

Александрович лесная растительность бассейна р. илыч в верхнем и среднем течении (в границах печоро-илычского заповедника)
На правах рукописи
Дубровский Юрий Александрович
Лесная растительность бассейна р. Илыч
в верхнем и среднем течении (в границах Печоро-Илычского заповедника)
03.00.05 – ботаника
АВТОРЕФЕРАТ
диссертации на соискание учёной степени
кандидата биологических наук
Сыктывкар 2009
Работа выполнена в Учреждении Российской академии наук Институте биологии Коми научного центра Уральского отделения Российской академии наук
Научный руководитель: доктор биологических наук
Дёгтева Светлана Владимировна
Официальные оппоненты: доктор биологических наук
Нешатаева Валентина Юрьевна
доктор биологических наук, профессор
Бобкова Капитолина Степановна
Ведущая организация: Петрозаводский государственный университет
Защита состоится 17 декабря 2009 года в 17 часов на заседании диссертационного совета Д 004.007.01 в Учреждении Российской академии наук Институте биологии Коми НЦ УрО РАН по адресу: 167982, г. Сыктывкар, ул. Коммунистическая, д. 28.
Факс: (8212) 24-01-63
E-mail: [email protected]
Сайт: http://www.ib.komisc.ru
С диссертацией можно ознакомиться в библиотеке Коми научного центра Уральского отделения Российской АН по адресу: 167982, г. Сыктывкар, ул. Коммунистическая, д. 24.
Автореферат разослан «____» ноября 2009 г.
Учёный секретарь
диссертационного совета
доктор биологических наук А.Г. Кудяшева
Общая характеристика работы
Актуальность темы. Интенсивное лесопользование в таёжной зоне привело к существенной трансформации лесных экосистем. Массивы мало нарушенных и девственных лесов в Европейской части России сегодня крайне немногочисленны и занимают подчинённые площади в составе лесного фонда (Леса…, 1999). В Республике Коми девственная темнохвойная тайга сохранилась преимущественно в предгорьях и на западном макросклоне Уральских гор в пределах двух особо охраняемых природных территорий федерального значения – Печоро-Илычского биосферного заповедника и национального парка «Югыд ва». Растительный покров этих резерватов, занимающих в общей сложности площадь порядка 3 млн. га, изучен ещё далеко не достаточно. Исследователи, работавшие на территории заповедника в разное время (Говорухин, 1929; Корчагин, 1940; Ланина, 1963; Горчаковский, 1966, 1968; Взаимосвязи компонентов…, 1980; Непомилуева, 1992; Флора и растительность…, 1997; Сукцессионный…, 2006) в основном делали акцент на изучении коренных старовозрастных темнохвойных лесов, развивающихся в режиме спонтанной динамики. Естественное развитие климаксовых лесных сообществ нарушается такими катастрофическими природными факторами, как пожары и ветровалы, в результате которых формируются массивы производных берёзовых и осиновых лесов. Вторичные мелколиственные леса, которые в настоящее время занимают в резервате заметные площади, остались малоизученными. Недостаточны сведения о лесных сообществах горной ландшафтной зоны, особенно о разнообразии редколесий подгольцового пояса. Необходимо было продолжить углублённое исследование отдельных лесных формаций и типов растительности.
Целью работы было выявление видового и ценотического разнообразия растительного мира лесов верхнего и среднего течения реки Илыч (в границах Печоро-Илычского биосферного заповедника), изучение их динамики. Для достижения поставленной цели необходимо было решить следующие задачи:
- выявить разнообразие ценофлор различных лесных формаций и их групп (светлохвойных – сосняки, лиственничники, темнохвойных – ельники, пихтарники, кедровые леса, мелколиственных лесов – березняки, осинники);
- изучить типологическую структуру лесных сообществ, выполнить их классификацию в рамках эколого-фитоценотического подхода;
- исследовать особенности трансформации структуры древостоев, изменения видового состава, ценотической роли растений травяно-кустарничкового яруса и мохового покрова в процессе демутационных сукцессий на примере сообществ березняков и ельников зеленомошной группы типов.
Научная новизна. Впервые выявлен и проанализирован видовой состав сосудистых растений основных лесных формаций в пределах верхнего и среднего течения реки Илыч, зарегистрировано 242 вида из 163 родов и 60 семейств. Проведён сравнительный анализ ценофлор 8 формаций. Показано, что наибольшим разнообразием характеризуются еловые леса. Составлена классификация лесов заповедника с использованием эколого-фитоценотического подхода. Дана геоботаническая характеристика 80 ассоциаций, 35 из которых являются новыми для территории Печоро-Илычского заповедника. Впервые для Республики Коми установлены закономерности изменения состава и структуры древостоев и нижних ярусов сообществ в процессе формирования лиственных насаждений на гарях и их смены темнохвойными фитоценозами в условиях заповедного режима территории. Показано, что восстановление древесного, травяно-кустарничкового и мохово-лишайникового ярусов идет более интенсивно, чем на вырубках.
Научная и практическая значимость работы. Результаты, полученные при анализе ценофлор и классификации лесных фитоценозов Печоро-Илычского заповедника, могут использоваться для мониторинга состояния экосистем территории. Обработанные данные могут найти применение при составлении карт растительности резервата и региона. Результаты исследований могут применяться в процессе чтения курсов лекции по различным дисциплинам ботаники и экологии.
Личный вклад. В основу диссертации положены геоботанические описания, выполненные автором в 2004–2009 гг. Автором была проведена обработка полученных данных с применением математических методов. Самостоятельно проведён флористический анализ общего списка видов а также формационный анализ видового состава изученных лесных формаций. Составлена схема классификации лесной растительности северной части резервата. Дана оценка разнообразия лесов на видовом и ценотическом уровнях. Построены схемы динамических смен растительного покрова в процессе реализации сукцессий.
Апробация. Материалы диссертационной работы представлены на молодёжных, всероссийских и международных научных конференциях: «Актуальные проблемы биологии и экологии» (Сыктывкар, 2005-2009), «Актуальные проблемы регионального экологического мониторинга: научный и образовательный аспекты» (Киров, 2006), «Актуальные проблемы геоботаники. III Всероссийская школа-конференция» (Петрозаводск, 2007), «I(III) Всероссийская молодёжная научно-практическая конференция ботаников в Новосибирске» (Новосибирск, 2007), «Принципы и способы сохранения биоразнообразия: III Всероссийская научная конференция» (Пущино, 2008), «Международная научная конференция, посвящённая 135-летию со дня рождения И.И. Спрыгина» (Пенза, 2008), «Международная научная конференция: Мониторинг и оценка состояния растительного мира» (Минск, 2008), «Международная молодёжная конференция «Молодёжь в науке – 2009» (Минск, 2009).
Публикации. По результатам исследований, проведённых в рамках подготовки диссертации, опубликовано 16 работ, в том числе одна статья в «Ботаническом журнале» (список ВАК).
Структура и объём диссертации. Текст диссертации изложен на 250 страницах, включая 19 таблиц и 8 рисунков. Диссертация содержит 13 приложений на 70 страницах.
Содержание работы
Глава 1. История изучения растительного покрова Печоро-Илычского заповедника.
В главе с привлечением доступного литературного материала рассматривается история изучения растительности Печоро-Илычского заповедника. Показано, что первые данные о растительности резервата были получены в XIX веке (Keyserling, 1846), а с начала прошлого столетия началось планомерное изучение экосистем территории (Поле, 1915; Нат, 1915; Говорухин, 1929; Самбук, 1930; Корчагин, 1940; Ланина, 1940, 1963; Леонтьев, 1963; Боч, Василевич, 1976, 1980; Взаимосвязи…, 1980;). В наиболее полной на сегодняшний день классификационной сводке А.А. Корчагина (1940) приводится 122 растительные ассоциации. Кроме исследований, посвящённых общей структуре растительного покрова резервата, на территории заповедника проводились изучение отдельных типов растительности и растительных формаций. Специалисты изучали кедровые леса (Ланина, 1963), сосновые (Взаимосвязи…, 1980) и еловые (Леонтьев, 1963) насаждения, пойменные луга (Говорухин, 1929б; Самбук В.Ф., 1930; Самбук С.Г., 1979) и болота (Боч, Василевич, 1976, 1980). В 80-х годах начались комплексные исследования флоры и растительности заповедника, проводимые сотрудниками Института биологии Коми научного центра Уральского отделения РАН (Непомилуева, 1992; Дёгтева, 1992, 2005; Лавренко, Улле, Сердитов, 1995; Флора и растительность…, 1997; Улле, 2005; Дубровский, 2007). Активные работы на территории резервата проводят специалисты других научных организаций РАН (Оценка и сохранение…, 2000; Восточно-европейские..., 2004; Харитонова, 2005, 2007; Сукцессионный…, 2006; Биоразнообразие…, 2007; Горчаковский, 2007). Тем не менее, имеющиеся данные о флоре и растительности не могут быть признаны полными, а исследования по многим направлениям необходимо было продолжить.
Глава 2. Физико-географическая характеристика района
В главе приведены основные сведения о климате, рельефе и геологическом строении территории, почвах и растительности резервата, расположенного в восточной части Республики Коми (Варсанофьева, 1940; Атлас, 1964; Почвы…, 1972; Лавренко, Улле, Сердитов, 1995; Земля девственных…, 2001).
Глава 3. Материалы и методы
Материал для написания диссертационной работы был собран совместно со Светланой Владимировной Дёгтевой в ходе пяти полевых сезонов (2004 –2009 гг.), и включает в себя 468 геоботанических описаний. В обработке также были задействованы около 70 геоботанических описаний лесов, выполненных С.В. Дёгтевой в 1989 г. в окрестностях кордона Егра-Ляга.
При выполнении геоботанических описаний использованы методики, которые являются общепринятыми в геоботанике и лесной типологии (Анучин, 1952; Сукачёв и др., 1957; Полевая геоботаника, 1964; Нешатаев, 1987; Ипатов, 1998). Для определения экологических параметров увлажнения и богатства почв использовали экологические шкалы Л.Г. Раменского (Экологическая оценка…, 1956), при работе с факторами освещённости и кислотности почвы использовали экологические шкалы Д.Н. Цыганова (1976, 1983). При описании растительности учитывали характеристики древостоя, подроста, подлеска, травяно-кустарничкового яруса, мохового и лишайникового покрова.
Для учёта количественных характеристик видов травяно-кустарничкового яруса и напочвенного покрова использовали методику, предложенную В.С. Ипатовым (1998). Степень доминирования вида оценивали непосредственно на всей пробной площади (400 м2), с использованием шкалы господства, (Ипатов, 1964, 1998). В процессе анализа геоботанического материала, для каждой формации определяли ценотическую роль видов травяно-кустарничкового яруса методом расчёта коэффициента участия (КУ), предложенного В.С. Ипатовым (1998), который учитывает данные о встречаемости и обилии видов:
![]() , где
, где
n – число описаний, m – встречаемость вида, a – ранг вида.
В целях оценки биоразнообразия использовали показатель -разнообразия. Определяли видовое богатство (общее число видов сосудистых растений) для синтаксонов рангом не ниже ассоциации (Оценка и сохранение…, 2000).
В дальнейшем, собранные материалы с использованием программ Turboveg и Microsoft Excel были обобщены в виде электронных баз данных, включающих в себя сводные геоботанические таблицы для изученных лесных формаций и выделенных нами ассоциаций, а также списки видового состава сосудистых растений. Формационный анализ флористических комплексов сосудистых растений выполнен с применением методики Н.В. Козловской (1978).
Списки видового состава документированы гербарными сборами, выполненными автором, С.В. Дёгтевой, В.А. Каневым, И.А. Плотниковой и хранящимися в гербарии Института биологии Коми НЦ УрО РАН (SYKO), и в гербарии Сыктывкарского государственного университета. Определение сосудистых растений выполнено с использованием монографии «Флора Северо-Востока европейской части СССР» (1974, 1976, 1977). По этой же сводке принята система типов ареалов видов. Определение мохообразных выполнила Т.П. Шубина, лишайников – Т.Н. Пыстина и Н.А. Семёнова. Анализ жизненных форм проводили по системе И.Г. Серебрякова (1962). Для определения сходства флористического состава выделенных лесных формаций использован коэффициент Стугрена-Радулеску (Шмидт, 1984). С целью определения экологической приуроченности описаний была проведена ординация в программном продукте PC-ORD методом. Использовали метод DCA - безтрендовый анализ соответствий (Hill, 1979; Hill, Gauch, 1980) с применением шкал Раменского и Цыганова.
При классификации полученных описаний лесной растительности использовали эколого-фитоценотический подход (Александрова, 1969; Сукачёв, 1975; Рысин, Савельева, 2002) и метод классификации массивов данных по Варду (Ward, 1963; Шитиков, Розенберг, Зинченко, 2003). При выделении ассоциаций помимо эдификаторов и доминантов принимали во внимание участие в формировании сообществ видов разных эколого-ценотических групп, выделенных для бассейна верхней и средней Печоры (Новаковский, Дегтева 2008; Дёгтева, Новаковский, 2009), а также особенности структуры сообществ, связанные с условиями экотопов. Ассоциации в случае необходимости подразделяли на субассоциации и варианты. Субассоциации выделяли с учетом присутствия индикаторных видов, отражающих специфику экологических условий, а также на основании содоминирования видов, варианты – по доминированию какого-либо вида эколого-ценотической группы, типичной для ассоциации. Ценотическое разнообразие изученных лесных формаций оценивали по числу синтаксонов разного ранга (групп типов леса, ассоциаций).
Глава 4. Флористический анализ видового состава лесов исследованной территории
К ведущим семействам относятся Asteraceae (33 вида), Poaceae (32), Cyperaceae (24), Ranunculaceae и Rosaceae (по 15 видов) и др. Преобладающие по числу видов семейства объединяют 171 вид или 58.6 % от общего числа таксонов. Данный спектр в целом совпадает с перечнями ведущих семейств как для всей зоны тайги (Флора…, 1987), так и для отдельных лесных формаций Республики Коми (Мартыненко, 1990; Дёгтева 1998; Ценотическая…, 2001; Флора…, 2007). Из особенностей стоит отметить уменьшение по сравнению с флорой всей зоны тайги значимости семейства Brassicaceae, что объясняется малым участием адвентивных видов в лесных экосистемах региона. Второй особенностью десятки ведущих семейств является повышение ранга сем. Salicaceae. Более значительную роль, чем во флоре региона, в лесных сообществах резервата играют также семейства Orchidaceae и Apiaceae. Более трети семейств являются одновидовыми, что отражает общую закономерность, обусловленную относительной молодостью флоры территории европейского севера России.
С фитогеографической точки зрения флору изученных лесных сообществ можно охарактеризовать как бореальную (204 вида или 69.9 %). В долготном спектре наибольшую долю (39 %) составляют виды (114), относящиеся к евразиатской группе. Наряду с видами, имеющими обширные ареалы, во флоре исследованных сообществ отмечены эндемичные растения, встречающиеся только в пределах Уральской горной страны – Lagotis uralensis и Anemonastrum biarmiense.
По жизненной форме абсолютное большинство растений, характерных для лесов заповедника (236 видов или 80.8 %), относится к травам. Анализ отношения видов, произрастающих в лесах, к фактору общего богатства почв показывает, что наибольшее их число (130 или 44.8 %) является олигомезотрофами. По отношению к фактору увлажнения наибольшую долю (50.7 % или 148 видов) в списке составляют мезофиты (Diplazium sibiricum, Pyrola minor, Vaccinium myrtillus и др.), предпочитающие нормально увлажненные хорошо дренированные почвы.
При анализе ценотической приуроченности видов, было выявлено, что среди выделенных групп видов наибольшим разнообразием отличаются долинные эколого-ценотические группы, растения которых предпочитают наиболее благоприятные с точки зрения факторов увлажнения и богатства почвы экотопы долин крупных и малых водотоков (39.7 % – 110 видов). В составе изученных лесных сообществ присутствуют представители 17 эколого-ценотических групп, что говорит о широкой области экологического пространства, занимаемой сообществами данного типа растительности. Леса в пределах бассейна среднего и верхнего течения реки Илыч являются местообитаниями 23 видов сосудистых растений, внесённых в список охраняемых видов Республики Коми (2008).
Таблица 1
Таксономический анализ видового состава сосудистых растений различных лесных формаций
| Группа формаций, формация | Число | Родовой коэффициент | Число | Доля ведущих семейств (% от общего числа видов) | |||
| Видов | Родов | Семейств | ведущих семейств | крупных родов | |||
| Темнохвойные леса | 229 | 140 | 54 | 1.64 | 16 | 7 | 71.7 |
| Ельники | 223 | 139 | 54 | 1.6 | 16 | 7 | 72.3 |
| Пихтарники | 106 | 75 | 38 | 1.41 | 4 | - | 34 |
| Кедровники | 51 | 38 | 26 | 1.21 | 1 | - | 15.2 |
| Светлохвойные леса | |||||||
| Лиственничники | 59 | 47 | 28 | 1.19 | 2 | 1 | 18.9 |
| Сосняки | 46 | 33 | 21 | 1.39 | 2 | 1 | 34.8 |
| Лиственные леса | 239 | 143 | 53 | 1.68 | 17 | 5 | 52.9 |
| Березняки | 180 | 116 | 47 | 1.57 | 14 | 5 | 69.8 |
| Горные берёзовые редколесья и криволесья | 137 | 89 | 40 | 1.54 | 8 | 2 | 54 |
| Осинники | 112 | 85 | 38 | 1.32 | 6 | 1 | 50 |
При формационных исследованиях (табл. 1) показано, что наибольшее богатство характерно для ценофлоры еловых лесов. Отрицательное значение коэффициента Стугрена-Радулеску (табл. 2), полученное при сравнении флористических списков ельников и березняков предгорных ландшафтов северной части заповедника, свидетельствует об их сходстве. Данный факт объясняется вторичным характером лиственных насаждений, которые в пределах исследованного района формируются в результате катастрофических воздействий на месте темнохвойных лесов. В таких случаях состав травяно-кустарничкового яруса остаётся в целом неизменным - меняется лишь роль отдельных видов. Наибольшее различие флористических списков (величина коэффициента Стугрена-Радулеску +0.32) обнаруживается при сравнении горных берёзовых редколесий и осинников.
Таблица 2
Матрица коэффициентов Стугрена-Радулеску, рассчитанных при сравнении флористических списков изученных формаций
| Формация | Пихтарники | Березняки | Горные берёзовые редколесья | Осинники |
| Ельники | 0.14 | –0.13 | 0.23 | 0.07 |
| Пихтарники | - | 0.08 | 0.13 | –0.06 |
| Березняки | - | - | 0.26 | 0.04 |
| Горные берёзовые редколесья | - | - | - | 0.32 |
Примечание: положительные значения коэффициента свидетельствуют о различии сравниваемых списков, отрицательные – о сходстве.
Основу флористических комплексов этих формаций составляют представители разных географических и эколого-ценотических групп.
Таблица 3
Значения коэффициентов участия наиболее ценотически значимых видов основных лесных формаций
| Ельники | Пихтарники | Березняки | Осинники | Берёзовые редколесья | |
| Vaccinium myrtillus | 0.421 | 0.423 | 0.383 | 0.482 | 0.449 |
| Gymnocarpium dryopteris | 0.228 | 0.355 | 0.197 | 0.402 | 0.023 |
| Trientalis europaea | 0.140 | 0.194 | 0.154 | 0.174 | 0.158 |
| Linnaea borealis | 0.199 | 0.137 | 0.129 | 0.106 | - |
| Oxalis acetosella | 0.166 | 0.147 | 0.092 | 0.128 | - |
| Equisetum sylvaticum | 0.159 | 0.046 | 0.052 | 0.031 | 0.025 |
| Dryopteris expansa | 0.043 | 0.430 | 0.021 | 0.019 | 0.01 |
| Maianthemum bifolium | 0.057 | 0.191 | 0.066 | 0.141 | - |
| Phegopteris connectilis | 0.002 | 0.108 | 0.001 | - | 0.001 |
| Rubus arcticus | 0.072 | 0.014 | 0.107 | 0.026 | 0.009 |
| Vaccinium vitis-idaea | 0.133 | 0.005 | 0.198 | 0.203 | - |
| Rubus saxatilis | 0.038 | 0.024 | 0.092 | 0.170 | - |
| Melampyrum pratense | 0.026 | 0.024 | 0.042 | 0.104 | 0.009 |
| Avenella flexuosa | 0.058 | 0.106 | 0.114 | 0.007 | 0.368 |
| Solidago virgaurea | 0.028 | 0.038 | 0.054 | 0.064 | 0.245 |
| Bistorta major | 0.001 | 0.035 | - | - | 0.229 |
| Calamagrostis purpurea | 0.076 | 0.159 | 0.057 | 0.057 | 0.198 |
| Veratrum lobelianum | 0.008 | 0.037 | 0.002 | - | 0.134 |
Примечание: Серым цветом выделены показатели КУ видов, значимых во всех формациях, жирным выделены ячейки с показателями КУ видов, играющих значительную роль в отдельных формациях.
В осинниках заметную роль играют виды неморальных и неморально-бореальных фракций. Тогда как в сообществах субформации горных берёзовых редколесий значительная часть списка сосудистых растений представлена видами северных фракций. Состав наиболее ценотически значимых видов горных берёзовых редколесий и криволесий совпадает с аналогичным списком сообществ горных тундр (Дубровский, 2007). Общей чертой списков наиболее значимых с ценотических позиций видов изученных формаций (табл. 3) является преобладание таёжно-лесных видов.
Глава 5. Классификация лесной растительности верхнего и среднего течения р. Илыч
В первом разделе главы даны обзор методов классификации растительности и оценка современного состояния проблемы с акцентом на проблемы лесной типологии и классификации (Алексеев, 1915, 1928; Сукачёв, 1938, 1954, 1972; Воробьёв, 1953, 1967; Погребняк, 1954, 1963; Ниценко, 1956; Колесников, 1961, 1967; Whittaker, 1962; Braun-Blanquet, 1964; Александрова, 1969; Трасс, 1976; Рысин, 1972, 1985, 1995; Миркин, 1989; Блюменталь, 1990; Василевич, 1995, 2007; Миркин, Наумова, 1997 и др.). Показано, что в настоящее время наиболее распространённым методом классификации лесной растительности является эколого-фитоценотический метод, который позволяет учитывать ключевые характеристики лесных сообществ и местообитаний и получать сравнимые результаты (Василевич, 2007). При этом на вооружение в процессе современных лесо-типологических исследований берутся такие практически ценные идеи школы Ж. Браун-Бланке, как детальность выявления видового состава, анализ групп совместно встречающихся видов (Василевич, 1995). Перспективным является также применение ординации описаний с использованием экологических шкал разного уровня (Оценка и сохранение…, 2000; Восточноевропейские…, 2002; Новаковский, 2006).
Во втором разделе дана эколого-фитоценотическая классификация лесной растительности верхнего и среднего течения реки Илыч. В данной схеме учтены результаты исследований, проводившихся на данной территории ранее. В ряде случаев выделены новые ассоциации.
Продромус лесной растительности бассейна р. Илыч
ТИП РАСТИТЕЛЬНОСТИ – ЛЕСНАЯ РАСТИТЕЛЬНОСТЬ
Группа формаций – светлохвойные леса
Формация Sylvatici Pineta
Асс. Pinetum vaginati eriophoroso-sphagnosum*[1]
Асс. Pinetum rostratae caricoso-sphagnosum
Асс. Pinetum globulari caricoso-sphagnosum
Формация Sibirici Lariceta
Субформация Montano-Lariceta
Асс. Montano-Laricetum empetroso-myrtilloso-hylocomiosum*
Субасс. typicum
Субасс. myrtilloso-hylocomiosum
Асс. Montano-Laricetum avenellosum*
Субасс. typicum
Субасс. myrtilloso-hylocomiosum
Группа формаций – темнохвойные леса
Формация Obovati Piceeta
Асс. Piceetum myrtilloso-hylocomiosum
Субасс. typicum
Субасс. avenelloso-myrtilloso-hylocomiosum
Субасс. oxalidoso-myrtilloso-hylocomiosum
Асс. Piceetum fruticuloso-hylocomiosum
Субасс. vaccinioso-hylocomiosum
Субасс. vaccinioso-myrtilloso-hylocomiosum
Субасс. uliginosi vaccinioso-myrtilloso-hylocomiosum
Асс. Piceetum equisetoso-caricoso-myrtilloso-hylocomiosum*
Субасс. typicum
Субасс. globulari caricoso-myrtilloso-hylocomiosum
Асс. Piceetum myrtilloso-gymnocarpioso-hylocomiosum
Субасс. typicum
Субасс. gymnocarpioso-hylocomiosum
Асс. Piceetum saxatili ruboso-hylocomiosum*
Субасс. typicum
Субасс. myrtilloso-saxatili ruboso-hylocomiosum
Асс. Piceetum oxalidoso-hylocomiosum
Вар. Avenella flexuosa
Асс. Piceetum pratensi equisetoso-hylocomiosum*
Вар. Oxalis acetosella
Вар. Gymnocarpium dryopteris
Асс. Piceetum sylvatici equisetoso-hylocomiosum
Асс. Piceetum expansae dryopteridoso-hylocomiosum
Асс. Piceetum myrtilloso-polytrichosum
Асс. Piceetum gymnocarpioso-polytrichosum*
Асс. Piceetum avenelloso-polytrichosum*
Асс. Piceetum aconitosum*
Асс. Piceetum expansae dryopteridosum
Асс. Piceetum sibirici diplaziosum*
Асс. Piceetum purpureae calamagrostidosum
Асс. Piceetum filipendulosum
Вар. Calamagrostis purpurea
Асс. Piceetum oxalidoso-gymnocarpiosum
Субасс. typicum
Субасс. sylvatici equisetosum
Асс. Piceetum sylvatici equisetosum
Вар. Diplazium sibiricum
Вар. Oxalis acetosella
Асс. Piceetum mixto-herbosum
Асс. Piceetum gymnocarpioso-myrtilloso-sphagnosum
Асс. Piceetum globulari caricoso-myrtilloso-sphagnosum
Вар. Equisetum sylvaticum
Вар. Rubus chamaemorus
Асс. Piceetum globulari caricoso-sylvatici equisetoso-sphagnosum
Вар. Rubus chamaemorus
Асс. Piceetum purpureae calamagrostidoso- sylvatici equisetoso-sphagnosum*
Асс. Piceetum pratensi equisetoso-sphagnosum*
Формация – Sibirici Abieteta
Асс. Abietetum myrtilloso-hylocomiosum
Субасс. typicum
Субасс. avenelloso-myrtilloso-hylocomiosum
Асс. Abietetum oxalidoso-hylocomiosum
Асс. Abietetum myrtilloso-gymnocarpioso-hylocomiosum
Субасс. typicum
Субасс. gymnocarpioso-hylocomiosum
Асс. Abietetum saxatili ruboso-hylocomiosum*
Асс. Abietetum gymnocarpiosum*
Асс. Abietetum phegopteridoso-expansae dryopteridosum
Вар. typica
Вар. Dryopteris expansa
Вар. Phegopteris connectilis
Асс. Abietetum sibirici diplaziosum
Асс. Abietetum distentifoli athyriosum*
Асс. Abietetum aconitosum
Асс. Abietetum expansae dryopteridoso-sphagnosum*
Формация – Sibirici Pineta
Асс. Sibirici Pinetum vitis-idaea vaccinioso-hylocomioso-cladinosum
Асс. Sibirici Pinetum myrtilloso-hylocomiosum
Группа формаций – лиственные леса
Формация – Pubescenti Betuleta
Асс. Betuletum vaccinioso-ruboso-hylocomiosum
Субасс. typicum
Субасс. saxatili rubosum
Асс. Betuletum avenelloso-myrtilloso-hylocomiosum
Субасс. typicum
Субасс. myrtillosum
Субасс. avenellosum
Асс. Betuletum myrtilloso-gymnocarpioso-hylocomiosum
Субасс. typicum
Субасс. gymnocarpiosum
Асс. Betuletum myrtilloso-polytrichosum*
Субасс. typicum
Субасс. avenellosum
Асс. Betuletum avenelloso-polytrichosum*
Асс. Betuletum globulari caricoso-polytrichosum*
Асс. Betuletum uliginosi vaccinioso-polytrichosum*
Асс. Betuletum vaccinioso-rubosum*
Асс. Betuletum avenellosum*
Асс. Betuletum saxatili rubosum*
Асс. Betuletum gymnocarpiosum
Асс. Betuletum obtusatae calamagrostidosum
Асс. Betuletum purpureae calamagrostidosum
Асс. Betuletum aconitosum
Асс. Betuletum mixto-herbosum
Асс. Betuletum globulari caricoso-sylvatici equisetoso-sphagnosum
Субасс. globulari caricosum
Субасс. sylvatici equisetosum
Асс. Betuletum purpureae calamagrostidoso-sphagnosum*
Субформация – Montano-Betuleta
Асс. Montano-Betuletum avenelloso-myrtylloso-cetrariosum*
Асс. Montano-Betuletum avenelloso-myrtilloso-hylocomiosum*
Субасс. typicum
Субасс. avenellosum
Асс. Montano-Betuletum myrtilloso-uliginosi vaccinioso-hylocomiosum*
Асс. Montano-Betuletum avenelloso-myrtilloso-polytrichosum*
Субасс. typicum
Субасс. avenellosum
Асс. Montano-Betuletum nardo-polytrichosum*
Асс. Montano-Betuletum avenellosum*
Асс. Montano-Betuletum calamagrostidosum
Асс. Montano-Betuletum albiflori geraniosum*
Асс. Montano--Betuletum mixto-herbosum
Асс. Montano-Betuletum athyriosum
Асс. Montano-Betuletum calamagrostidoso-sphagnosum*
Асс. Montano-Betuletum junco-sphagnosum*
Асс. Montano-Betuletum mixto-herboso-sphagnosum*
Формация – Tremulae Populeta
Асс. Tremulae Populetum myrtilloso-hylocomiosum
Асс. Tremulae Populetum gymnocarpioso-hylocomiosum
Асс. Populetum saxatili ruboso-hylocomiosum*
Асс. Tremulae Populetum vaccinioso-hylocomiosum
Асс. Tremulae Populetum gymnocarpiosum
Асс. Tremulae Populetum saxatili-purpurea ruboso-calamagrostidosum
Асс. Tremulae Populetum mixto herboso-aconitosum
Для каждого синтаксона (от формации до варианта) дана детальная ботаническая и экотопическая характеристика, построены синоптические таблицы. Проведена оценка -разнообразия сосудистых растений, выявлены наиболее значимые с ценотических позиций виды. 35 ассоциаций являются новыми для Печоро-Илычского заповедника. Высокое типологическое разнообразие еловых и берёзовых лесов обусловлено широкой экологической амплитудой видов-эдификаторов. Сообщества обеих формаций отмечены в долинных и водораздельных экотопах, а также в составе растительных комплексов горной ландшафтной зоны. Пихтарники и осинники в силу особенностей экологической валентности видов-эдификаторов формируются в более узком спектре местообитаний, тяготеют к экотопам с несколько более богатыми нормально увлажненными почвами, приуроченным к склонам увалов и речных долин.
Глава 6. Динамика лесной растительности Печоро-Илычского заповедника
В главе рассмотрены процессы восстановления лесной растительности в условиях заповедного режима территории. Первый раздел посвящён характеристике гипотетических рядов динамических смен растительности в пойменных и водораздельных экотопах. В условиях заповедного фонда лесная растительность развивается в режиме спонтанной динамики. Наиболее распространёнными на территории резервата являются старовозрастные еловые леса, которые длительное время не подвергаются сильному влиянию антропогенного фактора (Непомилуева, 1992; Коренные…, 2005). В процессе существования таёжных ненарушенных экосистем имеют место смены растительности, вызванные как изменением экологических условий (формирование лесной растительности на аллювиальных наносах, естественное заболачивание), так и катастрофическими факторами (пожары, ветровалы). Ранее динамическое развитие темнохвойных лесов послепожарного происхождения изучали такие исследователи, как С.В. Алексеев (1948). П.В. Воропанов (1950), Г. Сирен (1955), Валяев (1961), С.А. Дыренков (1984).
Результаты классификации лесов северной части Печоро-Илычского заповедника позволили выделить серии ассоциаций, объединяющие синтаксоны разных формаций, которые различаются только составом видов-эдификаторов. Данный подход используется нами вслед за С.Я. Соколовым (1962), Д.Н. Сабуровым (1972), В.И. Василевичем (2000) и С.В. Дёгтевой (2002). Коренные темнохвойные сообщества сменяются во времени лесами сложенными Betula pubescens и Populus tremula. При этом коренное сообщество либо восстанавливается, либо трансформируется в другой фитоценоз (Рис. 2). установлено, что разнообразие производных сообществ выше, чем коренных. Так, на месте фитоценозов асс. Piceetum/Abietetum myrtilloso-hylocomiosum после катастрофического воздействия в пределах исследованной территории формируются производные мелколиственные леса, принадлежащие пяти синтаксонам (рис. 2) зеленомошной и долгомошной групп типов (Betuletum avenelloso-myrtilloso-hylocomiosum, Populetum myrtilloso-hylocomiosum, Betuletum mixto-herbosum, B. avenelloso-polytrichosum, B. myrtilloso-polytrichosum). Данные сообщества сменяются фитоценозами, принадлежащими ассоциациям Piceetum/Abietetum myrtilloso-hylocomiosum, Piceetum avenelloso-polytrichosum, P. myrtilloso-polytrichosum.
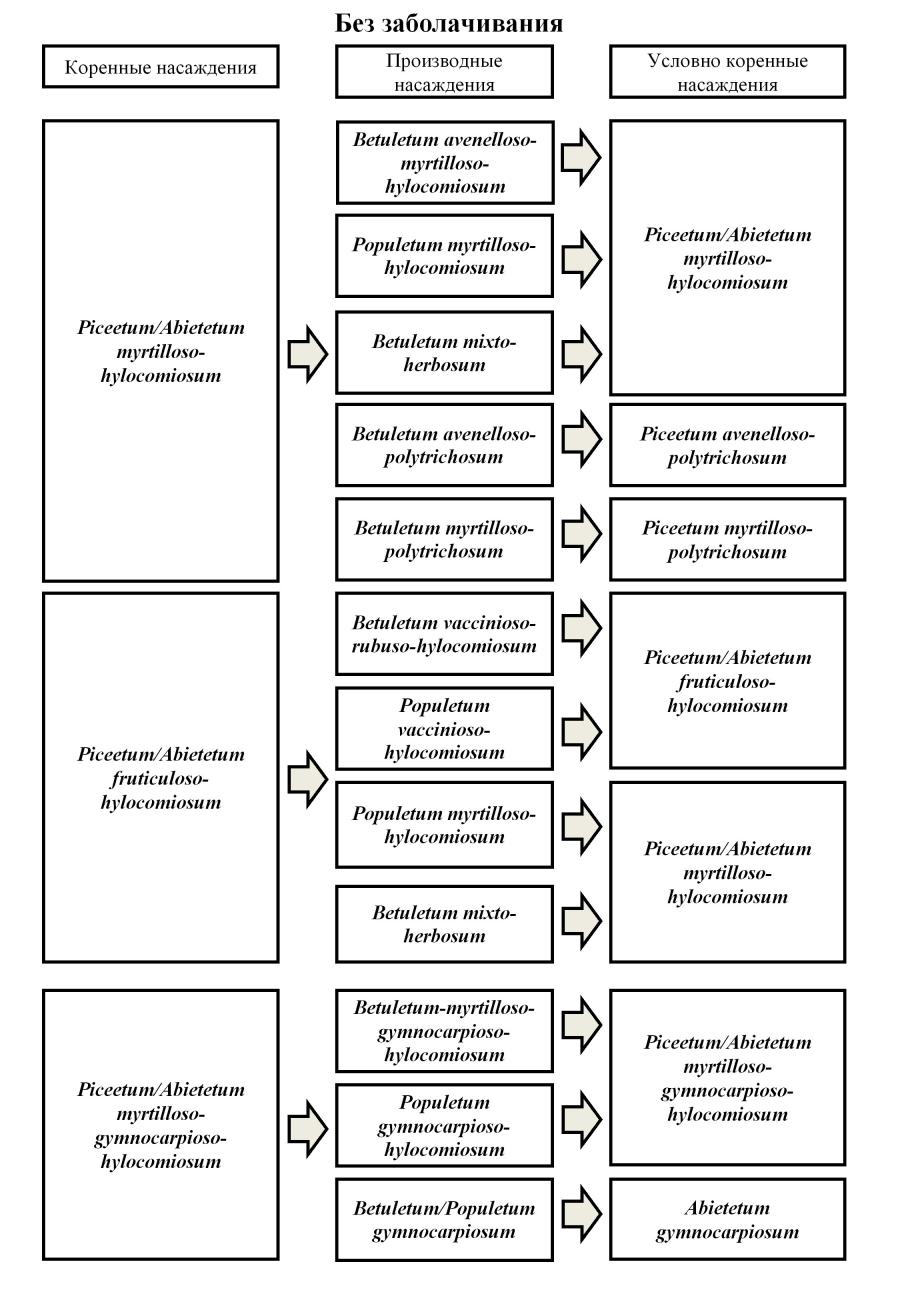
Рис. 2 Некоторые схемы вторичных (демутационных) сукцессий в северной части Печоро-Илычского заповедника (без заболачивания)
При уничтожении темнохвойных сообществ асс. Piceetum/Abietetum fruticuloso-hylocomiosum (рис. 2) образуются производные лиственные насаждения четырёх ассоциаций: Betuletum vaccinioso-ruboso-hylocomiosum, Populetum vaccinioso-hylocomiosum, P. myrtilloso-hylocomiosum, Betuletum mixto-herbosum. Такие сообщества в ходе смены эдификаторов постепенно трансформируются в условно-коренные фитоценозы асс. Piceetum/Abietetum fruticuloso-hylocomiosum (происходит восстановление сообществ исходного синтаксона) или асс. Piceetum/Abietetum myrtilloso-hylocomiosum (происходит смена синтаксона). Сообщества асс. Piceetum/Abietetum myrtilloso-gymnocarpioso-hylocomiosum сменяются березняками и осинниками ассоциаций Betuletum-myrtilloso-gymnocarpioso-hylocomiosum и Populetum gymnocarpioso-hylocomiosum, на месте которых в процессе демутации восстанавливаются исходные фитоценозы асс. Piceetum/Abietetum myrtilloso-gymnocarpioso-hylocomiosum. В результате необратимого уничтожения мохового покрова при пожарах происходит формирование лиственных сообществ асс. Betuletum/Populetum gymnocarpiosum, которые замещаются пихтарниками папоротничковыми (Abietetum gymnocarpiosum), или фитоценозами зеленомошной группы типов (Piceetum/Abietetum myrtilloso-gymnocarpioso-hylocomiosum).
Таблица 4
Таксационные параметры берёзовых насаждений зеленомошной группы типов
| Древостои | Класс возраста насаждения | Амплитуда высоты стволов, м | Амплитуда диаметра стволов,см | Общая сомкнутость крон |
| Молодые | I | 0.5–3 | 2–4 | 0.7–0.9 |
| II–III | 6–12 | 6–15 | 0.4–0.8 | |
| средневозрастные | IV | 8–12 | 8–24 | 0.4–0.7 |
| V | 12–16 | 10–24 | 0.4–0.8 | |
| Приспевающие | VI | 14–18 | 16–26 | 0.4–0.8 |
| Спелые | VII–VIII | 18–22 | 24–44 | 0.4–0.8 |
| Перестойные | XIII–XIV | 22–24 | 24–70 | 0.5–0.7 |
Примечание: римскими цифрами указаны классы возраста древостоев
Во втором разделе на примере сообществ березняков и ельников зеленомошной группы типов анализируется динамика растительного покрова при восстановительных сукцессиях на гарях. Показано, что в бассейне р. Илыч на гарях в течение первых десяти лет после пожаров формируются сомкнутые молодняки с доминированием Betula pubescens. В производных насаждениях различного возраста таксационные характеристики древостоев претерпевают закономерные изменения (табл. 4). При увеличении возраста насаждений происходит усложнение структуры древесного яруса, береза и осина в результате изменения светового режима под пологом леса замедляют темпы возобновления уже в сообществах III класса возраста. В насаждениях VI–VII классов возраста идёт активное формирование нижнего яруса из хвойных пород (в березняках VIII класса возраста ели и пихта явно доминируют в составе нижних пологов древостоя), которые в спелых насаждениях начинают выходить в I полог древостоя. Начиная с X–XI классов возраста происходит постепенный распад исходного лиственного насаждения и его смена сначала лиственно-хвойным (XII–XIV классы возраста), а затем хвойным древостоем. Число стволов берёзы в процессе демутационной сукцессии снижается с 36–76 тыс. до 64–232 шт./ га. Наиболее активная дифференциация насаждений березы (период наибольшего сокращения числа стволов) происходит в молодых древостоях. В результате описанных процессов формируются еловые древостои с типичной для темнохвойной тайги разновозрастной структурой. Активное возобновление Picea obovata уже в березняках I класса возраста (12–31 тыс. шт. подроста ели на 1 га) свидетельствует об ускоренном процессе восстановления коренного сообщества. Этот факт может быть связан с регулярным заносом семян ели из соседних ненарушенных экосистем.
Смена эдификаторов лесных сообществ приводит к трансформации нижних ярусов фитоценозов, смене доминантов травяно-кустарничкового яруса (Мелехов, 1954, 1968; Некоторые вопросы типологии…, 1972; Ильчуков, 1997; Ценотическая и флористическая…, 2001; Дегтева, 2002; Крышень, 2006; Уланова, 2007). На территории Печоро-Илычского заповедника такие исследования проводились впервые и позволили выявить виды, получающие преимущества в условиях улучшения светового режима на гарях и под пологом производных лиственных лесов (Rubus arcticus, Chamaenerion angustifolium, Avenella flexuosa, Luzula pilosa, Solidago virgaurea, Deschampsia cespitosa и др.), растения, восстанавливающие своё значение в ходе демутационной сукцессии (Maianthemum bifolium, Oxalis acetosella, Equisetum sylvaticum, Gymnocarpium dryopteris и др.), а также виды, относительно толерантные к пирогенным воздействиям (Vaccinium myrtillus, Linnaea borealis, Trientalis europaea, Carex globularis, Calamagrostis purpurea и др.). Напочвенный покров восстанавливается достаточно быстро (его общее проективное покрытие уже к 9-летнему возрасту фитоценоза достигает 80 %). На начальных стадиях доминируют два вида мхов (Pleurozium schreberi, Polytrichum commune). В процессе дальнейшего развития роль кукушкина льна снижается, число видов, способных достигать удельного обилия 4 балла увеличивается. В процессе дифференциации насаждений появляется мертвая древесина, на которой активно развиваются представители рода Dicranum. Микропонижения занимают сфагновые мхи, прежде всего Sphagnum girgensohnii.
Выводы
1. В лесных сообществах верхнего и среднего течения реки Илыч (в границах Печоро-Илычского заповедника) выявлено 292 вида сосудистых растений, принадлежащих к 163 родам и 60 семействам. С фитогеографической точки зрения флору изученных лесных сообществ можно охарактеризовать как бореальную (204 вида или 69.9 %) с преобладанием видов сосудистых растений с голарктическими и евразиатскими ареалами (36.6 % и 39 % соответственно). По жизненной форме абсолютное большинство растений, характерных для лесов (236 видов или 80.8 %), относится к травам. Анализ отношения видов, произрастающих в лесах, к фактору общего богатства почв показывает, что наибольшее их число (130 или 44.8 %) является мезотрофами. По отношению к фактору увлажнения наибольшую долю (148 видов или 50.7 %) в списке составляют мезофиты. Среди выделенных эколого-ценотических групп видов наибольшим разнообразием отличаются долинные, растения которых предпочитают наиболее благоприятные с точки зрения факторов увлажнения и богатства почвы экотопы долин крупных и малых водотоков (39.7 % – 110 видов).
2. В результате формационного анализа лесных ценофлор установлено, что наиболее богатой во флористическом отношении является формация ельников (223 вида), играющая определяющую роль в сложении растительного покрова резервата и отличающаяся широкой экологической амплитудой. По сравнению с объединенной ценофлорой сосудистых растений исследованной территории для ценофлор темнохвойных лесов характерно увеличение доли бореальных видов (до 73.4 %), снижение значения представителей северных фракций (до 17.4 %), сохранение соотношения долготных элементов и явное доминирование таёжно-лесных видов (21.4 %). Лиственные леса (березняки и осинники) характеризуются большим видовым разнообразием сосудистых растений, чем хвойные, что связано с увеличением богатства почв под пологом таких сообществ. Для сообществ березняков и осинников зарегистрировано увеличение доли неморальных и неморально-бореальных видов, а также растений из долинных эколого-ценотических групп. Берёзовые редколесья и криволесья характеризуется специфическими особенностями флористического состава, сближающими их с сообществами горных тундр.
3. В результате классификации лесной растительности северной части Печоро-Илычского заповедника составлен продромус лесной растительности. Выделено 80 ассоциаций, которые распределяются по формациям следующим образом: сосняки – 3, лиственничники – 2, ельники – 27, пихтарники – 10, кедровники – 2, березняки – 17, осинники – 7, берёзовые криволесья и редколесья – 12. Впервые в границах резервата выделены 35 ассоциаций.
4. При изучении трансформации структуры древостоев в ходе восстановительной сукцессии на примере сообществ березняков разного возраста, березово-еловых и еловых лесов зеленомошной группы типов показано, что на ранних стадиях пирогенной демутации формируется сомкнутый древостой из Betula pubescens. При увеличении возраста насаждений число стволов берёзы снижается с 36–76 тыс. шт./ га в древостоях I класса возраста до 64–232 шт./га в насаждениях XIV класса возраста, происходит усложнение структуры древесного яруса. Активное возобновление Picea obovata уже на ранних стадиях формирования древостоев (12–31 тыс. шт. подроста ели на 1 га в молодняках березы I класса возраста) свидетельствует об ускоренном процессе восстановления коренного сообщества.
5. Установлено, что на гарях и под пологом лиственных лесов в условиях улучшения светового режима получают преимущества травянистые растения: Rubus arcticus, Chamaenerion angustifolium, Avenella flexuosa, Luzula pilosa, Solidago virgaurea, Deschampsia cespitosa и др.). В процессе смены видов-эдификаторов постепенно восстанавливают своё ценотическое значение Maianthemum bifolium, Oxalis acetosella, Equisetum sylvaticum, Gymnocarpium dryopteris и др. Относительную толерантность к пирогенным воздействиям демонстрируют Vaccinium myrtillus, Linnaea borealis, Melampyrum pratense, Trientalis europaea, Carex globularis, Calamagrostis purpurea и др. Состав и структура нижних ярусов лесных сообществ в ходе демутационных сукцессий на гарях восстанавливаются быстрее, чем на вырубках.
Список основных работ, опубликованных по теме диссертации
– в изданиях, рекомендованных ВАК:
Дёгтева С.В. Ценотическое и флористическое разнообразие берёзовых криволесий и редколесий северной части Печоро-Илычского заповедника / С.В. Дёгтева, Ю.А. Дубровский, Т.П. Шубина // Ботан. журн. – 2009. - Т. 94. № 7. – С. 1037-1056.
– в прочих изданиях:
Дубровский Ю.А. Горные леса и редколесья северного Урала (Печоро-Илычский государственный биосферный заповедник) / Ю.А. Дубровский // Актуальные проблемы геоботаники. III Всероссийская школа-конференция. I часть. – Петрозаводск: Карельский научный центр РАН, 2007. – С. 172–176.
Дубровский Ю.А. Видовое и ценотическое разнообразие растительного покрова в высотном градиенте на западном макросклоне Северного Урала / Ю.А. Дубровский // Материалы I(III) Всерос. молодёж. научно-практической конф. ботаников в Новосибирске (Новосибирск, 17–21 октября 2007 г.). – Новосибирск: изд-во СО РАН, 2007. – С. 86–90.
Дубровский Ю.А. Видовое разнообразие и структура растительного покрова в высотном градиенте в пределах западного макросклона Северного Урала / Ю.А. Дубровский // Сибирский ботанический вестник: электронный журнал. – 2007. - Т. II. Вып. 2. – С. 3–8.
Дёгтева С.В. Ценотическое и флористическое разнообразие берёзовых криволесий и редколесий северной части Печоро-Илычского заповедника / С.В. Дёгтева, Ю.А. Дубровский // Принципы и способы сохранения биоразнообразия: матер. III всерос. науч. конф. – Пущино, 2008. – С. 134–136.
Дубровский Ю.А. Лесная растительность северной части Печоро-Илычского биосферного заповедника / Ю.А. Дубровский, С.В. Дёгтева // Материалы Междунар. науч. конф., посв. 135-летию со дня рожд. И.И. Спрыгина. – Пенза, 2008. – С. 193–194.
Дубровский Ю.А. Разнообразие лесной растительности северной части Печоро-Илычского биосферного заповедника / Ю.А. Дубровский // Мониторинг и оценка состояния растительного мира. Материалы Междунар. науч конф. – Минск: Право и экономика, 2008. – С. 150–152.
Дубровский Ю.А. Видовое разнообразие сосудистых растений в лесных экосистемах северной части Печоро-Илычского заповедника / Ю.А Дубровский // Актуальные проблемы биологии и экологии. Материалы докл. XVI Всеросс. молодёж. науч. конф. – Сыктывкар, 2009. – С. 53–55.
[1] * – ассоциации, выделенные для территории Печоро-Илычского заповедника впервые