%, мохово-лишайникового от 70 до 100%.
Circaea alpina часто поселяется на поваленных старых деревьях,
на начинающей разлагаться древесине в моховых подушках. Такой тип субстрата, его рыхлость является очень удобным для поселения растений,
слабо приспособленных к укоренению в уплотнённой почве. Проективное покрытие C. alpina на затенённых местах достигает 65-85%, на более освещённых – 35-40%.
Лимитирующими факторами для этого вида являются: освещение (необходимо затенение), увлажнённость (необходима средняя и избыточная), особенности почвообразующих пород (наличие кальция – карбонатных пород), проточность – в целом редкость подходящих местообитаний.
По типу жизненных стратегий Раменского (1935) и Грайма (1979)
Circaea alpina – виолент. В оптимальных экологических условиях местообитания при отсутствии антропогенного воздействия, особи C. alpina обладают достаточно высокой жизненностью и способны к расселению. В случае нарушения экологических условий или отсутствия хотя бы одного из перечисленных выше факторов популяции угнетаются и сокращаются.
К исчезновению вида могут привести как естественные изменения среды обитания (засуха), так и антропогенное воздействие – вырубки, прокладывание дорог, интенсивная рекреация, геологические разработки и мелиорация.
Но поскольку Circaea alpina произрастает на территории заказника «Кутса»,
то угрозу для вида представляет только рекреация. Кроме того, недопустимы вырубки недалеко от границ заказника, т.к. это может нарушить экологические условия и на территории заказника, что может отрицательно повлиять на фитоценоз, в котором произрастает C. alpina и другие редкие виды.
Прогноз дальнейшего существования вида в целом положительный.
Однако необходимо проводить мониторинг за состоянием и динамикой популяций.
3.2.2 ЭКОЛОГИЯ И ФИТОЦЕНОЛОГИЯ ВИДОВ РОДА EPILOBIUM
Из 12 выделенных М. Л. Раменской (1983) эколого-фитоценотических групп большинство (5) видов рода Epilobium (E. alpinum, E. alsinifolium, E. davuricum,
E. hornemannii и E. lactiflorum) отнесены к тундровым видам. Только Epilobium collinum входит в группу видов наиболее сухих и бедных почвогрунтов и большого светолюбия, довольно хорошо объединяющихся признаком ксерофильности. Встречается на сухих песчаных холмах и пустошных склонах, опушках, скалах,
в сосновых борах (Штейнберг, 1949; Раменская, Андреева, 1982; Власова, 1996).
Epilobium adenocaulon относится к заносным сорным видам – эрозиофилам (Раменская, 1983). Вид является более или менее закрепившимся в регионе, встречается по сырым и мокрым обнажениям почвы и грунта вдоль дорог,
по канавам, переувлажнённым карьерам, у гаражей, на полях, огородах, образуя большей частью открытые ценозы. Жизненность особей этого вида достаточно высокая – растения крупные, продуцируют большое количество плодов и семян.
Из аборигенных видов только Epilobium palustre относится к болотным видам. E. palustre произрастает по болотистым и илистым берегам ручьёв, рек и озёр, болот различных типов – от низинных и ключевых до переходных к верховым и в сырых тундрах (Раменская, 1983). Зафиксировано 112 видов сосудистых растений
и 26 мохообразных, произрастающих совместно с E. palustre. Господствуют травянистые многолетние (реже однолетние) растения и мохообразные. Деревья
и кустарники встречаются редко, в основном там, где проходит граница с лесом.
В отличие от Epilobium palustre, Е. davuricum – отчётливо кальцефильный вид, приуроченный к выходам карбонатных пород (Kytvuori, 1962; Раменская, 1983).
Он растёт чаще всего по ключевым болотцам или достаточно увлажнённым каменистым тундрам или тундроподобным луговинам, сырым разнотравно-ивнячково-моховым тундрам, на влажных скалах и обнажениях (Малышев, 1968; Скворцов, 1991; Воробьёв, 1981). На Кольском полуострове E. davuricum произрастает на ключевых болотцах, по берегам рек (Раменская, 1983).
Места произрастания Epilobium davuricum в районе исследования характеризуются богатыми кальцием почвами. Эвтрофное болото южного Прихибинья, где был найден E. davuricum, очень разнообразно по набору видов, является памятником природы, к настоящему времени единственное найденное нами местонахождение этого редкого вида. Совместно с Epilobium davuricum обитают 40 видов сосудистых растений и 15 мохообразных.
Местами обитания Epilobium alsinifolium являются берега рек, ручьёв,
на ключевых болотцах у выходов грунтовых вод. Как и предыдущий вид, приурочен к карбонатным породам (Раменская, 1983). Карбонатные породы (доломиты, известняки и др.) на территории Мурманской области довольно редки. Выходы этих пород не занимают больших площадей, но для растительности имеют большое значение как наиболее благоприятная в наших условиях почвообразующая порода,
и как субстрат, который обуславливает распространение растений-кальцефилов (Раменская, 1983). Строгая приуроченность рассмотренных видов-кальцефилов
к данным почвогрунтам позволяет нам считать их облигатными кальцефилами.
Анализ геологической карты Мурманской области (1996) позволяет предположить места произрастания кальцефильных видов Epilobium alsinifolium
и E. davuricum на карбонатных породах в различных районах Мурманской области, где в дальнейшем необходимо продолжить поиски данных видов.
Epilobium lactiflorum на территории Мурманской области встречается
по каменистым берегам рек, горных ручьев, на влажных скалах, ключевых болотцах, а также произрастает в тундровых луговинах и ивняках (Раменская, 1983). Этот вид отмечен нами также на антропогенно нарушенных участках, что может свидетельствовать о достаточно высокой семенном размножении и возможности быстрого заселения открытых субстратов.
М. Л. Раменская (1983) и наши исследования в области отмечают произрастание Epilobium hornemannii по берегам рек, ручьёв, озёр, на ключевых
и низинных болотах, в заболоченных ивняках и тундрах, на сырой моховой дернине у выходов грунтовых вод и по окраинам мест с поздно тающим снегом.
На 7 пробных участках с произрастанием Epilobium hornemannii нами зафиксировано 96 видов сосудистых растений и 22 мохообразных. В основном
это виды – мезо- и гигрофиты, что подтверждает приуроченность E. hornemannii
к сильно увлажнённым местам обитания. В тех местах, где Epilobium hornemannii растёт близко к лесу, встречаются лесные виды – Vaccinium vitis-idaea, Pyrola rotundifolia, Orthilia secunda, виды рода Melampyrum (M. pratense, M. sylvaticum), Trientalis europaea и др. В антропогенно нарушенных местообитаниях (интенсивная рекреация), в частности на берегу оз. Большая Имандра (окрестности ПОСВИРа),
E. hornemannii произрастает вместе с Deschampsia cespitosa, Potentilla norvegica, Rumex confertus, Taraxacum officinale, Tussilago farfara, которые считаются показателями нарушенности территории и причисляются к категории сорных видов.
Epilobium alpinum на Кольском полуострове произрастает по берегам ручьёв, речек и озёр, вдоль ключей, предпочитает влажные тундровые и горные луговины, основания склонов с поздно тающим снегом (Раменская, 1983). В экологическом отношении этот вид связан со снежными местами обитания. В сообществах,
где Epilobium alpinum произрастает совместно с E. hornemannii, в отличие от этого вида, селится на участках с более поздним сходом снега, с более низким разреженным травостоем и напочвенным покровом из мелких мхов.
Проективное покрытие видов рода Epilobium в слабо нарушенных местах
60-75%. Зафиксировано более 100 видов сосудистых растений и 25 видов мохообразных, произрастающих совместно с видами рода Epilobium. Общими сопутствующими видами сосудистых растений являются виды рода Salix – 5 видов, Carex – 4, Vaccinium – 3, Agrostis – 2, Equisetum – 2 вида, Galium – 2 вида, Melampyrum – 2, Poa – 2, по одному – 7 видов и мохообразные.
Лимитирующим фактором для всех видов рода Epilobium является увлажнённость почвы. Рассматриваемые нами виды рода Epilobium являются видами умеренного и избыточного увлажнения, исключение составляет E. collinum, который обитает на открытых и сухих местах. Жизненность популяций изученных видов может понижаться в связи с недостатком влаги в почве.
Кроме того, места произрастания исследуемых видов должны быть достаточно проточными. На непроточных, сильно заболоченных участках растения угнетаются, жизненность популяций становится низкой, исключение составляет E. palustre, который и на непроточных сфагновых болотах обладает высокой жизненностью.
Помимо перечисленных для Epilobium alsinifolium и E. davuricum важным фактором является наличие карбонатных пород, что встречается в регионе не часто.
У исследованных видов рода Epilobium, произрастающих в Мурманской области, экологическая амплитуда отличается. Широкой экологической амплитудой обладают E. alpinum, E. collinum, E. hornemannii, E. lactiflorum и E. palustre, узкой
E. alsinifolium и E. davuricum. Это можно объяснить различиями в географическом распространении, в требовательности к наличию определённых условий окружающей среды. Для изученных видов характерны определённые экологические ниши, поэтому внутривидовой конкуренции не наблюдается, как и с видами других родов и семейств. В оптимальных экологических условиях многие виды рода Epilobium образуют монодоминантные сообщества, как и другие растения.
К уязвимости Epilobium alpinum, E. alsinifolium, E. davuricum и E. lactiflorum могут привести как естественные изменения гидрологического режима,
так и антропогенное воздействие – вырубка леса, прокладывание дорог, геологические разработки, мелиоративные работы и интенсивная рекреация.
Прогноз дальнейшего существования Epilobium hornemannii и E. palustre вполне положительный. Виды проявляют достаточно высокую устойчивость
к изменению условий среды. Однако, в крайне нарушенных местах с изменением водного режима E. palustre становится угнетённым: снижаются морфометрические показатели, уменьшается количество особей единицу площади, преобладают растения меньших размеров, что соответствует низкому уровню жизненности,
в онтогенетическом спектре уменьшается количество генеративных растений.
Несмотря на то, что Epilobium alpinum и E. lactiflorum могут также встречаться также на нарушенных участках, экологическая амплитуда этих видов несколько уже, чем у E. hornemannii и E. palustre. Дальнейшее существование их популяций,
а тем более популяций редких уязвимых E. alsinifolium и E. davuricum будет зависить от степени антропогенной нагрузки на биотопы в целом и их популяции.
Согласно классификации типов поведения Раменского-Грайма изученным видам рода Epilobium свойственны различные жизненные стратегии. Заносный Epilobium adenocaulon на территории Мурманской области ведёт себя как эксплерент, замещающий аборигенные виды при сильных нарушениях среды.
В естественных местах обитания данный вид не был обнаружен, на антропогенных участках (окрестности г. Апатиты) найдены чистые заросли высокой жизненности.
Epilobium alsinifolium и E. davuricum – виоленты. В оптимальных экологических условиях – высокой увлажнённости, наличии карбонатных пород,
а также при отсутствии антропогенного воздействия популяции данных видов обладают достаточно высокой жизненностью и способны расселяться. В случае нарушения условий обитания или отсутствия хотя бы одного из перечисленных выше факторов популяции этих редких видов угнетаются и сокращаются, появляется угроза к их исчезновению.
Остальные виды рода Epilobium проявляют себя как виоленты с различной степенью патиентности. E. alpinum, E. collinum и E. lactiflorum в основном предпочитают естественные места произрастания, но могут адаптироваться и к небольшим нарушениям окружающей среды. Они становятся уязвимыми
в результате изменения общей экологической обстановки из-за вырубок леса, прокладки коммуникаций, разработок карьеров и нарастающей рекреации.
У видов Epilobium hornemannii и E. palustre патиентность выражается сильнее, что объясняет более широкое распространение на территории Мурманской области. Эти виды лучше приспособлены к меняющимся условиям среды, проявляют достаточно высокую адаптацию к антропогенно изменённым участкам, одинаково хорошо произрастают как в ненарушенных местах обитания, так и в нарушенных. Можно отметить ещё большую патиентность и пластичность E. hornemannii.
Исследованные редкие виды рода Epilobium зарегестрированы на особо охраняемых природных территориях Мурманской области: заповедниках – Лапландского, Кандалакшского; заказниках и памятниках природы. Нахождение этих редких видов на перечисленных выше особо охраняемых природных территориях позволит в определённой степени сохранить их популяции.
Тем не менее, необходим дальнейший мониторинг за динамикой и состоянием популяций. Растения можно сохранить не только в естественных местах обитания, но и в культуре. Например, Epilobium lactiflorum уже успешно культивируется на питомниках ПАБСИ, где отмечается повышение семенного размножения данного вида.
4. ОНТОГЕНЕЗ, СТРОЕНИЕ МОНОКАРПИЧЕСКОГО ПОБЕГА, ВОЗРАСТНОЙ СОСТАВ, ЖИЗНЕННОСТЬ ЦЕНОПОПУЛЯЦИЙ
И СЕЗОННОЕ РАЗВИТИЕ CIRCAEA ALPINA И ВИДОВ РОДА EPILOBIUM
4.1. Онтогенез, строение монокарпического побега, возрастной состав, жизненность особей в ценопопуляции и сезонное развитие Circaea alpina
4.1.1. Онтогенез Circaea alpina
Многие авторы, занимавшиеся ранее Circaea alpina, относят её к столонно-клубневым растениям с небольшой продолжительностью жизненного цикла (Голубев, 1956; Рысин, Рысина, 1987; Барыкина, 2000 и др.). Наши исследования показывают, что C. alpina является многолетним травянистым растением
с укороченным корневищем, в узлах которого образуются почки возобновления
и столоны. Возможно на северном пределе распространения (Мурманская область) C. alpina меняет жизненную форму, что говорит об адаптации этого более южного растения к крайним для него условиям существования. На территории региона
C. alpina является многолетним растением, подземные органы которого сохраняются в течение нескольких лет. Во флоре СССР (1949) C. alpina характеризуется как травянистый корневищный многолетник, у которого образуется продолговатые клубеньки и тонкие надземные побеги (Штейнберг, 1949:634).
Возможность образования столонов и надземных побегов указывает
на возможность вегетативного размножения вида при затруднении семенного.
Семена с прямым зародышем без эндосперма. Сведения о прорастании семян этого вида нам не известны. Семена с сильным экзогенным и глубоким физиологическим покоем (Николаева, Разумова, 1985; Kinzel, 1913). Эксперименты по изучению всхожести свежесобранных семян Circaea alpina не дали положительных результатов.
Проростки (p) Circaea alpina имеют первичный корешок и 2 семядоли овально-яйцевидной формы, которые не отмирают после образования настоящих листьев в течение 4-6 дней (рис. 1). На конусе нарастания закладываются листовые зачатки. Зародышевый корешок функционирует до перехода растения в имматурное состояние.
Переход в ювенильное (j) состояние характеризуется образованием первой пары настоящих листьев (рис. 1). Ювенильные растения Circaea alpina – однопобеговые. На этой стадии формируются первая и вторая пары настоящих листьев. Главный корень сохраняется, формируются боковые корни первого порядка.
Переход в имматурное (im) состояние характеризуется тем, что листья становятся похожи на взрослые, приобретая сердцевидную форму; побеги имеют
2 междоузлия и 3 пары листьев (рис. 1). В зависимости от расположения на побеге варьируют размеры и форма листьев, количество зубцов. В основании первого узла начинают формироваться 2 боковых побега. Происходит отмирание главного корня, образуется корневище на глубине 2-3 см. В пазухах листьев и на корневище закладываются почки возобновления (1-2).
Рис. 1. Особи Circaea alpina в прегенеративный период (бп – боковой побег,
вп – верхушечная почка, г/к – гипокотиль,
зк – зародышевый корешок, к – корневище,
м – междоузлия, о/л – отмершие листья,
пк – придаточные корни, с/д – семядоли,
э/к – эпикотиль, 1 л – первая пара листьев,
2 л – вторая пара листьев и т. д.)

Переход в виргинильное (v) состояние сопровождается интенсивным нарастанием надземной и подземной частей растений, образованием 4-й пары листьев, формированием боковых побегов второго порядка (1-2) из пазушных почек (рис. 1). Ветвление базитонное. Строение боковых побегов идентично главному. Побеги имеют
3 междоузлия. Листья
в количестве 3-4 пар, крупные, сердцевидно–яйцевидной формы, имеют хорошо выраженные зубцы по всему краю. В отличие от имматурных, у виргинильных особей на корневище, состоящем из 2-4(5) междоузлий, образуются уже не единичные,
а многочисленные почки возобновления, которые закладываются равномерно на всём протяжении корневища. У второго междоузлия появляются придаточные корни.
С образованием первого генеративного побега растения Circaea alpina переходят в генеративное состояние.
Молодые генеративные (g1) особи Circaea alpina имеют 3-4 междоузлия
(рис. 2). Листья в количестве 3-4 пар, крупные, с хорошо выраженными зубцами
по краю. Первая пара листьев отмирает, в их пазухах развиваются боковые побеги. Развиваются 1-2 боковых побега, несущие по 3-4 пары листьев. На верхушке побега формируется простое соцветие кисть. На корневище, помимо почек возобновления, образуются столоны. Столоны подземные, состоят из нескольких пар мясистых бесцветных чешуй, междоузлий и верхушечной почки. Нарастание моноподиальное. В пазухах чешуй столона находятся почки возобновления. Верхушечная почка столона состоит из чешуй, листовых бугорков и конуса нарастания.
Рис. 2. Генеративные особи Circaea alpina
(бп – боковой побег, к – корневище, м – междоузлия,
о/л – отмершие листья, пк – придаточные корни,
пл – плоды, с – соцветие, ст – столоны, ц – цветки,
1 л – первая пара листьев, 2 л – вторая пара листьев)

У средневозрастных (g2) особей на верхушке побега сформировано кистевидное соцветие, состоящее из главной оси и 2-4 паракладиев, каждый из которых несёт по (5)6-8(10) цветков (рис. 2). Побеги имеют 5-7 междоузлий. Листорасположение может быть как супротивным, так и очередным. Листья
в количестве 4-5 пар, крупные, сердцевидно-яйцевидной формы, имеют хорошо выраженные зубцы по краю листа.
На корневище продолжают формироваться почки возобновления и столоны. Происходит вегетативное размножение. За счёт образования столонов особь разрастается, увеличивается занимаемая ею площадь.
В следующий вегетационный период столоны отмирают, связь
с материнским растением прерывается, дочерние особи начинают самостоятельное существование.
У старых генеративных (g3) особей Circaea alpina начинается упрощение структуры надземных и подземных частей. Листья в количестве 4(5) пар,
менее крупные, чем у средневозрастных особей, сердцевидно-яйцевидной формы, зубцы по краю листа становятся менее выраженными. Отмирают старые участки корневища и первая пара листьев, столоны продолжают формироваться,
но уже в меньшем количестве (1-2).
Сенильные (s) и субсенильные (ss) особи Circaea alpina в природе
не обнаружены. Это объясняется тем, что C. alpina является вегетативно-подвижным растением. Благодаря наличию корневищ и формирующимся на них столонам особи постоянно омолаживаются и расселяются.
4.1.2. Строение монокарпического побега Circaea alpina
На многолетнем корневище Circaea alpinа у особей разных возрастных состояний, начиная с имматурных, развиваются двулетние монокарпические побеги.
Во второй половине вегетационного сезона формируются почки возобновления (рис. 3). В количестве от 4 до 10 они располагаются в пазухах бесцветных пленчатых чешуй на коротком корневище, состоящем из нескольких междоузлий. Почка состоит из бесцветных чешуй (1-2) с широкими основаниями, двух листовых зачатков и конуса нарастания.
Рис. 3. Строение почек и столонов Circaea alpina
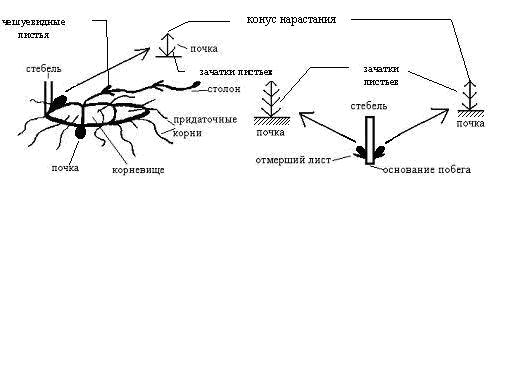
Почки возобновления после периода покоя в следующий вегетационный сезон дают начало новым метамерам подземных побегов с чешуевидными листьями.
К концу августа рост подземных побегов замедляется, на корневище развиваются придаточные корни, а в пазухах чешуевидных листьев формируются почки возобновления (1-3). Весной из почки формируется надземный побег с листьями, расположенными супротивно. Побег заканчивается соцветием. Цветоносный побег осенью отмирает, его базальная часть включается в систему подземных корневищ.
У основания надземных побегов находятся почки столонов (1-3).
Столон состоит из нескольких пар чешуй, междоузлий и верхушечной почки.
Длина столона 7-13.5 см. В пазухах чешуй столона расположены почки. Верхушечная почка состоит из чешуй, листовых бугорков и конуса нарастания.
Таким образом, многолетний поликарпик Circaea alpina формирует побеги дициклического типа, т. е. длительность их развития от пробуждения начальной почки до цветения и отмирания составляет 2 вегетационных периода.
4.1.3. Возрастной состав ценопопуляций Circaea alpina
Возрастные спектры ценопопуляции Circaea alpina, найденной в ущелье Пюхякуру неполночленные, левосторонние: господствуют особи прегенеративного периода (v) – 32% (рис. 4). Ювенильные (j) особи составляют 11% возрастного спектра, имматурные (im) – 17%, средневозрастные (g2) – 15%, старые генеративные особи (g3) – 18%. Проростки (p) и молодые генеративные особи (g1) составляют небольшую долю спектра (1 и 6%), сенильные (s) и субсенильные (ss) особи отсутствуют. Плотность ценопопуляции 37 шт. на 1 м2. В целом в ценопопуляции
C. alpina преобладают вегетативные особи (61%).
Изученная ценопопуляция Circaea alpina разновозрастная. Значительную роль играют способные к плодоношению генеративные особи. Тип возрастного спектра ценопопуляции является равновесным. Изученная популяция достаточно стабильна.

4.1.4. Жизненность особей в ценопопуляции Circaea alpina в Мурманской области
Жизненность Circaea alpina изучалась на той же ценопопуляции.
1. Выделение трёх уровней жизненного состояния в онтогенезе особей и применение этого метода для характеристики ценопопуляции Circaea alpina.
Рис. 4. Возрастной спектр ценопопуляции Circaea alpina
Первый уровень – низкая жизненность (небольшие размеры побегов и листьев, небольшое количество листьев и репродуктивных органов, отсутствием боковых побегов). Второй уровень – средняя жизненность (нормальные размеры побегов и листьев, большое количество листьев и репродуктивных органов и боковых побегов. Третий уровень – высокая жизненность (максимальные размеры побегов и листьев, большое количество листьев, репродуктивных органов и боковых побегов).

В исследованной ценопопуляции Circaea alpina в Мурманской области во всех возрастных группах преобладают особи с максимальным уровнем жизненности. Большую часть ценопопуляции составляют растения средних и максимальных размеров, т.е. 2-го и 3-го уровней жизненности (рис. 5).
Рис. 5. Возрастной спектр ценопопуляции Circaea alpina
с учётом дифференциации возрастных групп по уровням жизненности особей
В прегенеративном периоде самая высокая жизненность наблюдается у виргинильных растений (62% – 3-го, 33% – 2-го и 5% – 1-го уровня жизненности) и наименьшая
у ювенильных (36% – 3-го, 43% – 2-го и 21% –
1-го уровня жизненности). В генеративном периоде более мощными являются средневозрастные растения (76% – 3-го, 21% – 2-го и 3% – 1-го уровня жизненности), наименьшая жизненность у молодых (44% – 3-го, 44% – 2-го и 12% – 1-го уровня жизненности).
Жизненность особей прегенеративной
и генеративной части одинаковая. Особи 3-го и 2-го уровня жизненности составляют 52-52 и 37-42% соответственно. В целом можно сказать, что ценопопуляция находится
в нормальных условиях окружающей среды.
2. Метод многобалльной оценки жизненности особи и его применение
для характеристики ценопопуляции Circaea alpina.
Для того чтобы использовать более точные методы оценки жизненного состояния особей Circaea alpina, нами была разработана трёхбалльная шкала, основанная на численных характеристиках. В основу шкалы было взято 4 признака – высота побега, количество листьев, длина и ширина листовой пластинки, а также количество генеративных органов (цветков/плодов) для генеративных особей,
по которым были проанализированы особи всех возрастных групп.
В ценопопуляции преобладают особи со 2-м баллом жизненности, меньше особей с 1-м баллом и очень мало с 3-м баллом, причем по всем возрастным группам. Особи прегенеративной и генеративной части показывают примерно одинаковый разброс по уровням жизненности, у генеративных много особей со 2-м баллом, меньше с 1-м и с 3-м, а у вегетативных тоже много со 2-м, но больше с 1-м и меньше с 3-м.
Несмотря на то, что при применении второго метода показатели жизненности оказались ниже, чем при оценке по первому методу, что второй метод является более точным, поскольку он основан на численных характеристиках. Первый метод является более субъективным.
Оценка жизненного состояния показала, что ценопопуляция Circaea alpina обладает средней жизненностью, не процветает, и не угнетена – находится в пределах нормы, но отметим, что говорить об оптимальных условиях существования этого вида нельзя, так как Мурманская область является северным пределом его распространения.
4.1.5. Сезонное развитие Circaea alpina в Мурманской области
Circaea alpina – летнезелёное растение. Вегетация Circaea alpina начинается после таяния снега в конце мая – начале июня (иногда в середине июня).
Период цветения небольшой и длится всего 10 дней в годы с высокой температурой воздуха, как в 2003 году. Первые зрелые семена появляются в конце первой декады августа. Созревание плодов и семян идёт в течение 10-15 дней. Семена созревают
не каждый год. Отмирание растений происходит в конце августа – начале сентября, но может быть и раньше, в зависимости от погодных условий.
4.2. Онтогенез, строение монокарпического побега, возрастной состав, жизненность особей в ценопопуляциях и сезонное развитие
видов рода Epilobium
4.2.1. Всхожесть семян видов рода Epilobium
Скорость и продолжительность прорастания семян у исследованных видов рода Epilobium различны. Семена E. davuricum не проросли ни в одном из вариантов опыта. Семена E. hornemannii и E. palustre прорастают через 7-13 суток, максимальный процент всхожести наблюдается на 15-25-е сутки у семян обоих видов, не подвергавшихся действию низких температур, на 26-32 суток
у стратифицированных семян E. hornemannii и на 8-20-е сутки
у стратифицированных семян E. palustre.
Высокой всхожестью характеризуются семена Epilobium hornemannii во всех вариантах опыта, собранные в разных местах обитания (20-75%). Стратификация низкими температурами отрицательно сказывается на всхожести семян этого вида (1-32%). Семена E. palustre по сравнению с E. hornemannii имеют низкую всхожесть в опытах при комнатной температуре (2-7%), их всхожесть повышалась
после воздействия низкими температурами (25-56%), и прорастали они быстрее.
В природе семена видов рода Epilobium прорастают на следующий год.
4.2.2. Онтогенез видов рода Epilobium
Ввиду того, что возрастное развитие разных видов рода Epilobium незначительно отличается, онтогенез рассматривается на примере одного вида –
E. hornemannii. Для других видов указаны существенные характерные отличия.
Epilobium hornemannii – многолетнее травянистое растение с коротким корневищем, в узлах которого образуются почки возобновления. В отличие от
E. hornemannii, корневище E. alpinum удлинённое ползучее. У E. alsinifolium
и E. palustre на корневище помимо почек возобновления, образуются также специализированные органы вегетативного размножения – столоны (подземные
у E. palustre и надземные у E. alsinifolium).
Проростки (p) имеют белый тонкий первичный корень и две семядоли овальной формы, которые не отмирают после образования настоящих листьев
в течение 5-7 дней. Зародышевый корешок функционирует до перехода растения
в имматурное состояние. На конусе нарастания закладываются листовые зачатки. Проростки E. hornemannii, а также E. palustre появляются и в ненарушенных местах обитания, и в подверженных антропогенной нагрузке. Одним из главных условий для прорастания является достаточное увлажнение почвы. Проростки других видов, чувствительных к антропогенной нагрузке, встречаются только на слабо нарушенных участках, в увлажненных местах с умеренным плодородием почвы.
Переход в ювенильное (j) состояние характеризуется образованием первой пары настоящих листьев. Ювенильные растения E. hornemannii – однопобеговые, первое и второе междоузлия укорочены. На этой стадии формируются первая
и вторая пары настоящих листьев, которые располагаются супротивно.
Главный корень сохраняется, формируются боковые корни.
Переход в имматурное (im) состояние характеризуется тем,
что листорасположение меняется с супротивного на спиральное (кроме Epilobium alsinifolium, E. davuricum и E. palustre), листья становятся похожи на взрослые, приобретая яйцевидную форму. Побеги имеют 3 пары листьев и 2 междоузлия.
В зависимости от расположения на побеге размеры и форма листьев варьируют.
У основания главного побега в пазухах первой и второй пары листьев закладываются почки возобновления. Главный корень отмирает, идёт интенсивное образование придаточных корней и формирование корневища на глубине 2-3 см.
На ранних этапах онтогенеза у имматурных особей всех исследованных видов рода Epilobium форма листьев отличается и является постоянным диагностическим признаком: линейная (E. davuricum), узколанцетная (ланцетная) (E. palustre), широколанцетная (E. adenocaulon), овальная с закруглённой верхушкой (E. alpinum), яйцевидная (E. alsinifolium, E. collinum). В отличие от всех остальных видов,
у особей E. alpinum корневище не короткое, а длинное (ползучее).
Переход в виргинильное (v) состояние сопровождается интенсивным нарастанием надземной и подземной частей растений. Виргинильные особи
E. hornemannii имеют листья в количестве 3-4 пар, вполне сформированы, приобретают взрослую форму. Появляется хорошо выраженное сетчатое жилкование листьев. Идёт нарастание корневища. В отличие от имматурных,
у виргинильных особей на корневище образуются уже не единичные,
а многочисленные почки возобновления. У второго междоузлия появляются придаточные корни. У E. alsinifolium и E. palustre в пазухах низовых листьев
у основания побега на глубине 2-3 см формируются специализированные органы вегетативного размножения – столоны, нарастающие моноподиально.
С образованием первого генеративного побега растения переходят
в генеративное (g) состояние. Особи E. hornemannii характеризуются максимальным развитием надземных и подземных частей. Листья в количестве 3-4 пар, яйцевидной формы. Низовые листья отмирают, в их пазухах формируются боковые почки.
На верхушке побега формируется простое кистевидное соцветие, несущее
(3)5-6 цветков. Преобладают придаточные корни, образующиеся на нижних междоузлиях стебля. На корневище продолжают формироваться почки возобновления, а у видов E. alsinifolium и E. palustre и столоны длиной 6-9 см, имеющие по 7 междоузлий и 3-5 пар плёнчатых листьев. Происходит вегетативное размножение. За счёт образования корневища и столонов особи разрастаются, увеличивается занимаемая ими площадь. В следующий вегетационный период
связь с материнским растением прерывается, дочерние особи начинают самостоятельное существование.
4.2.3. Строение монокарпического побега видов рода Epilobium
В отличие от Circaea alpina, у видов рода Epilobium длительность развития монокарпических побегов от пробуждения начальной почки до цветения
и отмирания составляет 1 вегетационный период, т. е. побеги моноциклического типа. Побеги, образующиеся из почек на концах столонов или из придаточных почек на корневище, сначала проходят внутрипочечное развитие (в этот же вегетационный период), а внепочечное уже в следующем году.
4.2.4.-4.2.5. Возрастной состав и жизненность особей в ценопопуляциях видов рода Epilobium
Возрастные спектры ценопопуляций видов рода Epilobium изучены неравномерно. Обследовано больше ценопопуляций видов E. hornemannii,
E. palustre, широко распространённых в регионе, в то время как популяции
E. davuricum, встречающийся в специфических местообитаниях, изучены значительно меньше. Для видов разработаны трёхбалльные шкалы жизненности.
Анализ возрастных спектров ценопопуляций видов рода Epilobium указывает на то, что этим видам с вторичной жизненной стратегий (виоленты-патиенты) свойственна различная степень патиентности. У E. davuricum и E. palustre, которые образуют больше генеративных особей в местах, не подверженных антропогенному изменению, а в нарушенных местах обитания больше вегетативных особей
(78%-84%) патиентность выражена слабее. В отличие от них E. hornemannii образует больше генеративных особей в нарушенных местообитаниях (53-59%), проявляет себя как патиент достаточно хорошо. Нарушение окружающей среды
не препятствует прохождению онтогенетических стадий особям данного вида.
Особи всех видов обладают невысокой жизненностью (1-2-й уровни).
Самая высокая жизненность у особей E. hornemannii (2-3), меньше – у E. alpinum (2), E. palustre (2) и E. palustre var. lapponicum (2), самая низкая жизненность
у E. lactiflorum (1). У генеративных особей жизненность ниже, чем у вегетативных особей. Это, вероятно, можно объяснить тем, что в годы исследований вегетационные периоды были аномально жаркими.
Самая высокая жизненность у особей E. alpinum (3), средняя – у E. davuricum (2), E. hornemannii (2) и E. lactiflorum (2), низкая – у E. palustre (1-2) и E. palustre var. lapponicum (1-2).
4.2.6. Сезонное развитие видов рода Epilobium в Мурманской области
Виды рода Epilobium – летнезелёные растения. Фенологическое развитие продолжается в течение 2-2,5 месяцев. Вегетация начинается после таяния снега
в конце мая – начале июня, в некоторые годы в середине июня. Продолжительность созревания семян короче, чем у Сircaea alpina. Семена созревают ежегодно.
В конце августа начинается пожелтение листьев. Отмирание растений происходит
в конце августа – начале сентября. Продолжительность фенологических фаз исследованных видов рода Epilobium идентична. Межфазные периоды короткие.
4.3. Способы размножения и возможности к расселению представителей
cем. Onagraceae
На основании изучения онтогенеза, морфологических особенностей, всхожести семян исследуемых видов, их можно разделить на следующие группы
по способам размножения:
1. Виды, одинаково хорошо размножающиеся как вегетативным (участками корневища), так и семенным способами (Epilobium alpinum, E. adenocaulon,
E. collinum, E. hornemannii, E. lactiflorum). 2. Виды, одинаково хорошо размножающиеся как вегетативно при помощи специализированных органов (столонов), так и семенами (E. palustre). 3. Виды, возобновляющиеся преимущественно вегетативно при помощи столонов (Circaea alpina,
E. alsinifolium). 4. Виды, возобновляющиеся преимущественно вегетативно участками короткого корневища (E. davuricum).
Среди исследованных видов рода Epilobium лучше всего расселяются
E. palustre и E. hornemannii, т.к. у них очень хорошо развито как вегетативное,
так и семенное размножение, они обладают высокой семенной продуктивностью
и всхожестью семян, достаточно неприхотливы к условиям среды.
Epilobium alpinum и E. lactiflorum хорошо возобновляются как вегетативным способом при помощи корневища, так и семенным путём.
Широкое распространение E. alpinum в регионе может свидетельствовать
о достаточно высокой семенной продуктивности, способствующей расселению данного вида в благоприятных экотопах. Семенная продуктивность и всхожесть семян E. lactiflorum довольно высокие, что позволяет растению занимать вторичные местообитания. Вид достаточно успешен в культуре и неприхотлив.
Небольшая семенная продуктивность и всхожесть, а также стенотопность Circaea alpina, Epilobium alsinifolium и E. davuricum не позволяют этим видам распространяться на большие территории. Расселение Circaea alpina и E. alsinifolium происходит преимущественно вегетативно при помощи столонов на небольшие расстояния в подходящих условиях обитания. E. davuricum размножается участками короткого корневища, что не позволяет данному виду расселяться на большие площади в отличие от столонообразующих.
Заносный вид Epilobium adenocaulon впервые был зарегестрирован в регионе
в конце 1950х гг. За прошедшее время активно расселился и по-прежнему приурочен к вторичным местам обитания. E. adenocaulon очень хорошо размножается как семенным, так и вегетативным способами (участками корневища).
В отличие от всех предыдущих видов более южный вид Epilobium collinum занимает более сухие и открытые местообитания. Широкому распространению этого вида кроме вегетативного размножения (участками корневища) способствует высокое семенное.
Исследованные нами представители сем. Onagraceae занимают определённые экологические ниши, соответствующие их биологии. Эколого-биологические особенности этих видов содействуют распространению и нахождению их
в соответствующих биотопах, хотя возможности видов к расселению разные.
ВЫВОДЫ
1. В результате проведённого анализа онтогенеза и способов размножения подтверждено, что исследованные виды рода Epilobium и Circaea alpina относятся к многолетним травянистым корневищным растениям. Большинство из них являются короткокорневищными растениями, а отдельным из них (E. alsinifolium, E. palustre
и C. alpina) характерно также столонообразование. Длительность развития побегов C. alpina и видов рода Epilobium различна: C. alpina формирует монокарпические побеги дициклического типа, виды рода Epilobium – моноциклического.
Размножение у исследованных видов происходит как вегетативным,
так и генеративным способами. Семена изученных видов имеют разную всхожесть
и длительность прорастания. Некоторые виды (Circaea alpina и Epilobium davuricum) в лабораторных условиях характеризуются отрицательной всхожестью,
у других видов (E. hornemannii и E. palustre) всхожесть высокая.
В возрастных спектрах изученных видов в ненарушенных условиях существования преобладают генеративные особи высокой жизненности. Исключение составляет Epilobium hornemannii, популяции которого не угнетаются даже в условиях нарушения, особи достигают генеративного состояния.
2. Исследованные виды на территории Мурманской области достаточно хорошо адаптированы к условиям существования, занимают определённые экологические ниши. Распространение и нахождение в соответствующих биотопах обусловлено их эколого-биологическими особенностями, однако, возможности
к расселению у видов разные.
Лимитирующими факторами для произрастания большинства видов является увлажнённость, наличие карбонатных пород для кальцефильных представителей
и затенение (у сциофита C. alpina). Жизненное состояние ценопопуляций относительно благополучное, прогноз дальнейшего существования положительный.
Изученным видам характерны различные жизненные стратегии,
что подтверждается результатами изучения возрастных спектров, жизненности ценопопуляций, географического распространения и эколого-фитоценотических особенностей. Редкие в области Circaea alpina, Epilobium alsinifolium и E. davuricum требовательные к качеству окружающей среды и не способные выживать в условиях нарушения, являются виолентами. Остальные виды рода Epilobium характеризуются в различной степени переходом от конкурентной стратегии к стресс-толерантной, среди которых наибольшей патиентностью характеризуется E. horemannii.
3. На основании изучения морфолого-биологических особенностей видов рода Epilobium выявлены основные и вспомогательные признаки для их разграничения. Основными диагностическими признаками изученных видов являются: органы возобновления (столоны, корневища), ветвление, длина междоузлий, форма листьев, длина и ширина черешка, окраска и опушение коробочки и поверхность семян. Разработан дихотомический ключ для определения исследованных видов.
ПУБЛИКАЦИИ В РЕЦЕНЗИРУЕМЫХ ИЗДАНИЯХ, РЕКОМЕНДУЕЫХ ВАК:
Асминг С.В. Морфолого-биологические особенности и экология видов рода Epilobium L. в Мурманской области // Вест. Оренбургского гос. ун-та. – Оренбург, №10(74), Октябрь, 2007, С. 128-134.
СПИСОК ПУБЛИКАЦИЙ ПО ТЕМЕ ДИССЕРТАЦИИ:
- Асминг С.В. Экология и возрастные спектры популяций некоторых видов рода Epilobium L. Кольского полуострова // Кольский полуостров на пороге третьего тысячелетия: проблемы экологии. – Апатиты: Изд-во КНЦ РАН, 2003. – С. 84-90.
- Асминг С.В. Влияние антропогенного фактора и метеорологических условий на жизненность популяции Epilobium davuricum Fisch. ex Hornem. на эвтрофном болоте южного Прихибинья // Материалы школы молодых учёных «Сбалансированное природопользование на примере освоения минеральных ресурсов». – Апатиты: Изд-во КНЦ РАН, 2004. – С.138-141.
- Асминг С.В. Возрастные спектры и экология Circaea alpina L. в Мурманской области // VII Научная конференция КФ ПетрГУ. – Апатиты, 2004. – С. 35-36.
- Асминг С.В. Материалы к биологии некоторых видов рода Epilobium L., произрастающих в Мурманской области // Материалы докл. II Научной школы молодых специалистов РАН «Комплексность использования минерально-сырьевых ресурсов – основа повышения экологической безопасности региона». – Апатиты: Изд-во КНЦ РАН, 2005. – С. 99-103.
- Асминг С.В. Жизненность ценопопуляций некоторых видов рода Epilobium L., произрастающих в Мурманской области // Материалы междунар. конф. «Структурно-функциональные особенности биосистем Севера». – Петрозаводск: Изд-во ПетрГУ, 2005. – С. 27-29.
- Асминг С.В. Жизненность особей разных возрастных состояний ценопопуляции Circaea alpina L. в Мурманской области // Материалы международного контактного форума по сохранению местообитаний в Баренцевом регионе «International compact forum on habitat conservation in the Barents region». – Сыктывкар, 2005. – С. 13-14.
- Асминг С.В. Формирование жизненной формы представителей семейства Onagraceae в условиях Крайнего Севера (Мурманская область) // Материалы II Всеросс. науч. конф. «Принципы и способы сохранения биоразнообразия». – Йошкар-Ола, 2006. – С. 27-28.