

Базидиальные макромицеты юга амуро-зейского междуречья
На правах рукописи
Кочунова Наталья Анатольевна
БАЗИДИАЛЬНЫЕ МАКРОМИЦЕТЫ
ЮГА АМУРО-ЗЕЙСКОГО МЕЖДУРЕЧЬЯ
03.00.05 – ботаника
03.00.24 – микология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Владивосток - 2007
Работа выполнена в лаборатории защиты растений Амурского филиала Ботанического сада-института ДВО РАН
| Научные руководители: | доктор биологических наук, старший научный сотрудник Васильева Лариса Николаевна кандидат биологических наук Булах Евгения Мироновна |
| Официальные оппоненты: | доктор биологических наук, старший научный сотрудник Егорова Лина Николаевна кандидат биологических наук, доцент Назарова Маргарита Михайловна |
| Ведущая организация: | Тихоокеанский институт географии ДВО РАН |
Защита состоится 16 октября 2007 г. в 1400 часов на заседании диссертационного совета Д 005.003.02 при Биолого-почвенном институте ДВО РАН по адресу: 690022, г. Владивосток, Проспект 100 лет Владивостоку, 159. факс: 8 (4232) 310-193
С диссертацией можно ознакомиться в Центральной научной библиотеке ДВО РАН
Автореферат разослан "___" сентября 2007 г.

Ученый секретарь
диссертационного совета,
доктор биологических наук В.Ю. Баркалов
Общая характеристика работы
Актуальность темы. "Одна из главных особенностей жизни - круговорот органических веществ, основанный на постоянном взаимодействии противоположных процессов синтеза и деструкции" (Камшилов, 1979, c. 33). Грибы, являясь составным компонентом практически всех экосистем, активно участвуют в процессах биодеструкции, в ходе которых происходит возврат биогенных веществ в циклы. Ведущая роль в разложении органического материала принадлежит базидиальным макромицетам – класс Basidiomycetes (Частухин, Николаевская, 1969).
Несмотря на то, что науке уже известно около 100 тысяч видов грибов, до настоящего времени эти организмы остаются одной из наименее изученных групп живых существ. Поэтому проведение исследований в сфере биологического разнообразия грибов остается актуальным. Необходимым этапом таких работ является изучение региональных микобиот, поскольку отсутствие сведений о них делает невозможной оценку степени новизны находок и частоты встречаемости того или иного вида на исследуемой территории
В Амурской области до наших исследований планомерным изучением биоразнообразия макромицетов, в частности базидиальных, специально никто не занимался. Имеющиеся сведения по данной группе грибов немногочисленны и разрознены. Юг Амуро-Зейского междуречья является одной из наиболее освоенных и заселенных частей Приамурья, но стационарные микологические исследования проводились здесь только в окрестностях с. Климоуцы (Томилин, 1969). В условиях активного антропогенного преобразования экосистем особую значимость приобретает выявление природного разнообразия грибов и специфических микологических группировок, адаптированных к конкретным экологическим условиям.
Цель исследования - изучить биоту базидиальных макромицетов юга Амуро-Зейского междуречья.
Задачи исследования:
- выявить видовой состав базидиальных макромицетов юга Амуро-Зейского междуречья;
- провести комплексный анализ (таксономический, экологический, географический) исследуемой микобиоты;
- выявить съедобные, ядовитые и лекарственные виды; дать рекомендации по использованию грибов с полезными свойствами;
- установить редкие и нуждающиеся в охране виды базидиомицетов района исследования; предложить меры их охраны.
Научная новизна. Впервые проведена инвентаризация видового состава биоты базидиальных макромицетов подзоны хвойно-широколиственных лесов юга Амуро-Зейского междуречья, основанная на собственных сборах, а также на анализе литературных данных и образцов микологических гербариев Биолого-почвенного института ДВО РАН (VLA) и Ботанического института им. В.Л. Комарова РАН (LE). Составленный конспект микобиоты содержит 392 вида, из которых 108 являются новыми для территории Амурской области, 9 указываются впервые для российского Дальнего Востока и 5 видов приводятся впервые для территории России. Проанализированы особенности таксономической, географической и эколого-трофической структур микобиоты. Выявлены редкие и нуждающиеся в охране виды грибов.
Практическая ценность. Результаты исследования могут быть использованы для составления кадастров видов и определителей, написания Красных книг регионального и общероссийского уровней. Материалы по дереворазрушающим базидиомицетам Амурской области легли в основу составления коллекций в Институте леса Дальневосточного государственного аграрного университета и Благовещенского государственного педагогического университета и используются в образовательном процессе на практических занятиях по фитопатологии и систематике низших растений.
Апробация работы. Основные результаты исследования докладывались на заседаниях Ученого Совета Амурского филиала Ботанического сада-института ДВО РАН; четырех региональных конференциях молодых ученых (Благовещенск, 2000, 2001, 2002, 2003); XI съезде Русского ботанического общества (Новосибирск-Барнаул, 21-22 августа 2003 г.); VII и IX Дальневосточной молодежной школе-конференции по актуальным проблемам химии и биологии (МЭС ТИБОХ ДВО РАН, 15-22 сентября 2003 г и 16-23 сентября 2005 г.); научно-практической конференции «Новиковские чтения» (Благовещенск, 25 октября 2003 г.), VIII молодежной конференции ботаников (Санкт-Петербург, 17-21 мая 2004 г.) и на I (X) Международной конференции ботаников (Санкт-Петербург, 21-26 мая 2006 г.); отделе ботаники Биолого-почвенного института ДВО РАН (май 2007 г.).
Публикации. По теме диссертации опубликовано 18 работ.
Объем и структура диссертационной работы. Диссертация состоит из введения, 8 глав, выводов, списка цитируемой литературы, включающего 327 источников (195 отечественных и 132 зарубежных), и 4 приложений. Текст изложен на 264 страницах, иллюстрирован 14 таблицами и 15 рисунками.
Содержание работы
Введение
Во введении обоснована актуальность выбранной темы, сформулированы цель и основные задачи исследования, ее научная новизна и практическая ценность.
Глава 1. Природные условия района исследования
Исследование микобиоты проводилось на южном стыке Амуро-Зейской и Зейско-Буреинской равнин между 50°13' и 51°42' с.ш. и 127° и 128° в.д. в пределах Благовещенского и Свободненского административных районов Амурской области. Площадь исследованного района составляет 10,4 тыс. км2.
По последней схеме физико-географического районирования (Рациональное природопользование…, 1989) исследуемая территория находится в Амуро-Сахалинской стране и относится к Амуро-Зейской области, Зейско-Буреинской и Среднезейской провинциям. По ботаническому районированию В.Б. Сочавы (1969) Амуро-Зейское междуречье входит в подзону широколиственно-хвойно-таежных лесов; данной территории соответствует неморальный тип растительности и манчжурская флора.
Глава 2. История изучения базидиальных макромицетов Амурской области
Первые сведения о базидиальных макромицетах Приамурья опубликованы Б.А. Федченко и О.А. Федченко (1906) в списке растений, в котором приводится 10 видов из группы порядков агарикоидных и 1 вид из группы порядков гастероидных базидиомицетов.
Целенаправленные микологические исследования в регионе связаны с именами Б.И. Кравцева и Л.В. Любарского, которые преимущественное внимание уделяли грибам - разрушителям древесины. В 1928 году Б.И. Кравцев выполнил рекогносцировочное обследование дуба монгольского в Свободненском, Шимановском и Благовещенском районах. Для территории Амурской области автором (Кравцев, 1935) указано 45 видов базидиальных макромицетов, вызывающих болезни дуба. Некоторые гербарные образцы были переданы А. Пилату, Б. Бёрту и М.К. Зилинг, которые определили и опубликовали собранный Б.И. Кравцевым материал (Pilat, 1933, 1935, 1936-1942; Burt, 1931; Зилинг, 1936) и несколько дополнили видовой состав. Изучение грибов Дальнего Востока Л.В. Любарский начал в 1929 г. в связи с выявлением зараженности лесов. В Приамурье работа проводилась в лесничествах Магдагачинского, Зейского и Сковородинского районов. Автор в своих публикациях (1936, 1938) отметил сильную обедненность микофлоры изученных районов по сравнению с Приморским краем. В монографии по дереворазрушающим грибам Дальнего Востока (Любарский, Васильева, 1975) для территории Амурской области приводится 131 вид базидиальных макромицетов.
Стационарные исследования микобиоты хвойно-широколиственных лесов проведены сотрудником Ботанического института им. В.Л. Комарова Б.А. Томилиным в 1959 г. в Свободненском районе. В опубликованных работах автором (Томилин, 1962, 1964, 1969) детально проанализированы особенности распределения грибов, в том числе базидиальных, в растительных сообществах.
В 1977-1979 гг. на хр. Тукурингра (Зейский район) была организована экспедиция сотрудников Московского университета. Кроме собственных сборов, они обработали сборы Б.А. Томилина с хребтов Тукурингра и Соктахан, осуществленные в 1965 году. Всего было выявлено 110 видов базидиальных макромицетов (Флора…, 1981; Гордиенко и др., 1990).
Значительный вклад в изучение макромицетов Приамурья внесли сотрудники Биолого-почвенного института ДВО РАН (БПИ ДВО РАН). В 1958-1959 годах Л.Н. Васильевой и М.М. Назаровой проводились исследования в центральной и юго-восточной частях Амурской области; в 1976 г. отдельно М.М. Назаровой - в бассейне р. Селемджа. По итогам этих экспедиций опубликовано несколько статей, в которых приводятся свыше 300 видов базидиальных макромицетов (Васильева, 1960, 1971; Васильева, Назарова, 1962; Назарова, Васильева, 1974; Назарова, 1986). Также сотрудниками института Е.М. Буллах, О.К. Говоровой и М.М. Назаровой был составлен список грибов Хинганского заповедника, включающий 219 видов из класса Basidiomycetes (Азбукина и др., 1998). Информация о гетеробазидиальных и афиллофороидных грибах с территории Амурской области содержится в статьях О.К. Говоровой (1994, 1997, 1998, 1999, 2001, 2002a, b, 2003).
В 1961 г. в Тындинском районе в составе 2-й Дальневосточной комплексной экспедиции работали сотрудники Тартуского университета (Эстония) Э.Х. Пармасто и А.Г. Райтвийр. В опубликованных ими сводках по рогатиковым, кортициевым, гетеробазидиальным и лахнокладиевым грибам СССР (Parmasto, 1967, 1968; Пармасто, 1965, 1970; Райтвийр, 1967; Raitviir, 1971) указывается 43 вида с территории Приамурья.
Некоторые сведения о базидиальных макромицетах Амурской области встречаются в сводках по отдельным группам - родам, семействам и порядкам (Николаева, 1961, 1964, 1967; Давыдкина, 1980; Нездойминого, 1983, 1990, 1996; Бондарцева, Пармасто, 1986; Булах, 1986, 1990; Назарова, 1990; Коваленко, 1989, 1995; Бондарцева, 1998; Wells, Raitviir, 1975, 1977).
Несмотря на достаточно большое количество известных для области видов (около 700), базидиальные макромицеты юга Амуро-Зейского междуречья изучены очень слабо и неравномерно по отдельным группам. К началу наших исследований на данной территории они были представлены 125 видами, относящимися к 84 родам, 40 семействам и 19 порядкам.
Глава 3. Материал и методы исследования
Материалом для настоящей работы послужили собственные сборы автора, осуществленные в период с 2001 по 2006 гг., а также все имеющиеся по изучаемой территории литературные данные и образцы грибов из коллекций Регионального гербария БПИ ДВО РАН и Ботанического института РАН.
Сбор базидиомицетов осуществлялся маршрутным методом в течение вегетационного периода (с мая по октябрь). Максимальный интервал между сборами (10 дней) определялся скоростью и предельным развитием плодовых тел агарикоидных грибов (Малый, 1987; Столярская, 1998). Маршруты составлялись с учетом наиболее полного охвата биогеоценозов, типичных для территории юга Амуро-Зейского междуречья.
В результате полевых работ было собрано около 1500 образцов базидиальных макромицетов, основная часть которых хранится в коллекции Регионального гербария (VLA). Гербарные образцы изготавливались по стандартным методикам (Бондарцев, Зингер, 1950; Гербарное дело, 1995); при определении цветовой гаммы базидиом руководствовались шкалой цветов, разработанной А.С. Бондарцевым (1953).
Идентификация гербарного материала проводилась на базе Амурского филиала Ботанического сада-института ДВО РАН (г. Благовещенск) и Лаборатории низших растений БПИ ДВО РАН с применением микроскопов марок «Laboval-4» и «Amplival». Микроскопирование базидиом производилось с использованием стандартного набора реактивов и красителей (5-7% КОН или NaOH, реактив Мельцера, сульфованилин, сульфат железа, краситель Конго-Красный и др.). Для идентификации грибов взяты общие определители (Moser, 1978; Jlich, 1984; Nordic Macromycetes, 1992, 1997), а также использовались многочисленные монографии и публикации по отдельным группам базидиомицетов.
Для анализа микобиоты применялись математические методы и приемы, принятые в сравнительной флористике и геоботанике (Василевич, 1969; Шмидт, 1984; Зайцев, 1991). При сравнении видовых составов базидиальных макромицетов изучаемой микобиоты и микобиот заповедников Дальнего Востока России использовались стандартные коэффициенты сходства (различия). Построение дендрограммы сходства видового состава сообществ базидиомицетов различных древесных пород осуществлено с помощью компьютерной программы Statistica 6.0 методом взвешенного среднего арифметического связывания (WPGMC).
При анализе трофической структуры использовалась шкала трофических групп А.Е. Коваленко (Коваленко, 1980; Столярская, Коваленко, 1996).
Глава 4. Конспект биоты базидиальных макромицетов юга Амуро-Зейского междуречья
Конспект основан на собственных и некоторых литературных данных, а также на обработке гербарных образцов регионального гербария Биолого-почвенного института ДВО РАН (VLA) и Микологического гербария Ботанического института им. В.Л. Комарова РАН (LE).
Порядки и семейства приводятся по системе, принятой в 8-м издании «Словаря грибов» (Hawksworth et al., 1995), но с некоторыми изменениями. Для удобства порядки расположены в списке не по алфавиту, а в следующей последовательности по группам: гетеробазидиальные, афиллофороидные, агарикоидные и гастероидные базидиомицеты. Последовательность родов внутри семейств и видов внутри родов дается в алфавитном порядке.
Для каждого вида указывается местонахождение (ближайший населенный пункт или географический объект), местообитание, субстрат и дата сбора, номер (если таковой имеется) гербарного образца, хранящегося в коллекции гербария БПИ ДВО РАН (VLA), распространение в России и общее распространение, географический элемент (зональный / региональный). Для наиболее значимых видов приводятся пищевые свойства (съедобный или ядовитый); также в списке отмечены виды, обладающие лекарственными свойствами. Для видов, приведенных в литературных источниках, указываются синонимы.
Номенклатура грибов принята и сверена по Index Fungorum (сайт http//www.mykoweb.com). Сокращения имен авторов даны в соответствии с публикацией П.М. Кирка и А.Е. Энселла (Kirk, Ansell, 1992).
Глава 5. Таксономический анализ микобиоты
В результате проведенных исследований выявлено 392 вида базидиомицетов, относящихся к 149 родам, 59 семействам и 21 порядку (табл. 1). Новыми для территории Амурской области являются 108 видов, из которых 9 видов - новые для территории Российского Дальнего Востока, а 5 видов (Agrocybe brunneola, Cantharellus borealis, Mycena odorifera, Tricholoma sciodes,Volvariella surrecta) - новые для территории России.
В среднем на порядок приходится по 18,7 видов, что говорит о сравнительно низком видовом разнообразии изученной биоты. Первое место в микобиоте занимает порядок Agaricales, на долю которого приходится 37,76 % от общего числа видов. Также заметный вес имеют порядки Poriales (50 видов), Cortinariales (37 видов), Russulales (28 видов), Boletales (24 вида). Самыми малочисленными порядками, включающими по 2-5 видов, являются Auriculariales, Gomphales, Lachnocladiales, Schizophyllales, Sclerodermatales, Nidulariales и Phallales.
Таблица 1
Таксономическая структура базидиомицетов
юга Амуро-Зейского междуречья
| Порядки, семейства (число родов / видов) | Роды (число видов) | |
| Auriculariales (1/3) | ||
| Auriculariaceae (1/3) | Auricularia (3) | |
| Tremellales (2/7) | ||
| Exidiaceae (1/4) | Exidia (4) | |
| Tremellaceae (1/3) | Tremella (3) | |
| Cantharellales (9/12) | ||
| Cantharellaceae (1/2) | Cantarellus (2) | |
| Clavariaceae (1/1) | Ramariopsis (1) | |
| Clavariadelphaceae (1/2) | Clavariadelphus (2) | |
| Clavulinaceae (1/1) | Clavulina (1) | |
| Craterellaceae (1/1) | Craterellus (1) | |
| Hydnaceae (3) | Climacodon (1), Hydnum (2) | |
| Pterulaceae (1/1) | Pterula (1) | |
| Typhulaceae (1/1) | Pistillaria (1) | |
| Dacrymycetales (4/6) | ||
| Dacrymycetaceae (4/6) | Calocera (1), Dacrymyces (3), Dacryopinax (1), Ditiola (1) | |
| Ganodermatales (1/2) | |
| Ganodermataceae (1/2) | Ganoderma (2) |
| Gomphales (2/5) | |
| Lentariaceae (1/1) | Lentaria (1) |
| Ramariaceae (1/4) | Ramaria (4) |
| Hericiales (5/9) | |
| Auriscalpiaceae (1/1) | Auriscalpium (1) |
| Clavicoronaceae (1/1) | Clavicorona (1) |
| Gloeocystidiellaceae (1/1) | Laxitextum (1) |
| Hericiaceae (1/3) | Hericium (3) |
| Lentinellaceae (1/3) | Lentinellus (3) |
| Hymenochaetales (5/13) | |
| Hymenochaetaceae (5/13) | Coltricia (1), Hymenochaete (2), Inonotus (3), Onnia (1), Phellinus (6) |
| Lachnocladiales (1/2) | |
| Lachnocladiaceae (1/2) | Scytinostroma (2) |
| Poriales (28/50) | |
| Coriolaceae (21/34) | Bjerkandera (2), Cerrena (1), Coriolopsis (1), Daedaleopsis (2), Fomes (1), Fomitopsis (2), Funalia (1), Gloeophyllum (2), Hapalopilus (1), Laetiporus (1), Laricifomes (1), Lenzites (1), Osteina (1), Phaeolus (1), Piptoporus (1), Postia (2), Pycnoporus (2), Spongipellis (2), Trametes (6), Trichaptum (2), Tyromyces (1) |
| Lentinaceae (5/10) | Lentinus (2), Neolentinus (1), Panus (1), Phyllotopsis (1), Pleurotus (5) |
| Polyporaceae (2/6) | Cryptoporus (1), Polyporus (5) |
| Schizophyllales (2/2) | |
| Schizophyllaceae (2/2) | Plicaturopsis (1), Schizophyllum (1) |
| Stereales (10/13) | |
| Corticiaceae (1/1) | Cytidia (1) |
| Hyphodermataceae (1/1) | Cerocorticium (1) |
| Meruliaceae (3/3) | Chondrostereum (1), Gloeoporus (1), Merulius (1) |
| Peniophoraceae (1/2) | Peniophora (2) |
| Sistotremataceae (1/1) | Sistotrema (1) |
| Steccherinaceae (2/3) | Irpex (1), Steccherinum (2) |
| Stereaceae (1/2) | Stereum (2) |
| Thelephorales (4/13) | |
| Thelephoraceae (4/13) | Hydnellum (3), Thelephora (4), Tomentella (5), Tomentellopsis (1) |
| Agaricales (45/148) | |
| Agaricaceae (5/20) | Agaricus (6), Cystoderma (2), Lepiota (8), Leucocoprinus (1), Macrolepiota (3) |
| Amanitaceae (1/9) | Amanita (9) |
| Bolbitiaceae (2/5) | Agrocybe (4), Conocybe (1) |
| Coprinaceae (2/13) | Coprinus (9), Psathyrella (4) |
| Entolomataceae (2/4) | Entoloma (3), Rhodocybe (1) |
| Hygrophoraceae (2/7) | Hygrocybe (2), Hygrophorus (5) |
| Pluteaceae (2/9) | Pluteus (5), Volvariella (4) |
| Strophariaceae (5/16) | Hypholoma (2), Panaeolus (2), Pholiota (9), Pleuroflammula (1), Stropharia (2) |
| Tricholomataceae (24/65) | Armillariella (1), Arrhenia (1), Calocybe (2), Clitocybe (9), Collybia (2), Crinipellis (1), Dermoloma (1), Flammulina (1), Gymnopus (3), Hohenbuehelia (3), Laccaria (2), Lepista (4), Lyophyllum (2), Marasmiellus (1), Marasmius (6), Melanoleuca (1), Mycena (8), Ossicaulis (1), Panellus (3), Pseudoclitocybe (1), Resupinatus (1), Tricholoma (8), Tricholomopsis (1), Xeromphalina (2) |
| Boletales (11/24) | |
| Boletaceae (4/13) | Boletus (2), Chalciporus (1), Leccinum (4), Suillus (6) |
| Gomphidiaceae (1/1) | Chroogomphus (1) |
| Gyrodontaceae (2/5) | Boletinus (4), Psiloboletinus (1) |
| Hygrophoropsidaceae (1/1) | Hygrophoropsis (1) |
| Paxillaceae (1/1) | Paxillus (1) |
| Strobilomycetaceae (1/1) | Porphyrellus (1) |
| Xerocomaceae (1/2) | Xerocomus (2) |
| Cortinariales (7/37) | |
| Cortinariaceae (6/33) | Cortinarius (11), Galerina (2), Gymnopilus (4), Hebeloma (12), Inocybe (12), Naucoria (1) |
| Crepidotaceae (1/4) | Crepidotus (4) |
| Russulales (2/28) | |
| Russulaceae (2/28) | Lactarius (11), Russula (17) |
| Lycoperdales (4/11) | |
| Geastraceae (1/2) | Geastrum (2) |
| Lycoperdaceae (3/9) | Bovista (2), Handkea (1), Lycoperdon (6) |
| Nidulariales (2/2) | |
| Nidulariaceae (2/2) | Cyatus (1), Nidularia (1) |
| Phallales (2/2) | |
| Phallaceae (2/2) | Dictyophora (1), Mutinus (1) |
| Sclerodermatales (2/3) | |
| Astraeceae (1/1) | Astraeus (1) |
| Sclerodermataceae (1/2) | Scleroderma (2) |
| 21 порядок, 59 семейств | 149 родов, 392 вида |
Ведущие семейства - Tricholomataceae, Coriolaceae, Cortinariaceae, Russulaceae, Agaricaceae, Strophariaceae - являются характерными для микобиот умеренной Голарктики (Морозова, 2001; Перова, Горбунова, 2001; Сопина, 2001; Ставишенко, Мухин, 2002), но это не показывает своеобразия микобиоты исследованной территории. Гораздо более информативным оказалось ее сравнение с микобиотами ряда заповедников российского Дальнего Востока - по количеству видов и так называемым "пропорциям флоры" (табл. 2).
Таблица 2
Показатели таксономического богатства и
разнообразия («пропорции флоры») сравниваемых микобиот
| Микобиоты | Число | Пропорции флоры | ||||||
| видов | родов | семейств | порядков | в/р | в/с | р/с | с/п | |
| А-З | 392 | 149 | 59 | 21 | 2,63 | 6,64 | 2,53 | 2,8 |
| З | 266 | 118 | 46 | 19 | 1,91 | 5,78 | 2,57 | 2,42 |
| Х | 283 | 118 | 46 | 18 | 2,4 | 6,15 | 2,57 | 2,56 |
| К | 357 | 130 | 47 | 17 | 2,75 | 7,6 | 2,77 | 2,76 |
| Бх | 580 | 185 | 55 | 17 | 3,14 | 10,55 | 3,36 | 3,24 |
| У | 883 | 227 | 67 | 20 | 3,89 | 13,2 | 3,39 | 3,35 |
| Кп | 812 | 226 | 72 | 23 | 3,6 | 11,27 | 3,14 | 3,13 |
| Л | 678 | 227 | 74 | 21 | 2,99 | 9,16 | 3,07 | 3,52 |
| М | 154 | 69 | 27 | 12 | 2,23 | 5,7 | 2,6 | 2,25 |
Условные обозначения: А-З – Амуро-Зейское междуречье; заповедники: З – Зейский, Х – Хинганский, К - Комсомольский, Бх – Большехехцирский, У – Уссурийский, Кп – Кедровая падь, Л – Лазовский, М – Магаданский.
«Пропорции флоры» или «коэффициенты насыщенности» - это показатели таксономического разнообразия: среднее число видов в роде (в/р), среднее число видов в семействе (в/с), среднее число родов в семействе (р/с), среднее число семейств в порядке (с/п). Самым достоверным показателем из выше перечисленных считается коэффициент видовой насыщенности родов (в/р), который «является наиболее независимым от площади территорий отношением» (Шмидт, 1984). Однако в нашем анализе более достоверным оказался коэффициент видовой насыщенности семейств, так как его значения гораздо более отчетливо разделяют микобиоты на две группы - "богатые" (от 9.16 до 11.27) и "бедные" (от 5.7 до 7.6).
Микобиота юга Амуро-Зейского междуречья занимает 5-е место по количеству видов, но оказывается в "бедной" разнообразием группе, хотя данная территория относится к той же манчжурской (восточно-азиатской) флоре, что и "богатые" заповедники (Уссурийский, Кедровая падь, Лазовский и Большехехцирский). Это, очевидно, связано с характером лесной растительности междуречья, где набор доминирующих пород ограничен (дуб, береза, сосна, лиственница). Кроме того, они образуют довольно однообразные сообщества (дубово-сосновые и дубово-лиственничные леса, дубняки, сосняки, березняки) в отличие от полидоминантных лесов четырех указанных заповедников.
Низкие значения коэффициентов Жаккара, Серенсена-Чекановского и Стугрена-Радулеску (табл. 3.) также выявляют заметное преобладание различия над сходством микобиот.
Таблица 3
Коэффициенты сходства видового состава микобиот заповедников с биотой базидиальных макромицетов Амуро-Зейского междуречья
| Коэффициенты | Значения | |||||||
| З | Х | К | Бх | У | Кп | Л | М | |
| Жаккара, % | 30,0 | 21,6 | 28,9 | 27,7 | 25,9 | 25,0 | 26,5 | 19,7 |
| Стугрена-Радулеску | 0,40 | 0,57 | 0,42 | 0,45 | 0,48 | 0,49 | 0,42 | 0,61 |
| Серенсена-Чекановского, % | 46,2 | 35,5 | 44,9 | 43,4 | 41,2 | 40,0 | 41,9 | 32,9 |
Если дополнить количественное сравнение качественным, то опять выступает таксономическое своеобразие микобиоты Амуро-Зейского междуречья. В частности, 41 вид характерен только для изученной биоты и не зарегистрирован на территории рассматриваемых заповедников. Еще 46 видов найдены лишь в одном из них (12 отсутствуют в "манчжурских" микобиотах). Наконец, 57 видов отмечены еще в двух из заповедников, т. е. около 37 % видового состава микобиоты юга Амуро-Зейского междуречья - это виды сравнительно редкие на Дальнем Востоке России или отсутствующие в других его районах.
Глава 6. ЭКОЛОГический анализ микобиоты
В эколого-ценотическом анализе рассматривается распределение видов базидиальных макромицетов по местообитаниям и эколого-трофическим группам, а также их приуроченность к древесным породам.
В пределах территории Амуро-Зейского междуречья нами исследованы 19 типов местообитаний (это – различные типы лесных фитоценозов, пойменные экотопы, луга и рудеральные местообитания), которые отличаются качественно и количественно по видовому составу базидиальных макромицетов. Наибольшее количество видов отмечено в березово-дубово-сосновых (191), березово-дубовых (156), дубово-сосновых лесах (133), которые являются наиболее распространенными на юге Амуро-Зейского междуречья. Незначительно уступают по количеству видов чистые дубняки (106) и березово-дубовые леса с примесью ольхи (101). Специфическим видовым составом отличаются осиновые леса (осинники), в которых представлено всего 11 видов базидиальных макромицетов.
Для лугов и рудеральных местообитаний характерно небольшое число видов (25 и 33 соответственно), преимущественно из семейств Agaricaceae и Coprinaceae, которые, тем не менее, входят в состав ведущих семейств изученной группы грибов. Это говорит о засоренности микобиоты рудеральными видами, а присутствие в лесах такого вида как Coprinus comatus служит индикатором антропогенной нагрузки на природные экосистемы (Булах, 1977).
Все обнаруженные на исследуемой территории виды по типу питания отнесены нами к трем экологическим группам: сапротрофам (66,85 %), симбиотрофам (29,8 %) и факультативным паразитам (3,35 %). Такое соотношение трофических групп соответствует закономерности, установленной для всех региональных микобиот лесных экосистем Голарктики (Бурова, 1986).
К симбиотрофным грибам относится группа микоризообразователей. Нами на территории междуречья выявлен 121 вид грибов, способных образовывать симбиоз с корнями древесных растений, в том числе и факультативные микоризообразователи. Больше всего видов грибов-симбиотрофов принадлежит к агарикоидным базидиомицетам, а именно все представители семейств Amanitaceae, Russulaceae, Boletaceae; многие виды семейств Tricholomataceae, Cortinariaceae, Hygrophoraceae; некоторые представители семейств Cantharellaceae, Hydnaceae, Thelephoraceae, Lycoperdaceae.
На исследуемой территории выявлено небольшое число грибов, избирательно паразитирующих на древесных породах (30 видов). Наибольший процент паразитных базидиомицетов приходится на семейства Coriolaceae и Hymenochaetaceae. На территории междуречья отмечена высокая зараженность растущих деревьев дуба такими грибами, как Spongipellis delectans, Hericium erinaceum и Armillariella mellea.
Сапротрофы в свою очередь по типу питающего субстрата (при этом нами не учитывалась степень разложения древесины) подразделяются на 10 трофических групп. Общее соотношение эколого-трофических групп в изученной микобиоте показано на рисунке 1.
Наибольшее число сапротрофных видов относится к ксилотрофам. Хотя потенциально на мертвой древесине могут развиваться 169 видов от общего количества выявленных нами, но в условиях района исследования обнаружено на древесном субстрате 140 видов (35,71 %). В основном это представители афиллофороидных базидиомицетов преимущественно из порядков Poriales и Hymenochaetales.
Гумусовые сапротрофы не обладают узкой приуроченностью к субстрату (Васильева, 1973; Вассер, 1980) и растут как в лесах, так и вне леса на открытых пространствах. На почве нами обнаружено 62 вида (15,82 %), в основном это представители семейств Agaricaceae, Tricholomataceae и Coprinaceae.
Подстилочные сапротрофы и сапротрофы на опаде, в отличие от гумусовых, обладают тесными связями с лесными экосистемами. Им принадлежит ведущая роль в разложении целлюлозы и лигнина растительных остатков - листьев, хвои, веточек, шишек, кусочков коры. К подстилочным сапротрофам принадлежит 32 вида (8,2 %), а к «опадным» - 15 видов (3, 83 %). Например, на подстилке и опаде обнаружены Gymnopus confluens, Marasmius scorodonius, Mycena pura, Pluteus leoninus, Cystoderma granulosum, Xeromphalina cauticinalis, Sistotrema confluens, Auriscalpium vulgare, Lentaria dendroidea и др.
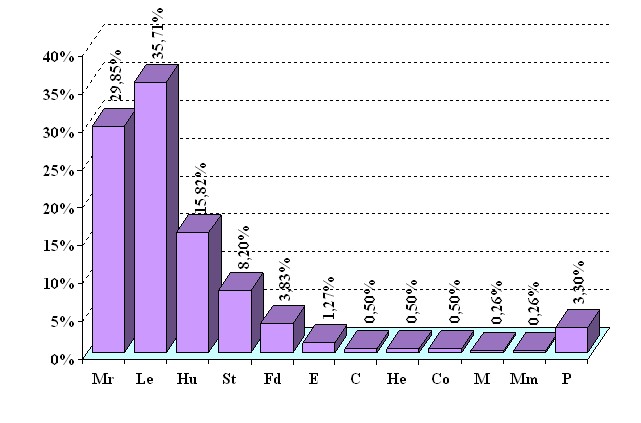
Рис. 1. Распределение базидиальных макромицетов юга
Амуро-Зейского междуречья по трофическим группам
Условные обозначения: Mr – микоризообразователи, Le – сапротрофы на древесине, Hu – гумусовые сапротрофы, St – подстилочные сапротрофы, Fd – сапротрофы на опаде, E – сапротрофы на экскрементах, М – сапротрофы на мхах, Mm – на плодовых телах макромицетов, С – сапротрофы на углях, He – сапротрофы на стеблях травянистых растений, Co – сапротрофы на коре деревьев, P – факультативные паразиты.
Вклад в биоту базидиомицетов Амуро-Зейского междуречья остальных трофических групп невелик и ограничивается 1-5 видами, наиболее специализированными к типу субстрата (на экскрементах, на мхах, на стеблях травянистых растений, на коре деревьев, на углях).
Проведенный анализ распределения видов по трофическим группам не показывает экологического своеобразия микобиоты юга Амуро-Зейского междуречья, но это можно выяснить при совмещении такого анализа с рассмотрением приуроченности макромицетов к определенным древесным породам[1]
.
Большинство базидиомицетов (табл. 4) консортивно связано с основными лесообразующими породами исследуемой территории – дубом (159 видов), березой (108 видов), сосной (76 видов) и лиственницей (41 вид). Из данных таблицы 4 следует, что почти на каждой породе ксилотрофные базидиомицеты преобладают над симбиотрофами, за исключением микоризообразователей сосны, которых численно больше.
Таблица 4
Распределение базидиомицетов района исследования
по эколого-трофическим группам и по древесным породам
| Трофические группы | Д | Б | С | Л | Ил | Ив | Ол | Т | Ос | Ч | К |
| Mr | 59 | 45 | 41 | 16 | 2 | 5 | 7 | 6 | 4 | – | – |
| Le | 85 | 57 | 26 | 19 | 26 | 19 | 16 | 17 | 15 | 12 | 5 |
| Fd | 8 | 4 | 6 | 1 | 2 | 1 | 1 | – | – | 2 | – |
| P | 7 | 2 | 3 | 5 | 4 | 3 | – | 1 | – | 1 | 1 |
| Итого видов: | 159 | 108 | 76 | 41 | 34 | 28 | 24 | 24 | 19 | 15 | 6 |
Условные обозначения: Хвойные породы: С – сосна; Л – лиственница. Лиственные породы: Д – дуб; Б – береза; Ил – ильм; Ив – ива; Ол – ольха; Т – тополь; Ос – осина; Ч – черемуха; К - клен.
Если рассмотреть две из основных пород-эдификаторов лесной растительности юга Амуро-Зейского междуречья - дуб и березу - и сравнить соотношение микоризообразователей и ксилотрофов на этих породах, например, в Уссурийском заповеднике, то можно наблюдать разницу ситуаций.
В изученной микобиоте с дубом связано 59 микоризообразователей и 85 ксилотрофных макромицетов, а в Уссурийском заповеднике микоризообразователи дуба (всего 28 видов; только под дубом - 20, другие также - под березой, лещиной и пихтой) преобладают над ксилотрофами (всего 22 вида, только на дубе - 15). Характерно, что дуб на юге Амуро-Зейского междуречья поддерживает гораздо большее разнообразие макромицетов, чем в самом богатом заповеднике Дальнего Востока (Азбукина и др., 2006). В данном случае, на основной доминирующий вид растительности как бы "стекаются" макромицеты, распределенные по разным породам в полидоминантных лесах.
В изученной микобиоте с березой связано 45 микоризообразователей и 57 ксилотрофов. В Уссурийском заповеднике микоризообразователи данной породы (38 видов) также преобладают над ксилотрофами (16 видов), и разнообразие в "грибной свите" березы также выше на юге Амуро-Зейского междуречья по сравнению с Уссурийским заповедником.
Если большее разнообразие макромицетов, связанных с дубом и березой в изученной микобиоте, объясняется преимущественным доминированием этих пород в лесной растительности, то преобладание ксилотрофов над микоризообразователями может говорить о том, что данные экологические группы находятся в разных микроклиматических условиях. Ксилотрофы чаще заселяют валежные (76 видов) и сухостойные (50 видов) стволы деревьев, пни (44 вида), валежные и сухостойные ветви (40 видов), нежели гнилушки, корни и погребенную в почве древесину, что предполагает ограничивающее влияние напочвенных условий.
Даже в Уссурийском заповеднике, где микоризообразователи дуба и березы преобладают, установлен крайне неравномерный режим увлажнения почвы: в первой половине вегетационного периода тепло и сухо, а во второй - большое количество осадков приводит к переувлажнению почвы (Таранков, 2006). По всей вероятности, в исследованном районе гидротермический режим напочвенного покрова еще более неблагоприятен для развития макромицетов в связи промерзанием почвы зимой и неравномерным сезонным распределением осадков.
Если же сравнить распределение трофических групп на сосне в Амуро-Зейском междуречье (41 вид - микоризообразователи, 26 видов - ксилотрофы), то здесь преобладание микоризообразователей можно объяснить тем, что сосна образует не только сообщества с другими породами, но также чистые сосняки, где микроклимат напочвенного покрова отличается от такового в других лесах. Например, в связи с бедным травяным покровом здесь почти не бывает избыточного переувлажнения почвы. Кроме того, известно, что густота напочвенного покрова отрицательно влияет на развитие плодовых тел агариковых грибов (Томилин,1969; Нездойминого, 1979; Вассер, 1980), а в сосняках этот негативный эффект снимается.
По связям с древесными породами рассматриваемые виды базидиомицетов можно подразделить на стенотрофы (связанные с одной древесной породой) и эвритрофы (связанные с двумя и более породами). В условиях района исследования свыше половины видового состава микоконсортов древесных пород проявляют себя как стенотрофы (148 видов, или 52,3 %). Явление стенотрофности в данном случае носит региональный характер, поскольку большинство видов базидиомицетов данной группы в других частях ареала связано с обширным кругом древесных растений. Это подтверждает высказанный тезис (см. глава 5) о влиянии моно- и олигодоминантных лесных сообществ на характер микобиоты.
Глава 7. Географический анализ микобиоты
Микогеографический анализ проводился согласно регионально-зональному принципу, которому следует большинство микологов (Змитрович, 1998; Столярская, 1998; Лосицкая, 1999; Морозова, 2001; Сопина, 2001; Косолапов, 2004).
Было выделено 5 зональных географических элементов (бореальный - b, неморальный - n, бореонеморальный – bn, панмультизональный - mz и мультизональный лесной – mzf). В рассматриваемой микобиоте преобладают виды, относящиеся к мультизональным элементам, которые в совокупности составляют около половины (187 видов, или 47,82 %) всех выявленных на территории междуречья видов грибов. Существенную долю (102 вида, или 26,09 %) составляют виды бореонеморального геоэлемента (ГЭ), распространенные в лесной полосе Голарктики. Преобладание неморального ГЭ (76 видов, или 19,44 %) над бореальным ГЭ (26 видов, или 6,65 %) подчеркивает неморальный тип растительности, характерный для территории исследования по районированию В.Б. Сочавы (1969).
Региональных географических элементов - 4 (палеарктический - P, голарктический - H, мультирегиональный - MR, космополитный - KOSM). Первое место в ряду региональных элементов занимает группа голарктических видов (162 вида или 41,43 %), общих для Палеарктики и Неарктики, что можно объяснить существованием в прошлом покрытого лесами Берингова моста, через который и распространялись грибы (Пармасто, 1979). Группа космополитных видов занимает второе место (25,58 %) в микобиоте, третье место принадлежит видам (16,62 %) палеарктического элемента.
Одновременное использование широтной (зональный элемент) и долготной (региональный элемент) характеристик носит название принципа "биогеографических координат" (Юрцев, 1968) и позволяет выделить 14 типов ареала для базидиальных макромицетов Амуро-Зейского междуречья (табл. 5).
Ядро биоты базидиомицетов территории исследования составляют виды, имеющие обширные ареалы. Первое место в микобиоте занимают виды с голарктическим бореонеморальным[2]
типом ареала, распространенные в лесной зоне Голарктики (bn/H). На их долю приходится 21,17 %. Второе и третье места занимают виды с космополитным мультизональным лесным (76 видов или 19,39 %) и с мультирегиональным мультизональным лесным (53 вида или 13,52 %) типами ареала. Слагающие эти группы виды встречаются практически на всех континентах в различных природно-климатических зонах, но ввиду своей трофической специализации приурочены к лесным биогеоценозам (сапротрофы на опаде, ксилотрофные и микоризообразующие грибы).
Таблица 5
Распределение видов базидиальных макромицетов района
исследования по географическим элементам и типам ареала
| РГЭ ЗГЭ | P | H | MR | KOSM | |
| b | 3 | 23 | — | — | 26 |
| n | 35 | 41 | — | — | 76 |
| bn | 19 | 83 | — | — | 102 |
| mz | 6 | 12 | 11 | 24 | 53 |
| mzf | 2 | 3 | 53 | 76 | 134 |
| 65 | 162 | 64 | 100 | 391 |
Условные обозначения: РГЭ – региональные географические элементы;
ЗГЭ – зональные географические элементы; – сумма видов.
Заметную долю (10,46 %) составляют виды, распространенные в зоне неморальных лесов Голарктики. Это преимущественно представители агарикоидных грибов (семейства Hygrophoraceae, Pluteaceae, Tricholomataceae, Russulaceae и др.). Следующее место занимают виды, также обитающие в зоне неморальных лесов, но в своем распространении ограниченные Палеарктикой. Их доля в изученной микобиоте составляет 8,93 % (35 видов).
Виды космополитного панмультизонального типа ареала (24 вида или 6,12 %) не имеют строгой приуроченности к субстрату и встречаются на всех континентах в разных природно-климатических зонах, как в лесах, так и вне леса. В эту группу входят, главным образом, сапротрофы на гумусе и экскрементах, факультативные микоризообразователи, рудеральные виды.
Близкой по объему к вышеназванной является группа видов, принадлежащих к голарктическому бореальному (23 вида или 5,87 %) типу ареала. Эта группа включает широко распространенные виды, общие для Палеарктики и Неарктики, обитающие в полосе хвойных лесов таежной зоны. К палеарктическому бореонеморальному типу ареала мы относим 19 видов (4,85 %).
Таким образом, биоту базидиомицетов юга Амуро-Зейского междуречья можно охарактеризовать как эвриголарктическую (голарктическую бореонеморальную) с выраженными в большей степени неморальными чертами.
Хотя проведенный анализ в общем нивелирует особенности распространения макромицетов юга Амуро-Зейского междуречья, его можно использовать как своего рода рамки для обсуждения некоторых конкретных ситуаций, связанных с ареалами, и подтвердить своеобразие изученной микобиоты, которое следовало из таксономического и экологического анализов.
Например, в группе палеарктических видов (65) фактически отсутствуют типично восточноазиатские виды, столь характерные для заповедников Приморья. Исключение составляют только условно эндемичный вид Mycena mellea, распространенный в дубовых лесах Российского Дальнего Востока, а также Psiloboletinus lariceti, связанный с лиственницей от Урала до Тихого океана. Напротив, микобиота исследованного района содержит довольно значительное число видов, общих с европейскими микобиотами.
Для формирования многих локальных микобиот Российского Дальнего Востока имел большое значение географический "берингийский мост" (Petersen, Hughes, 2007), но для генезиса микобиоты юга Амуро-Зейского междуречья, по-видимому, гораздо большее значение имел "экологический мост" в виде ареала сосновых лесов. Этот ареал простирается сплошной полосой от Европы через Сибирь до района исследований - с небольшим разрывом, который, вероятно, был заполнен раньше. По этому "сосновому мосту" могли распространиться до исследованного региона европейские виды, найденные нами впервые в России (Agrocybe brunneola, Tricholoma sciodes) или на территории Дальнего Востока (Cortinarius stillatitius, Dermoloma cuneifolium, Psathyrella olympiana f. amstelodomensis).
Березовые леса широко распространены, и по ним, по всей вероятности, тоже часто «приходили» виды с запада, так как сходство видового состава макромицетов, связанных с березой в восточных микобиотах и на исследованной территории, довольно низкое. Особенно интересные "березовые связи" с Европой демонстрируют представители семейства Cortinariaceae (Cortinarius tofaceus, Hebeloma sacchariolens, Inocybe splendens).
Интразональные ольшаники в поймах рек могли также служить проводником ряда европейских видов. Новый для России вид Cantharellus borealis обнаружен под ольхой, как и новый для Амурской области Naucoria striatula, отмечаемый в ольшаниках и хвойных лесах с примесью ольхи.
Если рассмотреть группу голарктических видов (162), то и в данном случае микобиота базидиальных макромицетов юга Амуро-Зейского междуречья стоит особняком при сравнении с рядом восточных микобиот заповедных территорий, использованных в наших анализах. Например, в отличие от последних, изученная микобиота содержит очень незначительное количество видов (Boletinus spectabilis, Boletus inedulis, Leccinum chromapes, Ramariopsis laeticolor) с характерным распространением, известным как "дизъюнкция Аза Грея" и означающим, что в восточной Северной Америке и восточной Азии встречаются одни и те же виды или замещающие друг друга в одном из этих регионов.
Столь же немногочисленна группа видов (Cryptoporus volvatus, Marasmius plicatulus, Mycena odorifera, Pholiota polychroa, Tremella mesenterella), ареал которых можно назвать "амфипацифическим", так как они распространены в Азии и Северной Америке по обе стороны Тихого океана - с различной глубиной проникновения на материки. Это почти полное отсутствие в группе голарктических видов амфипацифических и восточно-американских связей, столь характерных для более восточных микобиот (Wu, Mueller, 1997; Mueller et al., 2001), как и в случае с палеарктическим компонентом, говорит о возможности особых путей исторического формирования комплекса базидиальных макромицетов на юге Амуро-Зейского междуречья.
Рассматривая отличия дальневосточной и европейской флор, т. е. флор Тихоокеанской и Атлантической сторон Евразии, М.Г. Попов (1983) указывал, что флоры обоих побережий представляют разошедшиеся из единой аркто-третичной флоры разновозрастные "братские флоры". Лесную растительность, в которой доминантами служат бук, граб и дуб, он называл пребореальной и полагал, что сплошной полосы пребореального леса, протянувшейся через всю Евразию, не было даже в плиоцене, так что очень давно европейский "буково-грабово-дубовый округ" оторван от "грабово-дубового" дальневосточного. Тем не менее, многие микоризообразователи, связанные с буком и дубом в Европе, но умеющие формировать микоризу с хвойными породами, могли распространиться по сосновому "экологическому мосту" до Амура, а затем снова перейти в "дубовый комплекс" местной растительности. Еще более это справедливо для ксилотрофных базидиомицетов, которые как будто "стекаются" с разнообразных лиственных пород Европы на немногие доминанты изученной микобиоты.
Таким образом, микобиота юга Амуро-Зейского междуречья является, скорее, "атлантической" по происхождению, а не "тихоокеанской", или, скорее, "западно-атлантической", так как с восточной Северной Америкой связи такие же слабые, как и с побережьями Тихого океана.
Глава 8. Практическое значение
базидиальных макромицетов
В главе приводятся сведения о съедобных и ядовитых видах, описываются лечебные свойства 93 видов грибов и способы их применения в народной медицине. Здесь также помещен список редких и рекомендуемых к охране видов грибов района исследования; предложены меры по их сохранению.
ВЫВОДЫ
1. Биота базидиальных макромицетов юга Амуро-Зейского междуречья к настоящему времени насчитывает 392 вида, относящихся к 149 родам, 59 семействам и 21 порядку. Новыми для территории Амурской области являются 108 видов, причем 9 видов - новые для территории Российского Дальнего Востока, а 5 видов - новые для территории России.
2. В изученной биоте базидиомицетов преобладают представители агарикоидных и афиллофороидных групп порядков, а также семейств Tricholomataceae, Coriolaceae, Cortinariaceae, Russullacea и родов Russula, Inocybe, Lactarius, Cortinarius, что указывает на ее голарктический характер и характеризует в целом как неморально-бореальную.
3. Сопоставление таксономической структуры и видового состава изучаемой биоты базидиомицетов с таковыми заповедников Дальнего Востока России выявляет заметное преобладание различия над сходством. Микобиота юга Амуро-Зейского междуречья занимает 5-е место по количеству видов после таких заповедников, как Уссурийский, Кедровая падь, Лазовский и Большехехцирский, которые относятся к той же манчжурской флоре. Таксономическое своеобразие исследованной микобиоты состоит в том, что более трети видового состава (37 %) базидиальных макромицетов - виды сравнительно редкие на российском Дальнем Востоке или отсутствующие в других его районах.
4. В биоте базидиальных грибов Амуро-Зейского междуречья выделено 12 трофических групп, из которых лучше всего представлены группы микоризообразователей и сапротрофов на древесине, что в целом соответствует распределению трофических групп в лесных сообществах. Наибольшим разнообразием трофических групп отличаются семейства Tricholomataceae, Strophariaceae и Coprinaceae.
5. В изученной микобиоте большинство видов базидиомицетов консортивно связано с основными лесообразующими породами территории Амуро-Зейского междуречья - дубом, березой, сосной и лиственницей. Экологическое же своеобразие биоты базидиальных макромицетов состоит в том, что многие эвритрофные виды приурочены к одной из пород - эдификаторов лесной растительности исследованной территории и обнаруживают региональную стенотрофность. В связи с этим на каждой из доминантных пород выявлено значительно больше макромицетов, чем в одном из самых богатых по видовому составу грибов заповедников России - Уссурийском.
6. Наибольшим видовым разнообразием отличаются березово-дубово-сосновые, березово-дубовые и дубово-сосновые леса. В лесных ценозах преобладают группы микоризообразователей и сапротрофов на древесине; для лугов и рудеральных местообитаний характерно превалирование групы гумусовых сапротрофов.
7. Ядро биоты базидиомицетов района исследования образуют виды с обширными ареалами, В целом, биота базидиальных макромицетов юга Амуро-Зейского междуречья характеризуется как голарктическая бореонеморальная с выраженными в большей степени неморальными чертами, но по своим географическим связям является "западно-атлантической", а не "тихоокеанской".
8. На территории района исследования хозяйственно ценными являются около 200 видов грибов (съедобные - 136, а условно съедобные - 14). Перспективными источниками лекарственного сырья можно считать 93 вида базидиальных грибов.
9. В биоте базидиальных макромицетов территории Амуро-Зейского междуречья зарегистрировано 6 видов грибов, занесенных в Красную книгу России. Из общего числа выявленных видов базидиомицетов нами к охране рекомендуется 31 вид. Для распространения грибов на территории района исследования основными лимитирующими факторами являются: пожары; рекреационная нагрузка на лесные экосистемы и хозяйственная деятельность человека. Из основных мер охраны грибов предлагается: сохранение естественных местообитаний грибов; создание Красной книги Амурской области; создание сети заказников и микрозаказников.
Список работ, опубликованных по теме диссертации
Статьи, опубликованные в ведущих рецензируемых научных журналах:
1. Булах Е.М., Говорова О.К., Таранина Н.А. Базидиальные макромицеты Зейского заповедника // Микология и фитопатология. 2003. T. 37. Вып. 2. С. 1-7.
2. Таранина Н.А. Агарикоидные базидиомицеты лесного урочища «Мухинка» (Амурская область) // Микология и фитопатология. 2005. Т. 39. Вып 5. С. 55-63.
Статьи, опубликованные в отечественных журналах и
периодических изданиях:
3. Говорова О.К., Таранина Н.А. Биота афиллофоровых и гетеробазидиальных грибов Амурской области // Проблемы экологии Верхнего Приамурья: сборник научных трудов. Благовещенск: БГПУ, 2002. Вып. 6. С. 75-92.
4. Таранина Н.А. История изучения агарикоидных базидиомицетов Амурской области // Проблемы экологии Верхнего Приамурья: сборник научных трудов. Благовещенск: БГПУ, 2003. Вып. 7. С. 139-146.
5. Павлова Л.М., Таранина Н.А., Куимова Н.Г., Безбородов В.Г. Инвентаризация зеленого фонда г. Благовещенска // Вестник АмГУ. 2005. Вып. 30. Серия естеств.-биол. науки. С. 57-59.
6. Таранина Н.А. Микоризные грибы юга Амуро-Зейского междуречья // Проблемы экологии Верхнего Приамурья: сборник научных трудов. Благовещенск: БГПУ, 2006. Вып. 9. С. 129-139.
Работы, опубликованные в материалах региональных,
всероссийских, международных конференций:
7. Таранина Н.А. Проблема охраны афиллофоровых и агарикоидных грибов в Амурской области //Амурская наука на пороге III тысячелетия. Труды региональной научной конференции студентов, аспирантов и молодых ученых, посвященной 275-летию РАН. Благовещенск: АмурКНИИ АмурНЦ ДВО РАН, 2000. С. 44-45.
8. Таранина Н.А. Афиллофоровые базидиомицеты Зейского заповедника // Будущее Амурской науки. Труды региональной научной конф. студентов, аспирантов и молодых ученых. Благовещенск: АмурКНИИ АмурНЦ ДВО РАН, 2001. С. 356-360.
9. Таранина Н.А. Изучение дереворазрушающих базидиомицетов на территории Ботанического сада АмурНЦ ДВО РАН // Молодежь XXI века: шаг в будущее. Сборник трудов III регион. науч.-практ. конф. Благовещенск: Амурский гос. ун-т, 2002. С. 114.
10. Таранина Н.А. Базидиальные грибы г. Благовещенска // Молодежь XXI века: шаг в будущее: Матер. IV регион. науч.-практ. конф. 14-15 мая 2003 г. Благовещенск. 2003. С. 186-187.
11. Таранина Н.А. Гастеромицеты Амурской области // Сборник докладов VII Дальневосточной молодежной школы-конф. по актуальным проблемам химии и биологии, МЭС ТИБОХ, Владивосток, 22-25 сентября 2003 г.: Тезисы докладов. Владивосток: ДВО РАН, 2003. С. 61-62.
12. Таранина Н.А. Съедобные и ядовитые грибы Амурской области // Материалы научн.-практ. конф. «Приамурье от первопроходцев до наших дней», Благовещенск (23-24 октября 2003 г.). Благовещенск: Амурский областной краеведческий музей им. Новикова-Даурского, 2003. С. 40-43.
13. Таранина Н.А. Базидиомицеты юга Амуро-Зейского междуречья // Ботанические исследования в азиатской России: Матер. XI съезда Русского ботанического общества (21-22 августа, 2003 г., Новосибирск-Барнаул). Т. 1. Барнаул: “Азбука”, 2003. С. 66-67.
14. Таранина Н.А. Паразитические базидиомицеты Амурской области // Материалы VIII Молодежной конференции ботаников в Санкт-Петербурге (17-21 мая 2004 г.). СПб., СПГУТД, 2004. С. 74.
15. Таранина Н.А. Трофические группы базидиальных грибов Амуро-Зейского междуречья // Материалы VIII Молодежной конференции ботаников в Санкт-Петербурге (17-21 мая 2004 г.). СПб.: СПГУТД, 2004. С. 75.
16. Таранина Н.А. Мониторинг биоты базидиальных грибов урбанизированных территорий на примере г. Благовещенска (Амурская область) // Тезисы докладов ХI Дальневосточной молодежной школы-конф. по актуальным проблемам химии и биологии (16-23 сентября, МЭС ТИБОХ, Владивосток, 2005 г.). Владивосток: ДВО РАН, 2005 г. С. 56.
17. Павлова Л.М., Таранина Н.А., Куимова Н.Г., Безбородов В.Г. Состояние зеленых насаждений г. Благовещенска // Материалы IX конференции «Проблемы озеленения крупных городов», 6-9 декабря 2005 г. М.: «Прима-М», 2005. Вып. 11. С. 63-66.
18. Taranina N.A. Tremelloid Basidiomycetes of southern part of interfluve of the Amur-river and the Zeya-river // Материалы 1 (X) междунар. конф. ботаников в Санкт-Петербурге (21-26 мая 2006 г.). СПб., 2006. С. 340.
Кочунова Наталья Анатольевна
БАЗИДИАЛЬНЫЕ МАКРОМИЦЕТЫ
ЮГА АМУРО-ЗЕЙСКОГО МЕЖДУРЕЧЬЯ
Автореферат
Лицензия ЛР 020427 от 25.04.1997 г.
Подписано в печать 31.08.2007. Формат 60х84 1/16.
Бумага офсетная. Печать офсетная. Печ. л. 1,0.
Тираж 120 экз. Заказ 256.
______________________________________________________________
Отпечатано в отделе оперативной полиграфии издательства ДальГАУ
675000, г. Благовещенск, ул. Политехническая, 86
[1] Здесь рассматриваются только представители эколого-трофических групп, обладающие тесными консортивными связями с древесными растениями: микоризообразователи, ксилотрофы на мертвой и живой древесине, сапротрофы на опаде.
[2] Для удобства восприятия номенклатуры долготная составляющая в названиях типов ареала приводится первой.