

Исследование взаимодействия полностью транс -ретиналя с компонентами фоторецепторной мембраны и ретиналь-переносящими белками
На правах рукописи
Аболтин Павел Валерьевич
ИССЛЕДОВАНИЕ ВЗАИМОДЕЙСТВИЯ ПОЛНОСТЬЮ ТРАНС-РЕТИНАЛЯ С КОМПОНЕНТАМИ ФОТОРЕЦЕПТОРНОЙ МЕМБРАНЫ И РЕТИНАЛЬ-ПЕРЕНОСЯЩИМИ БЕЛКАМИ
03.01.02 – биофизика
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Москва, 2013
Работа выполнена в Федеральном государственном бюджетном учреждении науки Институте биохимической физики им. Н.М. Эмануэля Российской академии наук
| Научные руководители: | доктор биологических наук Каламкаров Григорий Рафаэлевич, заведующий лабораторией нейрохимии Института биохимической физики им. Н.М. Эмануэля Российской академии наук доктор химических наук Левин Петр Петрович, ведущий сотрудник лаборатории физико-химических основ регуляции биологических систем Института биохимической физики им. Н.М. Эмануэля Российской академии наук |
| Официальные оппоненты: | доктор физико-математических наук, профессор, Твердислов Всеволод Александрович заведующий кафедрой биофизики Физического факультета Московского Государственного Университета им. М.В. Ломоносова доктор биологических наук Тимошин Александр Анатольевич, старший научный сотрудник Федерального государственного бюджетного учреждения науки «Российский Кардиологический Центр» Министерства Здравоохранения Российской федерации |
| Ведущая организация: | Федеральное государственное бюджетное учреждение науки Институт «Международный томографический центр» Сибирского отделения Российской академии наук |
Защита состоится «19» февраля 2014 года в 13.30 часов на заседании Диссертационного совета Д 002.039.01 при Федеральном государственном бюджетном учреждении науки Институте биохимической физики им. Н.М. Эмануэля Российской академии наук по адресу: 119334, Москва, ул. Косыгина, д.4.
С диссертацией можно ознакомиться в библиотеке Федерального государственного бюджетного учреждения науки Института химической физики им. Н.Н. Семенова Российской академии наук.
Автореферат разослан «____» ______________ 2014 г.
| Ученый секретарь диссертационного совета, кандидат химических наук | Л.И. Мазалецкая |
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы
Основная функция глаза – световосприятие и преобразование светового кванта в различимый сенсорный сигнал. Зрительный аппарат развивался для выполнения именно этой функции и максимально для нее приспособлен. Для восприятия света в широком диапазоне освещенностей сформировалось много как клеточно-молекулярных, так и физических механизмов адаптации органа зрения к тому диапазону освещенностей, в котором происходит функционирование того или иного вида животных. Однако, кроме того, что свет является носителем сенсорного сигнала, он может выступать и как физиологически опасный фактор. Причиной этого является то, что, взаимодействуя с различными светопоглощающими веществами – хромофорами - квант света может инициировать образование активных форм кислорода (АФК), которые, в свою очередь, могут оказывать повреждающее действие на молекулярные и клеточные структуры глаза, такие, как хрусталик, роговица и сетчатка [Островскиий,1982]. Хотя сам факт, что пребывание экспериментальных животных в условиях высоких освещенностей приводит как к снижению зрительных функций, так и к заметным морфологическим изменениям, известен давно[Delmelle M.,1977; Островский М.А., 1982], особую актуальность эта проблема получила в последнее время, когда было установлено, что в сетчатке при относительно высокой освещенности развивается апоптоз. Поскольку сетчатка является постмитотической нервной тканью и не восстанавливается при гибели клеток, развитие апоптоза на протяжении всей жизни может иметь прямое отношение как к старению сетчатки, так и к развитию старческих патологий.
В сетчатке не так много хромофоров, и поиск хромофоров, ответственных за развитие апоптоза, а также механизмов светоиндуцированной генерации свободных радикалов имеет первостепенное значение как в фундаментальном, так и в прикладном смысле.
Одним из потенциальных хромофоров – генераторов свободных радикалов - является свободный полностью транс-ретиналь (ATR) [Delmelle M., 1978; Островский, 1982]. Ретиналь в 11-цис конфигурации является хромофорной группой светочувствительного белка родопсина, который находится в фоторецепторной клетке в очень высокой концентрации (3-4 ммоль/л). При освещении он переходит в полностью транс-форму и диссоциирует, находясь в фоторецепторной мембране в полностью транс-форме. Действительно, известно, что ATR способен при освещении генерировать как синглетный кислород, так и супероксид анион-радикал с относительно высоким квантовым выходом (0.001). Однако, высвобождаясь из белка, ATR восстанавливается в ретинол ферментом ретинолдегидрогеназой (РДГ), причем активность РДГ настолько высока, что in vivo свободный ретиналь не обнаруживается в сетчатке даже при распаде 90% родопсина при очень сильной освещенности. Ретинол же, в отличие от ретиналя, не является сенсибилизатором, способным генерировать свободные радикалы. В стабильном состоянии ретиналь в обеих изомерных формах в сетчатке встречается только в связанном с ретиналь-переносящими белками состоянии. В связи с этим важным представляется вопрос о том, является ли ATR фотосенсибилизатором, будучи связанным с ретиналь-переносящими белками. Накапливаясь при высокой освещенности внутри фоторецепторного диска, ATR формирует Шиффовы основании с аминогруппами белков и липидов, которые не являются фототоксичными. Однако, некоторое его количество может оставаться и в свободном виде.
Цель диссертационной работы
Цель диссертационной работы заключалась в исследовании фотосенсибилизирующих свойств ретиналя в связанном с ретиналь-переносящими белками состоянии, а также в оценке возможных концентраций свободного ретиналя в фоторецепторной мембране в том случае, когда он по каким-либо причинам не восстанавливается в ретинол, и оценке квантовых выходов фотоповреждения белков свободным ATR.
Основные задачи исследования
Определить, какие активные формы кислорода образуются при освещении ATR в растворе и в состоянии, связанном с ретиналь-связывающими белками.
Методом спиновой ловушки определить изменение квантового выхода генерации АФК при связывании ATR с ретиналь-связывающими белками.
Определить характеристики возбужденного триплетного состояния ATR методом лазерного импульсного фотолиза.
Исследовать кинетические характеристики тушения триплетного состояния ретиналя молекулярным кислородом в различных средах – водных растворах, липосомах, в состоянии, связанном с альбумином и специфическим ретиналь-переносящим белком из межфоторецепторного матрикса.
Определить соотношение между связанным (образующим Шиффовы основания) и свободным ATR в фоторецепторном диске при полном обесцвечивании родопсина.
Научная новизна
Впервые установлено, что эффективность генерации АФК снижается при связывании ATR с альбумином. Установлено, что при эквимолярном соотношении альбумина и ATR эффективность уменьшается в четыре раза.
Впервые изучены кинетические параметры тушения триплетного состояния ретиналя молекулярным кислородом в водных растворах, липосомах и в состоянии, связанном с ретиналь-связывающими белками. Показано, что скорость процесса тушения возбужденного триплетного состояния кислородом при связывании ATR с альбумином уменьшается в 8 раза, а в случае МРПБ - примерно в 50 раз.
Показано, что при связывании с фосфолипидными липосомами или фоторецепторными мембранами ATR локализуется на поверхности, а не внутри липидного бислоя. Определены константы связывания ретиналя с аминогруппами липидов в фоторецепторных мембранах. Установлено, что в фоторецепторных мембранах при нативном соотношении ATR и родопсина 70% ATR связывается с аминогруппами компонентов фоторецепторных мембран, и только 30% находится в свободном состоянии.
Научная и практическая ценность работы
Результаты диссертации имеют фундаментальное значение для понимания механизмов фотоповреждения сетчатки глаза и разработки средств защиты от него.
Основные положения, выносимые на защиту:
Связывание ATR с ретиналь-переносящими белками. Эффект экранирования ATR ретиналь-переносящими белками, приводящий к уменьшению квантового выхода активных форм кислорода.
Кинетические характеристики процесса тушения возбужденного триплетного состояния ATR молекулярным кислородом в различных средах. Уменьшение константы скорости реакции тушения триплетного состояния ATR молекулярным кислородом при связывании ATR с альбумином и межфоторецепторным ретиналь-переносящим белком (МРПБ).
Соотношение свободного и связанного с аминогруппами ATR при полном обесцвечивании родопсина в фоторецепторной мембране.
Апробация работы:
Основные результаты работы были доложены на следующих всероссийских и международных конференциях: Аболтин, П.В. Протекторное действие ретиналь-связывающих белков при генерации активных форм кислорода ретиналем. / П. В. Аболтин, Т. Ф. Шевченко, П. П. Левин, Г. Р. Каламкаров // Международная научная конференция «Молекулярные, мембранные и клеточные основы функционирования биосистем». - Сборник статей. – Минск, Беларусь. 2010, 23-25 июня. - С. 23-25; Аболтин, П. В. Фотоиндуцированная генерация активных форм кислорода производными и конъюгатами ретиналя / П. В. Аболтин, Т. Ф. Шевченко, П. П. Левин, К. Б. Шумаев, Г. Р. Каламкаров // Всероссийская конференция с международным участием «Спектроскопия и томография электронного парамагнитного резонанса в химии и биологии». – Москва 2011, 6-10 октября. – С. 107; Аболтин, П.В. Ретиналь-белковые взаимодействия как фактор, предотвращающий фототоксическое действие ретиналя в сетчатке глаза. / Аболтин П.В., Шевченко Т.Ф., Левин П.П., Шумаев К.Б, Каламкаров Г.Р. // УI Съезд Российского фотобиологического общества. - Материалы съезда. - Пос. Шепси 2011, 15-22 сентября. С. 79. Каламкаров, Г. Р. Ретиналь-белковые взаимодействия как фактор, предотвращающий окислительный и нитрозитивный стресс в сетчатке глаза / Г. Р. Каламкаров, П. В. Аболтин, Т. Ф. Шевченко, П. П. Левин, Т. С. Константинова, А. Е. Бугрова, К. Б. Шумаев // Седьмая научно-практическая конференция с международным участием «Активные формы кислорода, оксид азота, антиоксиданты и здоровье человека». - Материалы конференции. – Смоленск 2011, 14-18 сентября. - С. 112-113. Аболтин, П.В. Фотоиндуцированная генерация активных форм кислорода производными и конъюгатами ретиналя / П. В. Аболтин, Т. Ф. Шевченко, П. П. Левин, К. Б. Шумаев, Г. Р. Каламкаров // Съезд биофизиков. - Материалы докладов. – Нижний Новгород 2012, 20-26 августа. - Симпозиум II. С.8.
Публикации
По материалам диссертации опубликовано 3 статьи в журналах, рекомендованных ВАК, и 5 тезисов докладов, опубликованных по материалам конференций.
Структура и объем диссертации
Диссертация состоит из введения, 3 глав, заключения и выводов. Общий объем работы - 85 страниц и включает 1 таблицу, 21 рисунок и список литературы из 149 наименований.
Основное содержание работы
В главе 1 – Обзор литературы – анализируются современные представления о фотоиндуцированных процессах в сетчатке глаза позвоночных. Рассмотрены молекулярные механизмы фотохимических повреждений, происходящих вследствие генерации свободных радикалов хромофорами сетчатки и пигментного эпителия.
В главе 2 – Материалы и методы – описаны методы выделения и очистки белков и фоторецепторных мембран из тканей глаза, методы получения липосом, определения АФК с использованием спиновых ловушек (ЭПР) и импульсного лазерного фотолиза для исследования спектрально-кинетических характеристик ретиналя.
В главе 3 - Результаты и обсуждение - приведены результаты исследования влияния связывания ATR с липидами и ретиналь-связывающими белками на его способность генерировать свободные радикалы.
3.1 Исследование ретиналь-белковых взаимодействий методом спиновой ловушки
Для регистрации активных форм кислорода применялся метод спиновой ловушки, в качестве которой использовался ДЕПМПО (5-диэтоксифосфорил-5-метил-1-пирролин-N-оксид).
Приведенные в данной главе эксперименты проводили в модельной системе, где в качестве ретиналь-связывающего белка использовали бычий сывороточный альбумин (BSA), который, как и МРПБ, находится в фоторецепторном матриксе и, вероятно, также может участвовать в транспорте ретиноидов. Кроме того, использование продажного препарата BSA существенно облегчало решение поставленных задач, поскольку выделить из глаз крупного рогатого скота достаточное для таких экспериментов количество МРПБ невозможно.
На рисунке 1 представлены спектры ЭПР аддуктов спиновой ловушки ДЕПМПО со свободными радикалами кислорода, возникающими при освещении (макс= 375 нм) ATR в 10% водном растворе этанола. Спектр представлен характерной для аддуктов ДЕПМПО сверхтонкой структурой, состоящей из 8 компонент (рис. 1, спектр 4). Для того, чтобы выявить, какие свободные радикалы кислорода могут привести к появлению такого спектра, моделировались базисные спектры аддуктов спиновой ловушки DEPMPO с супероксид-анион радикалом и гидроксильным радикалом (рис.1, спектры 1, 2, 3).
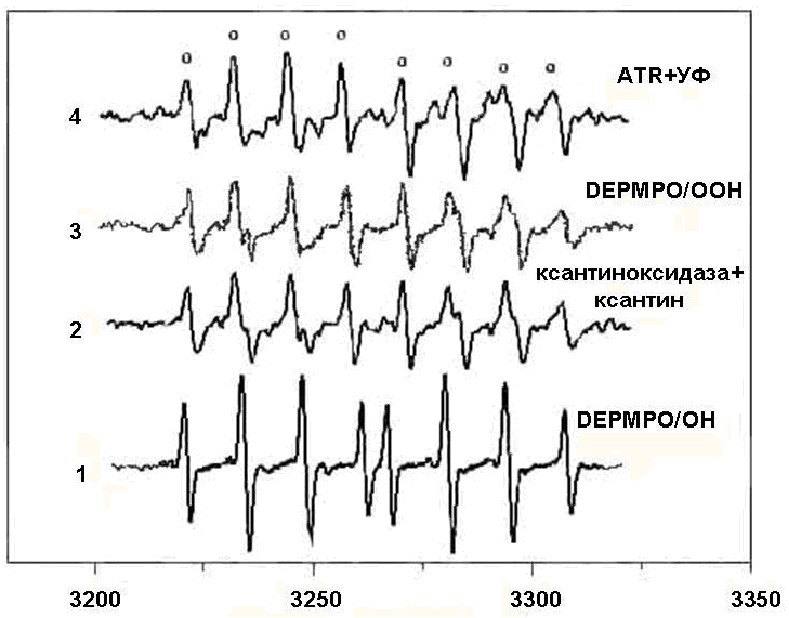 | Рис. 1. Спектры ЭПР аддуктов DEPMPO. 1 - спектр аддукта гидроксильного радикала; 2 - спектр аддуктов, возникающий в ферментативной реакции ксантин-ксантиноксидаза; 3 - разностный спектр, соответствующий сигналу ЭПР спинового аддукта супероксида; 4 - спектр аддуктов ДЕПМПО в среде, содержавшей ATR, после облучения светом. |
Генерацию гидроксильного радикала проводили в системе Fe2+/H2O2, т. е. в реакции Фентона:
Fe+2 + Н2О2 Fe+3 + OH• + OH- (1).
Супероксидный радикал (О2•) продуцировался ферментативно в системе, содержавшей ксантин и ксантиноксидазу. Реакционная среда содержала 1 ммоль/л ксантина и 0.2 ед/мл ксантиноксидазы. В случае этой ферментативной реакции регистрируемый спектр ЭПР является суперпозицией спектров аддуктов ДЕПМПО с О2• и гидроксильным радикалом (рисунок 1, спектр 2), поэтому для получения спектра, наиболее близкого сигналу ЭПР спинового аддукта ДЕПМПО/OOH (аддукт с супероксидом), рассматривался разностный спектр (рис. 1, спектр 3): 100% спектра 2 12,5% спектра 1. С помощью полученных спектров аддуктов супероксид анион-радикала и гидроксильного радикала с ДЕПМПО производилось разложение экспериментального спектра на компоненты. Было выяснено, что больший вклад в зарегистрированный сигнал вносили сигналы аддуктов cпиновой ловушки с супероксидом (на рис 1, спектр 4 - компоненты сверхтонкой структуры, отмеченные кружками). Как показано на рисунке, вклад аддукта(ов) других свободных радикалов в спектр 4 сравнительно невелик.
Таким образом, было показано, что спектр ЭПР, зарегистрированный при облучении системы, содержавшей АТR и ДЕПМПО, представляет собой суперпозицию следующих компонент: ~80% супероксид-анион радикала, ~15% гидроксильного радикала и ~5% радикала неизвестной природы.
В наших экспериментах нельзя исключить образование свободных радикалов, являющихся производными АТR. Например, может образоваться анион-радикал АТR:
(АТR* + e- АТR• ) (2)
а также катион-радикал молекулы АТR в результате одноэлектронного окисления молекулярным кислородом в реакции:
АТR* + О2 АТR•+ + О2• (3).
Образование супероксид-анион радикала возможно также в следующих реакциях:
АТR• + О2 АТR + О2• (4),
АТR* 3АТR (5),
3АТR + 3O2 1АТR + 1O2 (6),
1O2 + e О2• (7), где 3ATR – триплетное состояние ATR, 1ATR – синглетное состояние ATR. Прямо оценить квантовый выход образования свободных радикалов кислорода не представляется возможным, однако можно рассчитать относительную скорость их образования и, соответственно, относительный квантовый выход образования свободных радикалов по величине максимальной скорости нарастания интегральной интенсивности сигнала ЭПР при освещении. Интегральную интенсивность приближенно оценивали, пользуясь выражением Sпл = Iмакс(Hмакс)2, где Sпл - площадь под кривой поглощения, Iмакс - интенсивность линии, Hмакс - ширина линии. В качестве Iмакс мы рассматривали амплитуду третьей компоненты зарегистрированных сигналов ЭПР, так как именно эта компонента максимально совпадает с сигналом, соответствующим спиновому аддукту супероксид-анион радикала, вносящему основной вклад. Таким образом минимизируется вклад свободных радикалов, являющихся производными АТR. Кинетика образования АФК представлена на рис. 2. Оценивалась относительная скорость образования АФК при освещении для ATR, А2Е и ATR в присутствии BSA.
Относительные величины квантовых выходов генерации АФК представлены на рис. 3.
Квантовый выход для А2Е в полтора раза выше, чем для ATR. Этот факт хорошо согласуется с данными других авторов. Квантовый выход генерации АФК ATR значительно снижался при связывании BSA с ретиналем. Причем, эта величина оказалась пропорциональной относительной концентрации BSA.
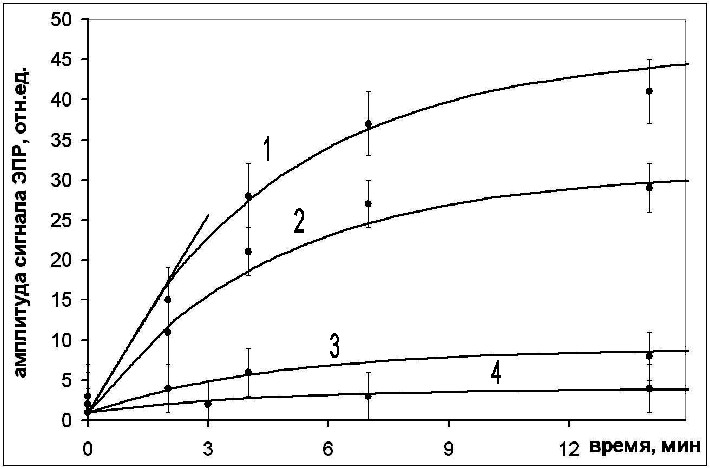 | Рис. 2. Зависимость амплитуды низкополевой компоненты спектра ЭПР аддукта DEPMPO, возникающего при освещении: 1 - А2Е, 2 - ATR, 3 - ATR в присутствии эквимолярного количества BSA, 4 - ATR в присутствии 4-кратного избытка BSA |
 | Рис. 3. Относительная величина квантового выхода генерации АФК при освещении растворов А2Е, ATR и ATR в комплексе с BSA в разных концентрациях. |
3.2 Исследование спектрально-кинетических характеристик триплетного состояния ATR
Во второй части работы методом лазерного импульсного фотолиза исследовалось взаимодействие ATR как с альбумином, так и с МРПБ. В разделе ставились те же задачи, что и в предыдущем – изучить влияние связывания ATR с ретиналь-связывающими белками на скорость тушения возбужденного триплетного состояния молекулярным кислородом. Тушение возбужденного состояния кислородом является одним из путей образования радикальных форм кислорода, хотя и не единственным. Донором электрона в сложной биологической системе может выступать практически любая группа с соответствующими донорными свойствами, например, ароматические аминокислотные остатки, сульфгидрильные группы или сам ретиналь, который при этом может окисляться. В модельной системе, в которой находится только ретиналь в водно-спиртовом растворе, донором электрона является, скорее всего, сам ретиналь, что показано и в литературе. Таким образом, несмотря на различие экспериментальных подходов (ЭПР и импульсный фотолиз), в разделах ставились, по существу, сходные задачи. Кроме того, и это крайне важно с биологической точки зрения, при использовании импульсного фотолиза удалось исследовать взаимодействие ATR не только с BSA, но и с МРПБ – именно того белка, который и осуществляет транспорт ретиналя и ретинола в нативной фоторецепторной клетке.
3.2.1 Спектрально-кинетические характеристики 3ATR в растворах
Исследовалось поведение ATR в метанольном растворе, водном растворе с небольшой добавкой метанола, в суспензии липосом, в водных растворах альбумина и водных растворах МРПБ.
Импульсное фотовозбуждение (1 нс, 337 нм) растворов ATR приводит за время лазерной вспышки к образованию триплетного состояния 3ATR, характеризующегося известным спектром поглощения с max около 470 нм (рис. 4, спектр 1).
Кинетика гибели 3ATR как в обескислороженном, так и в воздушнонасыщенном метанольном растворе подчиняется закону первого порядка. Соответствующие константы скорости (kT и kTо), а также рассчитанная из этих значений константа скорости тушения 3ATR молекулярным кислородом (kq) приведены в таблице 1.
 | Рис. 4. Дифференциальные спектры поглощения промежуточных продуктов, полученные при лазерном фотолизе метанольных растворов ATR (5х10-5 моль/л) через 0.5 (1) и 100 (2) мкс. |
При лазерном фотолизе ATR в метаноле (МеОН), помимо относительно короткоживущего триплет-триплетного поглощения, наблюдается также слабое долгоживущее наведенное поглощение с максимумом при max 400 нм (рис. 4, спектр 2). Выход этого промежуточного продукта не зависит от присутствия кислорода в растворе, что свидетельствует о том, что он образуется из синглетного электронно-возбужденного состояния и, по литературным данным, он представляет собой изомер или радикал ретиналя.
Таблица 1 Константы скорости гибели триплетного состояния полностью транс-ретиналя в различных обескислороженных (kT) и воздушнонасыщенных (kTo) средах и константа скорости его тушения молекулярным кислородом (kq).
| Среда | kT105, c-1 | kTo105, c-1 | kq109, л моль-1 с-1 |
| Липосомыa | 0.90 | 14 | 4.6d |
| H2Ob | 1.7 | 4.7 | 1.1d |
| МРПБc | 6.0e | 6.4e | ~0.1d |
| BSAc | 6.7e | 8.5e | 0.62d |
| MeOH | 0.70 | 110 | 5.0 |
| ФРМc | 0.89 | 13 | 4.3 d |
a 0.15 моль/л K-P буфер, pH 7.4, содержащий 810-3 моль/л фосфатидилхолина и 1% метанола по объему.
b 0.15 моль/л K-P буфер, pH 7.4, содержащий 1% метанола по объему.
c 0.14 моль/л NaCl, 0.05 моль/л трис-HCl, pH 7.4, содержащий 10-5 моль/л белка и 1% метанола по объему.
d Рассчитана, исходя из концентрации кислорода в воздушнонасыщенной воде, равной 2.8х10-4 моль/л.
e Для быстрой компоненты кинетики гибели 3ATR (kTs). Рассчитана, исходя из концентрации кислорода в воздушнонасыщенном MeOH, равной 2.210-3 моль/л.
3.2.2 Спектрально-кинетические характеристики 3ATR в липосомах.
Квантовый выход 3ATR суспензии липосом из фосфатидилхолина (л-ФХ) (810-3 моль/л ФХ) примерно в 5 раз ниже, чем в растворах метанола Значение констант в 100% метаноле и водной суспензии л-ФХ существенно выше, чем в водном растворе, где концентрация спирта не превышала 1%. Такие значения kq типичны для процесса тушения триплетных состояний органических молекул, обладающих умеренными электронно-донорными свойствами, кислородом в полярных растворителях. Процесс в этом случае может осуществляться по двум каналам: путем переноса энергии с образованием синглетного кислорода (в синглетном комплексе встречи, спин-статистический фактор 1/9) и за счет спиновой конверсии в триплетном комплексе с переносом заряда (спин-статистический фактор 1/3). Реализация такого механизма приводит к тому, что выход синглетного кислорода при тушении 3ATR, близкий к 100% в неполярных растворителях, существенно уменьшается при переходе к полярным растворителям. Однако высокое значение kq в липосомах обусловлено, по-видимому, не только наличием двух каналов процесса, но также высокой концентрацией О2 в липидном бислое (выше, чем в водном объеме) и его высокой латеральной подвижностью. При лазерном фотолизе ATR в л-ФХ наблюдается также заметное долгоживущее наведенное поглощение в области 400–500 нм, природа которого обсуждалась выше.
3.2.3 Спектрально-кинетические характеристики триплетного состояния ATR в белковых растворах.
При фотолизе ATR в водных растворах BSA и МРПБ (max около 460 нм) наблюдается триплетное состояние 3ATR с квантовым выходом примерно в 2 раза ниже, чем в л-ФХ. При этом в присутствии белков происходит, с одной стороны ускорение, а с другой стороны - замедление гибели 3ATR в обескислороженных растворах. Аппроксимация кинетических кривых по двухэкспоненциальному закону дает «быструю» и «медленную» компоненты приблизительно с одинаковыми вкладами и различающимися в 10 раз константами скорости (константы скорости «быстрой» компоненты kT приведены в таблице 1) (рис. 5, кривые 1).
При переходе к воздушнонасыщенным растворам наблюдается ускорение «быстрой» компоненты и возрастание ее вклада до 70 и 80% для МРПБ и BSA, соответственно (константы скорости «быстрой» компоненты kTo, а также соответствующие значения констант скорости тушения этих 3ATR молекулярным кислородом kq приведены в таблице 1) (рис. 5, кривые 2). Константа скорости гибели «медленной» компоненты практически не изменяется. Таким образом, как в концентрированных, так и в разбавленных белковых растворах наблюдается экранирование 3ATR по отношению к молекулярному кислороду.
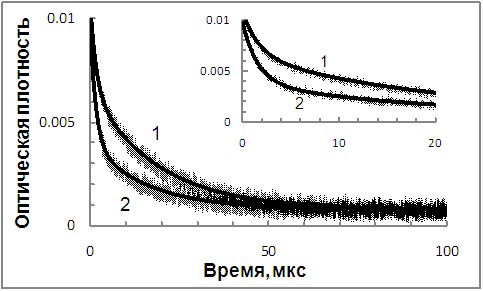 | Рис. 5. Кинетика гибели промежуточных продуктов, поглощающих при =460 нм, полученная при лазерном фотолизе обескислороженных (1) и воздушнонасыщенных (2) водных (0.15 моль/л K-P буфер, рН 7.4) растворов ATR (1.510-5моль/л) в присутствии 3.510-3 моль/л ФХ. На экспериментальные кривые наложены кривые, полученные при аппроксимации по одноэкспоненциальному закону с незначительной постоянной составляющей |
Существенное экранирование ретиналя от молекулярного кислорода при его связывании с белками необходимо учитывать при рассмотрении окислительных процессов в биосистемах. Так, то, что ретиналь в зрительной клетке и в пигментном эпителии постоянно находится в состоянии, связанном с ретиналь-переносящими белками, может быть важным протекторным фактором, предотвращающим образование критических концентраций активных форм кислорода.
При фотолизе ATR в водных белковых растворах, помимо триплетного состояния, также наблюдается образование незначительного количества более долгоживущих промежуточных продуктов (около 10% по поглощению при 460 нм относительно поглощения 3ATR), характеризующихся полосой поглощения с максимумом около 400 нм и имеющих, по-видимому, радикальную природу. Введение в белковые растворы липосом приводит к резкому ускорению гибели 3ATR в воздушнонасыщенных растворах. Кинетика гибели 3ATR подчиняется одноэкспоненциальному закону с константой скорости, близкой к значению kTo в л-ФХ. Однако в обескислороженных растворах кинетика гибели 3ATR остается двухэкспоненциальной. По мере увеличения концентрации ФХ «быстрая» компонента замедляется (рис. 6), а ее вклад возрастает.
 | Рис. 6. Зависимость константы скорости гибели «быстрой» компоненты 3ATR в растворах МРПБ (1) и БСА (2) (10-5 моль/л) от концентрации ФХ. |
Константа скорости «медленной» компоненты при этом практически не изменяется. Таким образом, ATR остается локализованным в белках. Только при большой концентрации липидов ( 5 ммоль/л) кинетика гибели 3ATR совпадает с кинетикой гибели 3ATR в л-ФХ.
В целом, можно сделать вывод о том, что ретиналь при наличии в системе ретиналь-переносящих белков (как альбумина, так и специфического МРПБ из фоторецепторного матрикса) всегда будет переходить из липидного бислоя в карманы белка. Это, в свою очередь, значительно снижает вероятность образования под действием света активных форм кислорода и, таким образом, оказывает значительное протекторное действие, защищающее клеточные компоненты от вредного действия света.
3.2.4 Образование Шиффовых оснований ATR в липосомах
Концентрация родопсина в фоторецепторной клетке составляет 3-4 ммоль/л, и поэтому формально при интенсивном освещении, приводящем к полному обесцвечиванию родопсина, в фоторецепторной клетке могло бы образовываться до 4 ммоль/л ATR. Под обесцвечиванием в биологической литературе понимается изменение окраски родопсина (макс = 500 нм) при освещении, приводящем к распаду родопсина на белок и ATR (макс=380 нм). Ясно, что при такой огромной концентрации, несмотря на невысокий квантовый выход генерации АФК, ATR оказывал бы катастрофическое повреждающее действие. В нативной клетке этого не происходит из-за высокой активности ретинолдегидрогеназы – фермента, катализирующего переход ATR в полностью транс-ретинол. Если бы этого не происходило, высокая химическая активность ретиналя приводила бы к образованию Шиффовых оснований с аминогруппами, которые в избытке находятся в клетке. Именно высокая активность фермента является причиной того, что даже при очень высоких освещенностях значительные количества ретиналя не удается обнаружить в фоторецепторной клетке. Однако восстановление ретиналя в ретинол происходит в цитоплазме клетки, а высвобождается ретиналь во внутридисковое пространство, откуда транспортируется в цитоплазму. Во внутридисковом пространстве ATR образует Шиффовы основания с аминогруппами, прежде всего, с аминогруппами ФЭА, концентрация которого в фоторецепторе оставляет 10 ммоль/л. Как известно, Шиффовы основания ATR не образуют триплетного состояния и поэтому не являются активными генераторами АФК. В связи с эти мы поставили задачу определить, какая часть ATR во внутридисковом пространстве образует Шиффовы основания и не обладает фототоксическими свойствами. Поскольку Шиффовы основания не образуют триплетов, по концентрации возбужденного триплетного состояния можно определить константы равновесия Шиффовых оснований как в липосомах из ФЭА (в таблице 1 – липосомы), так и в нативных, содержащих родопсин, фоторецепторных мембранах (ФРМ). В данной серии экспериментов к суспензии л-ФХ добавляли суспензию липосом из ФЭА. Введение в раствор ФЭА приводит к уменьшению наблюдаемого выхода 3ATR, но не влияет на кинетику его гибели. ФЭА образует Шиффово основание с ATR, которое не обладает карбонильной группой, обеспечивающей эффективное спин-орбитальное взаимодействие в первом синглетном электронно-возбужденном состоянии и, соответственно, не образует триплетное состояние при прямом фотолизе. Таким образом, наблюдаемый относительный выход 3ATR может служить мерой доли ATR, находящейся в виде Шиффова основания (ШО). Соответствующая концентрационная зависимость (рис. 7) указывает на существование равновесия (7):
ATR + ФЭА ШО (7).
 | Рис.7. Зависимость интенсивности поглощения 3ATR при 470 нм, зарегистрированная через 20 нс после лазерного импульса в присутствии л-ФХ (410-3 моль/л ФХ) от концентрации ФЭА в системе. |
Зависимость на рис.7 хорошо описывается соответствующим выражением
[3ATR]набл = [3ATR]0/(1 + K[ФЭА]) (8),
где [3ATR]набл наблюдаемая начальная концентрация 3ATR в присутствии ФЭА, [3ATR]0 начальная концентрация 3ATR, наблюдавшаяся в отсутствие ФЭА, а К = 8,0 (мольная доля)-1 – константа равновесия (7).
Выражение (8) получается при соответствующем решении кинетической схемы (7) при условии [ATR] << [ФЭА], которое хорошо выполняется в данных экспериментальных условиях.
3.2.5 Образование Шиффовых оснований ATR в ФРМ
В следующей серии экспериментов использовали суспензию ФРМ, выделенных из зрительных клеток и засвеченных до полного распада комплекса опсина с 11-цис-ретиналем. Фотолиз такого образца приводит к образованию 3ATR, спектрально-кинетические характеристики которого практически полностью совпадают с аналогичными параметрами в л-ФХ (рис. 8, спектр 2; табл. 1).
 | Рис. 8. Дифференциальные спектры поглощения промежуточных продуктов, полученные при лазерном фотолизе воздушнонасыщенных растворов ATR (10-5 моль/л) в присутствии л-ФХ (4. 10-3 моль/л ФХ) и в системе с ФРМ через 0.02 (2) и 5 (4) мкс после лазерного импульса. |
Однако, квантовый выход 3ATR в ФРМ составляет только 30% от соответствующего выхода, получающегося при фотолизе ATR, добавленного в раствор л-ФХ в эквимолярном количестве. Таким образом, на основании полученных данных по равновесию (7) можно сделать вывод, что 70% образующегося при полном обесцвечивании родопсина ATR образует Шиффовы основания и не генерирует АФК.
Важно отметить, что определить константы равновесия ATR и аминогрупп (Шиффовых оснований) можно по сдвигу максимума поглощения, однако, в нативной фоторецепторной мембране, содержащей родопсин, это сделать невозможно из-за наличия долгоживущих продуктов фотолиза родопсина, максимум поглощения которых отличается от ATR из-за его локализации в «белковых карманах» родопсина. Поэтому можно полагать, что определение равновесия по квантовому выходу 3ATR является более адекватным методом.
Выводы.
1. С использованием метода спиновой ловушки показано, что связывание полностью транс-ретиналя с альбумином приводит к значительному снижению относительного квантового выхода фотогенерации супероксид анион-радикала.
2. Методом наносекундного лазерного фотолиза изучена кинетика гибели возбужденного триплетного состояния ATR в воде, в водных растворах липосом и ретиналь-связывающих белков, а также в фоторецепторных мембранах. Установлена природа мест локализации АТR.
3. В белках обнаружено как минимум два места локализации АТR, которые существенно (до 2 порядков) различаются по степени экранирования триплетного состояния ATR от молекулярного кислорода.
4. В результате прямых кинетических измерений установлено, что константа скорости тушения возбужденного триплетного состояния ATR молекулярным кислородом при его связывании с альбумином и ретиналь-переносящим белком уменьшается в 10 и 50 раз, соответственно.
5. По выходу 3ATR определены константы равновесия обратимого процесса связывания ATR с аминогруппами (образование Шиффовых оснований) компонентов фоторецепторной мембраны. Показано, что при полном обесцвечивании родопсина 70% ATR образуют Шиффовы основания с аминогруппами и не генерируют АФК.
Список публикаций по теме работы:
1. Левин, П.П. Исследование кинетики тушения триплетного состояния полностью-транс-ретиналя кислородом в водных растворах альбумина и липосом методом лазерного фотолиза / П.П. Левин, П.В. Аболтин, Т.Ф. Шевченко, Г.Р. Каламкаров. // Химия высоких энергий. - 2010. - Т.44. - №6. - С. 555-559.
2. Левин, П.П. Исследование спектрально-кинетических характеристик триплетного состояния полностью транс-ретиналя в водных растворах ретиналь-связывающих белков и липосом методом лазерного фотолиза. / П.П. Левин, П.В. Аболтин, Т.С. Константинова, Т.Ф. Шевченко, Г.Р. Каламкаров. // Химия высоких энергий. – 2013. - Т.47. - № 3. - С. 1–4.
3. Аболтин, П.В. Фотоиндуцированная генерация активных форм кислорода производными и конъюгатами ретиналя / П.В. Аболтин, Т.Ф. Шевченко, К.Б. Шумаев, Г.P. Каламкаpов. // Биофизика. – 2013. - Т. 58. - вып. 2. - С. 246–251.
4. Аболтин, П.В. Протекторное действие ретиналь-связывающих белков при генерации активных форм кислорода ретиналем. / П. В. Аболтин, Т. Ф. Шевченко, П. П. Левин, Г. Р. Каламкаров // Международная научная конференция «Молекулярные, мембранные и клеточные основы функционирования биосистем». - Сборник статей. – Минск, Беларусь. 2010, 23-25 июня. - С. 23-25.
5. Аболтин, П. В. Фотоиндуцированная генерация активных форм кислорода производными и конъюгатами ретиналя / П. В. Аболтин, Т. Ф. Шевченко, П. П. Левин, К. Б. Шумаев, Г. Р. Каламкаров // Всероссийская конференция с международным участием «Спектроскопия и томография электронного парамагнитного резонанса в химии и биологии». – Москва 2011, 6-10 октября. – С. 107.
6. Каламкаров, Г. Р. Ретиналь-белковые взаимодействия как фактор, предотвращающий окислительный и нитрозитивный стресс в сетчатке глаза / Г. Р. Каламкаров, П. В. Аболтин, Т. Ф. Шевченко, П. П. Левин, Т. С. Константинова, А. Е. Бугрова, К. Б. Шумаев // Седьмая научно-практическая конференция с международным участием «Активные формы кислорода, оксид азота, антиоксиданты и здоровье человека». - Материалы конференции. – Смоленск 2011, 14-18 сентября. - С. 112-113.
7. Аболтин, П.В. Ретиналь-белковые взаимодействия как фактор, предотвращающий фототоксическое действие ретиналя в сетчатке глаза. / Аболтин П.В., Шевченко Т.Ф., Левин П.П., Шумаев К.Б, Каламкаров Г.Р. // УI Съезд Российского фотобиологического общества. - Материалы съезда. - Пос. Шепси 2011, 15-22 сентября. С. 79.
8. Аболтин, П.В. Фотоиндуцированная генерация активных форм кислорода производными и конъюгатами ретиналя / П. В. Аболтин, Т. Ф. Шевченко, П. П. Левин, К. Б. Шумаев, Г. Р. Каламкаров // Съезд биофизиков. - Материалы докладов. – Нижний Новгород 2012, 20-26 августа. - Симпозиум II. С.8.