

Генетический полиморфизм популяций юга центральной россии (по данным об иммуно-биохимических генных маркерах)
| РОССИЙСКАЯ АКАДЕМИЯ МЕДИЦИНСКИХ НАУК ГУ МЕДИКО-ГЕНЕТИЧЕСКИЙ НАУЧНЫЙ ЦЕНТР |
На правах рукописи
УДК 575.17
Жерлицына Мария Сергеевна
ГЕНЕТИЧЕСКИЙ ПОЛИМОРФИЗМ ПОПУЛЯЦИЙ ЮГА ЦЕНТРАЛЬНОЙ РОССИИ
(ПО ДАННЫМ ОБ ИММУНО-БИОХИМИЧЕСКИХ
ГЕННЫХ МАРКЕРАХ)
03.00.15 генетика
03.00.14 антропология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени кандидата
биологических наук
Москва – 2006
Работа выполнена на кафедре медико-биологических дисциплин ГОУ ВПО Белгородского государственного университета
Научные руководители:
доктор медицинских наук, профессор М.И. Чурносов
доктор биологических наук Е.В. Балановская
Официальные оппоненты:
доктор биологических наук, профессор В.А. Спицын
кандидат биологических наук О.Л. Курбатова
Ведущее учреждение:
Московский государственный медико-стоматологический университет
Защита состоится «16» октября 2006 года в 14 часов на заседании Диссертационного совета Д 001.016.01, при ГУ МГНЦ РАМН по адресу: 115478, г. Москва, ул. Москворечье, д.1.
С диссертацией можно ознакомиться в библиотеке ГУ Медико-Генетического Научного Центра РАМН
Автореферат разослан «13» сентября 2006 года
Ученый секретарь Диссертационного совета
доктор биологических наук, профессор Л.Ф. Курило
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы. Изучение структуры генофонда различных групп народонаселения и решение вопросов микроэволюции популяций человека, их происхождения, родства, взаимодействия со средой является одной из важнейших задач современной антропогенетики (Рычков, 2000; Алтухов, 2003; Гинтер, 2003).
К настоящему времени проведены масштабные популяционно-генетические исследования различных этнотерриториальных групп Северной Евразии: народов Поволжья и Урала, Кавказа, Средней Азии, Сибири и Дальнего Востока (Рычков и др., 1983, 1984; Спицын и др., 1991, 1995, 1999; Кравчук и др. 1996, 1998; Шнейдер и др., 1991, 1995; Инасаридзе и др., 1990; Насидзе и др., 1990; Нерсисян и др., 1994, 1996; Шенгелия и др., 1991; Кучер и др., 1993, 2000; Сукерник и др., 1977, 1986; Посух и др., 1990; Соловенчук и др., 1982, 1985; и др.). Однако для описания генетической структуры популяций каждая группа исследователей использовала свой, зачастую ограниченный, набор иммуно-биохимических маркеров. Из-за этого проведение корректного сравнительного анализа генофондов изученных популяций весьма затруднено и возможно лишь по 5-6 биохимическим маркерам (HP, GC, TF, GLO1, PGM1, ACP1) и некоторым иммунологическим (АВО, RH, MNSs, LEW, KEL). Но при этом классические маркеры иммуно-биохимического полиморфизма изучены лучше, чем ДНК маркеры.
Сейчас все большее число генетических лабораторий переходит к изучению только ДНК полиморфизма, сворачивая исследование классических маркеров. Однако потенциальный набор ДНК маркеров столь велик, что сложилась ситуация, когда провести сравнительный анализ разных генофондов по аутосомным ДНК маркерам почти невозможно – каждый изучен по собственному набору маркеров. Классические же маркеры охватывают много более широкий круг популяций, чем ДНК маркеры. Например, дать характеристику генофонда народов Восточной Европы можно лишь по 6 аутосомным ДНК маркерам, охватив в среднем 28 популяций (от 11 до 47 по разным маркерам). В то же время по 33 классическим маркерам мы охватываем в среднем 103 популяции (от 12 до 881 популяции по разным маркерам) (Балановская и др., 2006). Показано, что, изучая классические маркеры, можно составить надежный прогноз изменчивости генофонда по ДНК маркерам: межпопуляционная изменчивость по ДНК маркерам и по классическим маркерам одинакова (Bowcock et all., 1987; Cavalli-Sforza et all., 1994; Лимборская и др., 2002; Балановская и др., 2006). Анализ классических маркеров в настоящее время позволяет дать не только более объективную характеристику генофонда, но и прогноз для изучения ДНК полиморфизма. Поэтому в «ДНК эру» изучение классических маркеров приобретает новое и актуальное значение.
Несмотря на обширный фактический материал по иммуно-биохимическому полиморфизму народонаселения России, популяционно-генетические сведения о русском народе – самом многочисленном в нашей стране – остаются явно недостаточными. Большинство русских популяций, проживающих в пределах исторического ареала, исследовано по ограниченному набору маркеров. Изучение генетической структуры различных групп русского населения по широкому спектру иммуно-биохимических маркеров проводилось лишь в нескольких исследованиях (Спицын и др., 2001; Шнейдер и др., 1994, 2002; Балановская и др., 2001а,б). Информация о генетическом полиморфизме с привлечением большого единого набора иммуно-биохимических маркеров (8-10 маркеров) имеется лишь для небольшого числа (около 15) русских популяций. Как отмечает коллектив авторов «Генофонд и геногеография народонаселения. Т.1. Генофонд населения России и сопредельных стран», в силу большого объема этого генофонда, обширности ареала, сложности этнической истории, интенсивных изменений в структуре населения, миграции сельского населения в города, и ряда других причин генофонд русского населения остается наименее исследованным и наиболее трудным для изучения.
Недостаточная изученность русского генофонда по единому большому спектру иммуно-биохимических маркеров диктует необходимость проведения дальнейших исследований.
Есть и еще один важный аспект данной работы – она впервые проведена для небольшого подразделения русского народа. Обычно же оценки генетической изменчивости даются на межэтническом уровне или основаны на масштабных внутриэтнических выборках, представляющих географически и исторически контрастные группы популяций. Предприняв изучение генетической структуры узкого круга популяций юга Центральной России, нам предстояло впервые выяснить, имеется ли сколько-нибудь значимое генетическое разнообразие локальной выборки русских, представляющих лишь небольшую часть русского народа?
Цели и задачи исследования. Изучить генофонд коренного русского населения юга Центральной России и его место среди популяций русского народа и других восточных славян.
В соответствии с поставленной целью были сформулированы следующие задачи:
1. Дать характеристику генофонда коренного русского населения юга Центральной России по данным о распределении частот 29 аллелей 11 локусов иммуно-биохимических маркеров.
2. Выявить степень генетической дифференциации русских популяций юга Центральной России.
3. Изучить генетические соотношения между разными популяциями юга Центральной России.
4. Оценить место генофонда населения юга Центральной России в системе генофондов русского народа и восточных славян.
Научная новизна. Впервые изучены особенности структуры генофонда популяций юга Центральной России. Получены данные о распределении 29 аллелей 11 иммуно-биохимических локусов. Дана оценка уровня генетической дифференциации коренного русского населения четырех областей юга Центральной России.
Выявлены особенности генетической подразделенности популяций юга Центральной России и их соответствие географическим расстояниям.
Установлена дифференциация населения юга Центральной России на две группы, одна из которых генетически сходна с населением Московской области, другая – с жителями Рязанской области, что соответствует истории формирования изученных популяций.
Определено положение генофонда популяций юга Центральной России в структуре украинского и белорусского генофонда.
Научно-практическая значимость работы. Изучена генетическая структура коренного русского населения юга Центральной России по единому большому спектру иммуно-биохимических маркеров. Выявлены особенности иммуно-биохимического полиморфизма среди различных групп русского населения.
Проведена оценка генетической дифференциации коренного русского населения юга Центральной России. Установлено наличие подразделенности между группами русского населения юга Центральной России. На основе полученных данных определено положение генофонда популяций юга Центральной России в системе восточнославянского генофонда.
Полученные данные послужат основой для генетического и экологического мониторинга населения юга Центральной России. Результаты исследования используются в учебном процессе в ГОУ ВПО Белгородском государственном университете, в работе ГУ Медико-генетического научного центра РАМН, ГОУ ВПО Кубанской государственной медицинской академии, Харьковского государственного университета им. В.Н. Каразина.
Положения, выносимые на защиту.
1. Генофонд коренного русского населения юга Центральной России имеет четко выраженные западно-евразийские характеристики.
2. Генетические расстояния между популяциями юга Центральной России соответствуют географическим расстояниям между ними.
3. Популяции юга Центральной России дифференцируются на две группы, генетически близкие к тем русским популяциям, которые служили источниками миграций в X - XVIII веках.
4. В популяционной системе восточных славян население юга Центральной России генетически близко к «среднерусской» популяции и украинцам, и значительно удалено от белорусов.
Апробация работы. Основные результаты диссертации доложены и обсуждены на: V Бунаковских чтениях «Теория антропологии и ее методы. Итоги и развитие» (Москва, 2001), Итоговой научной сессии молодых ученых Курского госмедуниверситета (2002, 2003, 2004), Годичной научной конференции сотрудников Белгородского госуниверситета (Белгород, 2002, 2003, 2004, 2005), Международной конференции «Антропология на пороге III тысячелетия (Москва, 2004), Третьем съезде ВОГиС «Генетика в XXI веке: современное состояние и перспективы развития» (Москва, 2004), Втором Международном экономическом форуме Собора славянских народов (Белгород, 2004), Третьих Антропологических чтениях памяти акад. В. П. Алексеева «Экология и демография человека в прошлом и настоящем» (Москва, 2004), Юбилейной научной конференции КГМУ и сессии Центрально-Черноземного научного центра РАМН, посвященной 70-летию КГМУ (Курск, 2005), Пятом съезде Российского общества медицинских генетиков (Уфа, 2005), Российской научной конференции с международным участием «Медико–биологические аспекты мультифакториальной патологии» (Курск, 2006), 71-й научной конференции КГМУ и сессии Центрально-Черноземного центра РАМН (Курск, 2006).
Публикации. По теме диссертации опубликовано 15 работ.
Структура и объем диссертации.
Диссертация изложена на 226 страницах и состоит из введения, обзора литературы, описания материалов и методов исследования, собственных результатов, заключения, выводов, списка литературы. Работа иллюстрирована 19 таблицами и 32 рисунками. Дано приложение на 17 страницах. Библиографический указатель содержит 215 наименований, из которых 63 иностранных.
МАТЕРИАЛЫ И МЕТОДЫ
Проведено изучение генетической структуры популяций юга Центральной России по 11 локусам иммуно-биохимических маркеров (АВО, RH, HP, GC, TF, C’3, ACP1, GLO1, PGM1, ESD, 6-PGD). Объектами исследования послужили пять районных популяций четырех областей юга Центральной России: Белгородской (Яковлевский район), Орловской (Ливенский район), Воронежской (Репьевский район), Курской (Пристенский и Черемисиновский районы) (рис.1). Выбор данных популяций юга Центральной России в качестве объектов исследования обусловлен особенностями их формирования в 10-18 в.в., их различным славянским субстратом, разнонаправленностью миграционных потоков в эти популяции из разных территориальных групп русских, различным вкладом генофонда восточнославянских народов (украинцев, белорусов) в генофонд населения этих районов. В настоящее время данные области входят в состав Центрального Федерального Округа России.
Население рассматриваемых областей практически однонационально: русские составляют 93-96%, на долю украинцев приходится 3-5%, представители других национальностей (армяне, узбеки, татары и др.) вместе составляют 1.6-2.6%.
Общий объем выборки составил 493 человека, представляющих коренное русское население четырех областей: 141 человек - из Белгородской области (Яковлевский район), 127 – из Орловской области (Ливенский район), 117 – из Воронежской области (Репьевский район), и 108 – из Курской области (46 человек из Пристенского района, 62 – из Черемисиновского района). Средний возраст изученного населения составил 42,2 года.
Важнейшая задача работы – выявить, существуют ли межпопуляционные различия между территориально близкими популяциями русского народа. Поэтому для исследования были выбраны территориально близкие популяции в пределах четырех областей юга Центральной России: среднее расстояние между изучаемыми популяциями составляет 125 км. Для выявления достоверной картины мы при планировании исследования формировали более многочисленные (100 - 150 индивидуумов) выборки по каждой из исследуемых областей, чем это необходимо для выявления
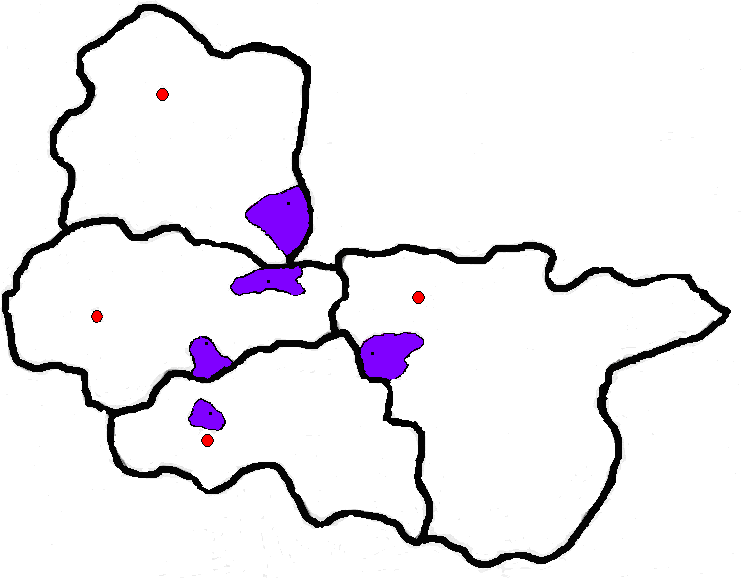
Рис.1. Территориальное расположение ряда областей юга Центральной России (отмечены районы, в которых производился сбор материалов для исследования).
различий между этносами, где достаточной считается выборка из 50 – 70 человек.
Полевой сбор образцов проведен М.С. Жерлицыной, И.Н. Лепендиной, В.Ю. Песик, Н.А. Рудых, И.Н. Сорокиной под рук. д.б.н. Е.В. Балановской и проф. М.И. Чурносова.
Особое внимание было уделено корректному формированию репрезентативных выборок: в них включались только неродственные (до третьей степени родства) индивиды, все предки которых на глубину трех поколений происходят из данной популяции. Взятие образцов крови осуществлялось согласно международным стандартам с письменного согласия обследуемых и под контролем Этической комиссии ГУ МГНЦ РАМН.
Материалом для лабораторного исследования послужила венозная кровь, взятая из локтевой вены у пробанда. Общий объем полученного образца составил 5-6 мл. Разделение на сывороточные и эритроцитарные фракции производилось путем центрифугирования (3 тыс. об/мин.) в вакуумных пробирках с гелем, который разделял фракции крови. Дальнейшее хранение материала осуществлялось при -20 С°.
Группы крови АВО и RH выявляли в полевых условиях по результатам опроса обследуемых лиц. Идентификацию биохимических локусов HP, С'3, 6-PGD осуществляли стандартным методом вертикального электрофореза в 7,5% полиакриламиде (ПААГ), GLO1 - в 5% ПААГ (Спицын, 1985; Westermeier, 2001). Локусы GC, TF, PGM1, ESD типировались методом изоэлектрофокусирования (ИЭФ) в ПААГ (Altland еt all. 1980; Спицын, 1990; Gradl еt all., 1985; Rigetti, 1983), а локус АСР - методом ИЭФ в агарозе (Frants еt all., 1978, Спицын, 1985).
Статистический анализ. Определение частот аллелей, генотипов, оценку соответствия характера распределения частот генотипов равновесию Харди-Вайнберга (2), расчет наблюдаемой (HО) и ожидаемой гетерозиготности (HЕ), индекса фиксации Райта (D) производили по стандартным формулам (Животовский, 1991; Спицын и др., 1996; Ли, 1978). Показатели генной идентичности (IT), общего (HT), внутрипопуляционного (HS) и межпопуляционного (GST, DST) генетического разнообразия рассчитывали согласно Nei (1973, 1977).
Генетические расстояния между популяциями были рассчитаны с помощью программы DJ genetic (версия 0,03 beta), разработанной Ю.А. Серегиным и Е.В. Балановской в ГУ МГНЦ РАМН.
На основе полученных матриц генетических расстояний были построены дендрограммы, проведено многомерное шкалирование; по корреляционным матрицам - факторный анализ (Дерябин, 2001) с использованием пакета статистических программ Статистика (версия 5).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
- Генетический полиморфизм населения юга Центральной России
Среди популяций юга Центральной России частоты генов изученных 11 иммуно-биохимических систем составили: АВО*А=0.25, АВО*В=0.16, АВО*О=0.59, RH*D=0.62, HP*1=0.34, С’3*S=0.88, GC*2=0.27, GC*1S=0.64, GC*1F=0.09, TF*C1=0.77, TF*C2=0.15, TF*C3=0.07, TF*В=0.01, GLO1*1=0.30, ESD*1=0.91, ESD*2=0.08, ESD*5=0.01, 6-PGD*A=0.97, ACP1*A=0.31, ACP1*В=0.60, ACP1*С=0.09, PGM1*1S=0.65, PGM1*1F=0.07, PGM1*2S=0.24, PGM1*2F=0.04. Средние частоты генов RH*D и GLO1*1 у населения юга Центральной России достоверно ниже (р<0.01-0.001), чем среднерусские показатели по этим аллелям. Частоты генов GC*1S, PGM1*1S, ACP1*С лежат на верхней границе изменчивости русского генофонда и достоверно (р<0.01) выше среднерусских частот соответствующих аллелей (рис.2). Выявлено, что генетическая структура популяций юга Центральной России характеризуется ярко выраженным западно-евразийским распределением частот аллелей TF*C2, GLO1*1, ESD*2 и ACP1*С.
Сравнивая вариабельность частот аллелей в популяциях юга Центральной России с данными по русскому генофонду, можно выделить несколько групп аллелей по наличию в их изменчивости географических закономерностей.
Во-первых, это аллели, которые имеют четкий географический градиент в изменчивости среди населения юга Центральной России - АВО*О и АВО*В. Так, для аллеля АВО*О, как для русского генофонда в целом (Балановская и др., 2006), характерна тенденция снижения частоты с севера, северо-востока на юг, юго-запад (с 0.62 до 0.52). Аллель АВО*В, в отличии от паттерна «лоскутного одеяла», выявленного для русского населения (Балановская и др., 2006), в популяциях юга Центральной России закономерно снижается с юга (0.22) на север (0.11).
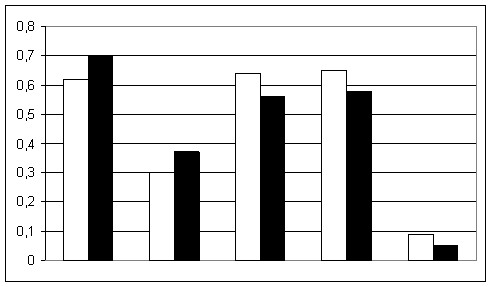
Рис.2. Cредние частоты ряда иммуно-биохимических маркеров среди русского населения и населения юга Центральной России
Во-вторых, ряд аллелей на территории юга Центральной России имеют изменчивость, связанную с пиком возрастания (АВО*А, PGM1*1S, ACP1*С) или падения (PGM1*1F) частоты по линии «северо-запад - юго-восток», проходящей по территории Курской области.
В-третьих, в популяциях юга Центральной России максимальная вариабельность частот, достигающая размаха изменчивости всех русских популяций, выявлена для аллелей локуса GC (GC*2 – 0.19< р <0.37; GC*1S – 0.56< р <0.71). Однако при этом географическая изменчивость частот этих аллелей полностью согласуется с общерусским генетическим рельефом аллеля GC*2 (Балановская и др., 2006): по изученным популяциям юга Центральной России проходит как область минимальных (по 40° долготы), так и область субмаксимальных частот этого аллеля.
В-четвертых, самую многочисленную группу составили аллели, вариабельность которых в популяциях юга Центральной России незначительна (аллели локусов HP, С’3, TF, GLO, ESD, 6-PGD) или их изменчивость не имеет четких географических закономерностей (RH).
- Генетические различия между русскими популяциями юга Центральной России
По данным о частотах 29 аллелей 11 локусов иммуно-биохимических систем крови дана оценка генетической дифференциации населения юга Центральной России. Исследование проведено на модели семи районных популяций: пять из них изучены автором, две популяции привлечены для сравнения, поскольку изучены нашим коллективом (Лепендина, 2005) по идентичному набору маркеров. Установлено, что уровень генной дифференциации коренного русского населения юга Центральной России составляет ![]() =0.0052. Наибольший вклад в формирование генетической изменчивости популяций юга Центральной России, как и ожидалось в соответствии с наибольшим вариационным размахом частот аллелей, вносит система GС (GST=0.0258), что может указывать на действие дифференцирующего отбора в отношении аллелей данного локуса. Действие стабилизирующего отбора в популяциях юга Центральной России можно предположить в отношении аллелей локусов HP, С’3, GLO1, 6-PGD, ACP1, которые характеризуются наименьшим уровнем генной дифференциации (GST<0.0031).
=0.0052. Наибольший вклад в формирование генетической изменчивости популяций юга Центральной России, как и ожидалось в соответствии с наибольшим вариационным размахом частот аллелей, вносит система GС (GST=0.0258), что может указывать на действие дифференцирующего отбора в отношении аллелей данного локуса. Действие стабилизирующего отбора в популяциях юга Центральной России можно предположить в отношении аллелей локусов HP, С’3, GLO1, 6-PGD, ACP1, которые характеризуются наименьшим уровнем генной дифференциации (GST<0.0031).
Важно подчеркнуть, что для анализируемых нами популяций юга Центральной России получены три оценки по трем разным типам генетических маркеров: 1) классические маркеры (иммуно-биохимические) - ![]() =0.0052 (наши данные); 2) аутосомные ДНК маркеры -
=0.0052 (наши данные); 2) аутосомные ДНК маркеры - ![]() =0.0059 (Песик, 2006); 3) квазигенетические маркеры (фамилии) - fr* =0.0062 (Сорокина, 2006). Выявлено полное совпадение всех оценок генетической дифференциации -
=0.0059 (Песик, 2006); 3) квазигенетические маркеры (фамилии) - fr* =0.0062 (Сорокина, 2006). Выявлено полное совпадение всех оценок генетической дифференциации - ![]() =0.00520.00590.0062. Сходство всех трех оценок показывает высокую надежность полученных нами оценок дифференциации популяций юга Центральной России - небольшого регионального подразделения русского генофонда. Это соответствует выводам исследования Ю.Г. Рычкова и Е.В. Балановской, которые еще в 1980-х гг. с помощью теоретического популяционно-генетического анализа показали, а затем подтвердили фактическим материалом закономерность: данные о внутрипопуляционной изменчивости (гетерозиготности НS) для классических и ДНК маркеров могут чрезвычайно сильно различаться; однако при этом оценки межпопуляционной изменчивости (дифференциации популяций FST и GST) для классических и ДНК маркеров должны быть одинаковы, если выборки репрезентативны.
=0.00520.00590.0062. Сходство всех трех оценок показывает высокую надежность полученных нами оценок дифференциации популяций юга Центральной России - небольшого регионального подразделения русского генофонда. Это соответствует выводам исследования Ю.Г. Рычкова и Е.В. Балановской, которые еще в 1980-х гг. с помощью теоретического популяционно-генетического анализа показали, а затем подтвердили фактическим материалом закономерность: данные о внутрипопуляционной изменчивости (гетерозиготности НS) для классических и ДНК маркеров могут чрезвычайно сильно различаться; однако при этом оценки межпопуляционной изменчивости (дифференциации популяций FST и GST) для классических и ДНК маркеров должны быть одинаковы, если выборки репрезентативны.
Масштаб генетической изменчивости популяций юга Центральной России можно выявить лишь при сравнении с другими генофондами. Генетическое разнообразие населения юга Центральной России (![]() =0.0052) оказалось велико в европейском масштабе. Оно не только приближается к уровню генетических различий (
=0.0052) оказалось велико в европейском масштабе. Оно не только приближается к уровню генетических различий (![]() =0.0071, d.f.=20) между всеми популяциями целого народа Западной Европы (немцев, португальцев, ирландцев, норвежцев и др.), но и превышает генетическую изменчивость многих народов Европы: англичан (
=0.0071, d.f.=20) между всеми популяциями целого народа Западной Европы (немцев, португальцев, ирландцев, норвежцев и др.), но и превышает генетическую изменчивость многих народов Европы: англичан (![]() =0.0015, d.f.=32), португальцев (
=0.0015, d.f.=32), португальцев (![]() =0.0015, d.f.=15), датчан (
=0.0015, d.f.=15), датчан (![]() =0.0023, d.f.=30), шведов (
=0.0023, d.f.=30), шведов (![]() =0.0026, d.f.=36), болгар (
=0.0026, d.f.=36), болгар (![]() =0.0022, d.f.=11), чехов (
=0.0022, d.f.=11), чехов (![]() =0.0027, d.f.=7), поляков (
=0.0027, d.f.=7), поляков (![]() =0.0031, d.f.=26) (Балановская и др., 2006).
=0.0031, d.f.=26) (Балановская и др., 2006).
Изученные нами популяции юга Центральной России составляют небольшую часть русского этноса. Априорно юг Центральной России должен представлять очень небольшую часть от генетической изменчивости русского генофонда в целом, так как он охватывает небольшой круг территориально близких популяций (в среднем на расстоянии 125 км). Для сравнения мы использовали данные по селективно-нейтральному уровню изменчивости русского генофонда в целом (Балановская и др., 2006), оцененного по частотам 44 аллелей 17 локусов, который составил ![]() =0.020.
=0.020.
Вопреки исходному предположению, разнообразие популяций юга Центральной России на общем фоне русского генофонда оказалось весьма значимым – на популяции юга Центральной России приходится около 25% всего разнообразия русского генофонда, охватывающего всю изменчивость от казаков Черного моря до поморов Белого моря, от Смоленска на западе до Вятки на востоке. Столь значительная доля – четверть всего генетического разнообразия русского народа - свидетельствует о том, что мы в своей работе, посвященной изучению населения юга Центральной России, анализируем немалую часть изменчивости русского генофонда. Наши данные согласуются с результатами, полученными Е.В. Балановской и др. (2006) при изучении «фамильных портретов» пяти русских регионов – Западного, Центрального, Восточного, Северного и Южного. Авторы установили, что Южный регион России, представленный в работе Белгородской, Курской и Воронежской областями, характеризуется ярко выраженным своеобразием как по спектру, так и по частотам фамилий среди всех других рассмотренных регионов России.
3. Таксономический анализ популяций юга Центральной России
На основе методов многомерной статистики (кластерный анализ, многомерное шкалирование, факторный анализ) проведено исследование генетических соотношений пяти изученных нами районных популяций и двух ранее изученных популяций (Прохоровский и Красненский районы Белгородской области) юга Центральной России (Лепендина, 2005). Показано, что семь рассматриваемых районных популяций юга Центральной России образуют два четко дифференцирующихся кластера. Первый кластер - «юго–западный» - сформирован популяциями Белгородской области, Прохоровского района Белгородской области и Пристенского района Курской области (граничит с Белгородской областью). Второй кластер- «северо–восточный» - образован популяциями Орловской области, Черемисиновского района Курской области (граничит с Орловской областью), Воронежской области и Красненского района Белгородской области (граничит с Воронежской областью) (рис.3). Такое объединение популяций обнаруживается всеми методами многомерной статистики – кластерным анализом, многомерным шкалированием, факторным анализом, что подтверждает реальность выявленных кластеров. При этом расположение популяций в генетическом пространстве (рис.4) хорошо соответствует их

Рис. 3. Расположение в пространстве трех главных факторов семи изученных популяций юга Центральной России (ось Х – первый фактор; ось Y – второй фактор; ось Z – третий фактор) (по частотам 29 аллелей 11 иммуно-биохимических локусов)
Рис. 4. Схема географического расположения семи изученных популяций юга Центральной России (овалами обведены популяции, которые формируют кластеры при кластерном анализе, многомерном шкалировании и факторном анализе)
реальному географическому расположению. Коэффициент корреляции между матрицами генетических и географических расстояний составляет =0.47 (p<0.05). Граница выявленной генетической дифференциации проходит по территории Курской области по линии «юго-запад – северо-восток» и согласуется с изменчивостью ряда аллелей (АВО*А, PGM1*1S, PGM1*1F, ACP1*С).
Установленная нами дифференциация популяций юга Центральной России на два кластера хорошо согласуется с историей формирования населения этого региона. Так, согласно историческим фактам (Восточные славяне, 2002), в X –XIII вв. территория современной Белгородской области и Пристенского района Курской области была заселена племенами северян. Территорию современной Воронежской области заселяли вятичи и племена славян, переселившихся из Донского бассейна в Рязанское поочье.
4. Генетические соотношения населения юга Центральной России с другими русскими популяциями
По единому спектру классических маркеров (22 аллеля 9 локусов) изучены соотношения генофонда юга Центральной России с другими русскими популяциями: московской, курской, белгородской, рязанской, вологодской и тверской (Спицын и др.,2001; Шнейдер и др., 1994, 2002; Лепендина, 2005). Выявлено (рис.5), что популяции «юго-западного» кластера генетически близки к населению Московской и Курской областей, а популяции «северо-восточного» кластера имеют минимальные генетические расстояния от населения Рязанской области. При этом население юга Центральной России генетически удалено от популяций Тверской и Вологодской областей. Полученные данные согласуются с историческими фактами (Загоровский, 1998), согласно которым в XVII-XVIII вв. территорию современной юго-западной части Курской области и западной части Белгородской области заселяли жители Московского региона (ссыльные из Москвы и замосковных городов). За счет переселенцев из Рязанского региона в XVI в. было сформировано население г.Ливны, а затем в конце XVIII века - население современного Черемисиновского района Курской области.
Рис.5. График трехмерного шкалирования, отражающий расположение в пространстве популяций юга Центральной России и некоторых русских популяций (по частотам 22 аллелей 9 иммуно-биохимических локусов): 1- Белгородская область; 2- Орловская область; 3- Воронежская область; 4- Курская область (Пристенский район); 5- Курская область (Черемисиновский район); 6- Белгородская область (Прохоровский район); 7- Белгородская область (Красненский район); 8-Московская область; 9- Рязанская область; 10- Вологодская область; 11- Тверская область; 12- Курская область
Итак, выявлена четкая дифференциация населения юга Центральной России на два кластера («юго-западный» и «северо-восточный») и различные соотношения кластеров с другими русскими популяциями. Можно выдвинуть гипотезу, что такая структура популяций задана историческими факторами - особенностями формирования населения этих территорий в X-XVIII веках: с разным славянским субстратом, лежащим в основе населения этих территорий; разнонаправленностью миграционных потоков различных славянских племен, а затем и разных территориальных групп русских в эти популяции.
Важно подчеркнуть, что генетическая дифференциация популяций юга Центральной России на два кластера не исчезает, а полностью сохраняется и при включении в анализ популяций других языковых семей (рис.6), что свидетельствует о репрезентативности полученных данных. При этом на фоне удаленности от популяций алтайской языковой семьи (якуты, монголы, чуваши) население юга и центра Центральной России формирует общий суперкластер с популяциями уральской языковой семьи (карелы, удмурты, коми), русскими популяциями Вологодской и Тверской областей, популяциями северокавказской (адыгейцы) и индоевропейской (осетины) языковых семей. Полученные результаты могут свидетельствовать о
Рис. 6. Дендрограмма генетических соотношений между популяциями юга Центральной России, некоторыми русскими популяциями и популяциями, относящимися к другим языковым семьям (построена методом Уорда по частотам 22 аллелей 9 иммуно-биохимических локусов)
значимой, хотя и не столь выраженной (в сравнении с миграционными потоками различных территориальных групп русских), роли финно-угорского субстрата, скифо-сарматских корней, тюркского и понтийского влияний на формирование генофонда, как населения юга Центральной России, так и русского этноса в целом.
5. Положение генофонда населения юга Центральной России в популяционной системе восточных славян
С целью оценки места генофонда популяций юга Центральной России в системе восточнославянского генофонда было проведено исследование генетических соотношений коренного русского населения юга Центральной России со «среднерусской» популяцией (для анализа взяты средние частоты изучаемых иммуно-биохимических маркеров для русского генофонда из работ В.А. Спицына и др. (2001) и Е.В. Балановской и др., (2001, 2006) коренным восточным (Белгородская область), центральным (Хмельницкая область) и западным (Львовская область) украинским населением и коренным населением Белоруси (Брестская и Витебская области) (Лепендина, 2005) (рис.7). Установлено, что популяции «юго-западного» кластера имеют ожидаемые минимальные генетические расстояния до «среднерусской» популяции, до территориально близких к ним украинцев Белгородской области. Однако они оказались также генетически близки к украинцам Львовской области. Этот результат подтверждается также данными по изучению гаплогрупп мтДНК и Y хромосомы (Пшеничнов и др., 2006). Это может свидетельствовать о значимом и существенном влиянии мигрантов из Западной Украины на формирование населения юга Центральной России. Наряду с этим, для населения «северо-восточного» кластера юга Центральной России отмечаются минимальные генетические расстояния до украинцев Хмельницкой области. Все популяции белорусов оказались генетически далеки от населения юга Центральной России.
Полученные материалы могут свидетельствовать, во-первых, о значимой и различной соотносительной роли разных групп украинцев в формировании двух территориальных групп русских юга Центральной России в XVII-XVIII веках. Во-вторых, показывают минимальное влияние белорусов на генофонд населения юга Центральной России. Установленные особенности генетических соотношений вышерассмотренных популяций могут быть объяснены известными историческими фактами (Загоровский, 1968,1981,1998), которые свидетельствуют о более значимом и существенном влиянии украинцев на формирование популяций юга Центральной России по сравнению с белорусами.
Рис. 7. График многомерного шкалирования, отражающий расположение в пространстве «северо-западного» и «юго-восточного» кластеров населения юга Центральной России, «среднерусской» популяции и популяций украинцев и белорусов (построен по частотам 25 аллелей 10 иммуно-биохимических локусов)
Обозначения: - белорусская этническая группа; - украинская этническая группа; - русская этническая группа; - изучаемые русские
популяции юга Центральной России
ВЫВОДЫ
1. Дана характеристика структуры генофонда русских популяций четырех областей юга Центральной России (5 популяций, 493 человека) по данным о распределении частот 29 аллелей 11 иммуно-биохимических маркеров: АВО, RH, HP, TF, GC, С’3, ACP1, PGM1, ESD, GLO1, 6-PGD.
2. Генетическая изменчивость населения юга Центральной России (=0.0052) велика в европейском генетическом масштабе: она составляет 25% изменчивости всего русского народа и превышает генетическое разнообразие многих народов Европы.
3. Население юга Центральной России дифференцируется на два кластера – «юго-западный» и «северо-восточный». Выявленные кластеры популяций характеризуются своеобразием генетических соотношений с русскими популяциями и позволили обосновать гипотезу о различном славянском субстрате юго-западных и северо-восточных популяций региона.
4. Население юга Центральной России имеет минимальные генетические расстояния до «среднерусской» популяции и украинцев, но генетически удалено от популяций белорусов.
5. Генетические и географические расстояния между изученными популяциями юга Центральной России обнаружили высокую и достоверную связь (коэффициент корреляции =0.47, р<0.05).
6. Генетическое разнообразие населения юга Центральной России, оцененное по иммуно-биохимическим маркерам (=0.0052), полностью соответствует литературным данным по аутосомным ДНК маркерам (=0.0059) и квазигенетическим маркерам (=0.0062), полученным для этих же популяций.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ:
1. Чурносов М. И., Сокорева И. Н., Костоглодова И. Н., Песик В. Ю., Жерлицына М. С. К вопросу о генетических характеристиках Белгородской популяции. V Бунаковские чтения «Теория антропологии и ее методы. Истоки и развитие». Тезисы докладов. М., 2001. Ч.2. С. 62-63.
2. Чурносов М. И., Костоглодова И. Н.,Сорокина И. Н., Жерлицына М.С., Балановская Е. В. Анализ популяционной структуры Белгородской области по комплексу биохимических и «квазигенетических» маркеров. Материалы международной конференции «Антропология на пороге III тысячелетия (итоги и перспективы)». М., 2003. С.443-453.
3. Жерлицына М. С., Аристова И. К., Чурносов М. И. Изучение распределения фенотипических и генных частот системы GLO1 в русских популяциях юга Центральной России. Третьи антропологические чтения к 75-летию со дня рождения В. П. Алексеева «Экология и демография человека в прошлом и настоящем», М., 2004. С. 253.
4. Жерлицына М. С., Аристова И. К. Биохимический полиморфизм НР в русских популяциях юга Центральной России. Молодежная наука и современность. Материалы 69-й научной конференции студентов и молодых ученых. Курск, 2004. С. 51-52.
5. Чурносов М. И., Балановская Е. В., Сорокина И. Н., Костоглодова И. Н., Песик В. Ю., Рудых Н. А., Жерлицына М. С. Анализ структуры генофонда населения Белгородской области. Экономическая интеграция Белоруси, России и Украины: опыт, проблемы, перспективы. Материалы II Международного Экономического форума славянских народов. Белгород, 2004. Ч.2. С. 199-202.
6. Чурносов М. И., Песик В. Ю., Рудых Н. А., Жерлицына М. С., Сорокина И. Н., Костоглодова И. Н., Аристова И. К., Соловьева Д. С., Балановская Е. В. Комплексное изучение структуры генофонда популяций юга Центральной России. Тезисы III съезда ВОГиС «Генетика в XXI веке: современное состояние и перспективы развития». М., 2004. Т.2. С.146.
7. Чурносов М. И., Сорокина И. Н., Костоглодова И. Н., Песик В. Ю., Рудых Н. А., Аристова И. К., Жерлицына М. С., Балановская Е. В. Структура генофонда населения Белгородской области: квазигенетические, биохимические, молекулярно-генетические и популяционно-демографические параметры. Третьи антропологические чтения к 75-летию со дня рождения В. П. Алексеева «Экология и демография человека в прошлом и настоящем». М., 2004. С.289-290.
8. Жерлицына М. С., Аристова И. К., Костоглодова И. Н. Оценка генетических взаимоотношений русских популяций юга Центральной России. Научные ведомости БелГУ. Белгород, 2005. №.3 (23), вып.4. Серия «Медицина». С. 278-281.
9. Чурносов М. И., Балановская Е. В., Сорокина И. Н., Лепендина И.Н., Песик В. Ю., Рудых Н. А., Аристова И. К., Цапкова Л. А., Ващилин В. С., Жерлицына М. С. Популяционно-генетические характеристики населения Центральной России. Материалы юбилейной научной конференции КГМУ и сессии Центрально-Черноземного научного центра РАМН, посвященной 70-летию КГМУ. Курск, 2005. Т.1. С.189-190.
10. Аристова И. К., Жерлицына М. С., Лепендина И.Н., Чурносов М.И., Балановская Е. В. Полиморфизм русских популяций Центральной России по локусу PGM1. Медицинская генетика. 2005. Т.4. №4. С. 148.
11. Жерлицына М. С., Аристова И. К., Лепендина И. Н., Балановская Е. В., Чурносов М. И. Полиморфизм систем групп крови АВО и Rhesus в десяти русских популяциях Центральной России. Медицинская генетика. 2005. Т.4. № 4. С.183.
12. Чурносов М. И., Песик В. Ю., Рудых Н. А., Сорокина И. Н., Цапкова Л. А., Ващилин В. С., Лепендина И.Н., Аристова И. К., Жерлицына М.С., Балановская Е. В. Материалы по изучению структуры генофонда русского населения Центральной России. Медицинская генетика. 2005. Т.4. № 6. С.289.
13. Аристова И.К., Жерлицына М.С., Лепендина И.Н., Чурносов М.И., Балановская Е.В. Изучение распределения частот фенотипов и генов локуса 6-PGD в русских популяциях Центральной России. Сборник трудов 71-й научной конференции КГМУ и сессии Центрально-Черноземного научного центра РАМН. Курск, 2006. Т.1. С.143-144.
14. Аристова И.К., Жерлицына М.С., Лепендина И.Н., Чурносов М.И., Балановская Е.В. Анализ полиморфизма локусов НР и GC в русских популяциях Центральной России. Сборник материалов Российской научной конференции с международным участием «Медико-биологические аспекты мультифакториальной патологии». Курск, 2006. Т.1. С.9-12.
15. Чурносов М. И., Сорокина И. Н., Лепендина И.Н., Аристова И. К., Жерлицына М.С., Песик В. Ю., Рудых Н. А., Цапкова Л. А., Ващилин В. С., Балановская Е. В. Описание структуры генофонда русского населения юга Центральной России. Медицинская генетика. 2006. Т.5. № 6. С.16-20.