

Эколого–ценотические и биохимические особенности солодки голой (glycyrrhiza glabra l.) в калмыкии
На правах рукописи

Алексеева Таисия Баатровна
ЭКОЛОГО–ЦЕНОТИЧЕСКИЕ И БИОХИМИЧЕСКИЕ ОСОБЕННОСТИ СОЛОДКИ ГОЛОЙ (GLYCYRRHIZA GLABRA L.) В КАЛМЫКИИ
03.00.16 – экология
Автореферат
диссертации на соискание ученой степени
кандидата биологических наук
Саратов – 2007
Работа выполнена в государственном образовательном учреждении
высшего профессионального образования
«Калмыцкий государственный университет»
на кафедре ботаники и зоологии
Научный руководитель: доктор биологических наук, профессор
Бакташева Надежда Мацаковна
Официальные оппоненты: доктор биологических наук, профессор
Березуцкий Михаил Александрович
кандидат биологических наук, доцент
Смекалова Тамара Николаевна
Ведущая организация: ГОУ ВПО «Астраханский государственный
университет»
Защита состоится «_8_» ноября 2007 г. в 12.00 часов на заседании диссертационного совета Д 212.243.13 при государственном образовательном учреждении высшего профессионального образования «Саратовский государственный университет им. Н. Г. Чернышевского» по адресу: 410012, г. Саратов, ул. Астраханская, д. 83. E-mail: [email protected].
С диссертацией можно ознакомиться в Зональной научной библиотеке ГОУ ВПО «Саратовский государственный университет им. Н. Г. Чернышевского».
Автореферат разослан «_6_» октября 2007 г.

Ученый секретарь
диссертационного совета С. А. Невский
Общая характеристика работы
Актуальность исследования. Корни и корневища солодки голой (Glycyrrhiza glabra L.) – лакричный корень – издавна применяются в медицине и являются источником целого ряда лечебных препаратов, применяемых человеком при самых различных заболеваниях (Оболенцева, 1966, 1999; Быков, 1996; Степанова, 1997). Кроме медицины, солодковый корень находит разнообразное применение более чем в 20 отраслях промышленности: табачной, пищевой, в цветной металлургии и т.д. (Михайлова, 1976; Шитов, 1980). Надземная масса солодки используется в качестве корма для животных. В сене и силосе она хорошо поедается всеми видами животных, но на пастбище – только в отаве (Ларин, 1951).
Основные запасы сырья G. glabra находятся в республиках Средней Азии и Казахстане. В современной России обозначилась необходимость изучения состояния популяций солодки голой уже в пределах новых границ – Астраханской, Волгоградской и Ростовской областях, Республике Калмыкия (РК) и Краснодарском крае (Швыдкая, 2001). В РК солодка встречается спорадически и не образует самостоятельных зарослей. Ее запасы до конца не исследованы, а также не проводилось изучение химического состава, содержания биологически активных веществ и других компонентов, что позволило бы определить возможности рационального использования солодкового корня и введения G. glabra в культуру.
Цель и задачи исследования. Основная цель работы – выявление эколого-ценотических и биохимических особенностей солодки голой, а также вопросов ее охраны в РК.
Для достижения этой цели решались следующие задачи:
- выявить эколого-ценотические и анатомические особенности солодки голой в РК;
- определить содержание в солодке голой биологически активных веществ (БАВ) и минеральных элементов в зависимости от различных экологических факторов;
- изучить динамику накопления БАВ в подземных органах солодки голой для установления оптимальных сроков ее заготовки в условиях РК;
- оценить химический состав надземной массы солодки голой для предполагаемой рекомендации в качестве корма для сельскохозяйственных животных;
- изучить запасы солодки голой для определения статуса вида и разработки возможных мер по ее охране.
Научная новизна. Впервые в условиях РК наиболее полно исследовались эколого-ценотические и биохимические особенности G. glabra. Изучена динамика накопления БАВ и выявлена степень влияния различных экологических факторов на их накопление. Определен микроэлементный состав надземной и подземной частей солодки, а также почв с мест ее произрастания. Рассмотрен химический состав надземной массы для возможной рекомендации в качестве корма сельскохозяйственным животным. Изучены запасы солодки голой и определен статус вида; внесены предложения по ее охране.
Практическое значение. Результаты изучения динамики накопления биологически активных веществ в солодке по фазам вегетации дают возможность установить оптимальные сроки для ее возможной заготовки в условиях РК. На основе установленных эколого-ценотических и биохимических особенностей солодки голой определены оптимальные условия для введения этого вида в культуру. Полученные данные о химическом составе надземной фитомассы позволят использовать этот вид в качестве корма для сельскохозяйственных животных.
Апробация работы. Результаты исследований доложены на XI Съезде Русского Ботанического общества (Барнаул, 2003); VIII Молодежной конференции ботаников (Санкт-Петербург, 2004); I (IX) Международной конференции молодых ботаников (Санкт-Петербург, 2006); Международной научно-практической конференции, посвященной 100-летию со дня рождения профессора В.В. Иванова (Уральск, 2005); IV Международном симпозиуме «Степи Северной Евразии» (Оренбург, 2006); Международной научно-практической конференции, посвященной 10-летию Государственного природного заповедника «Ростовский» (Ростов-на-Дону, 2006); Международной научно-практической конференции, посвященной 75-летию Астраханского государственного университета (Астрахань, 2007).
Публикации. По материалам диссертации опубликовано 10 печатных работ, в том числе одна в издании перечня ВАК РФ.
Объем и структура работы. Диссертация изложена на 174 страницах текста, состоит из введения, пяти глав и выводов, содержит 33 таблицы и 21 рисунок. Список использованной литературы включает 183 наименования отечественных и зарубежных авторов. Приложение состоит из списка 262 видов сосудистых растений, 14 фотографий и 10 таблиц матриц коэффициентов сопряженности.
Основные положения, выносимые на защиту
- Начало вегетации солодки голой в значительной степени зависит от возрастной структуры ее популяций и температурного фактора.
- Оптимальными сроками заготовки солодкового корня являются летний и осенний периоды.
Содержание работы
Во введении обосновываются актуальность проведенных исследований, формулируются цель и задачи исследования, его новизна, теоретическое и практическое значение.
- ФИЗИКО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА
РЕСПУБЛИКИ КАЛМЫКИЯ
РК расположена на юго-востоке европейской части Российской Федерации между 44040/ и 47035/ северной широты и 40010/ и 44050/ восточной долготы. Общая площадь республики – 75,9 тыс. км2 (Агроклиматические ресурсы КАССР, 1974).
Рельеф республики равнинный с предельными колебаниями высот от 28 до 222 м (Агроклиматический справочник, 1967). Климат резко континентальный – лето жаркое и очень сухое, зима малоснежная. Континентальность возрастает с запада на восток. Среднегодовое количество осадков невелико – от 210 до 420 мм. Особенностью климата является преобладание ветров восточных румбов. Сильные ветры со скоростями 15-20 м/сек и более отмечаются не только в течение вегетационного периода, но и в зимнее время. Число таких дней колеблется от 15 до 30.
Почвенный покров РК отличается комплексностью (Бакинова, Зеленская, 1998). На долю черноземов, каштановых, светло-каштановых, бурых полупустынных почв, солонцов автоморфных каштановых и полупустынных, в РК приходится 71,6% общей площади.
Гидрографическая сеть развита в западной части республики и представлена балками склона Ергеней, Сарпинского и Даванского понижений, долины рек Маныча и Кумы. Главным источником питания рек являются талые снеговые воды. Грунтовое питание незначительно, убывает с севера на юг и зависит от размеров водосбора (Емгушев, 1972). На юго-востоке территория РК выходит к Каспийскому морю, на востоке на небольшом расстоянии – к Волге.
Территория РК расположена на стыке двух зон растительности: степной и пустынной. Степные сообщества образуют комплексы с пустынными (Димо, Келлер, 1907). В этих комплексах степные и пустынные компоненты встречаются рядом, образуя пятна, приуроченные к разным элементам микрорельефа. Зональные же изменения растительности в направлении с запада на восток выражаются в соотношении между компонентами комплексов.
2. МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Исследования проводили в 2001–2006 годах. Объектом исследования послужила G. glabra L., произрастающая в естественных местообитаниях. За единицу исследования нами была взята ценотическая популяция (ЦП) (Миркин, 1989). Маршрутные и стационарные обследования проведены в 36 ЦП на территории Юстинского (ЦП1.1 – ЦП1.5), Малодербетовского (ЦП2.1 – ЦП2.6), Сарпинского (ЦП3.1 – ЦП3.8), Кетченеровского (ЦП4.1 – ЦП4.5), Целинного (ЦП5.1 – ЦП5.8), Приютненского (ЦП6.1 – ЦП6.3), Ики-Бурульского (ЦП7.1) районов РК в составе разнотравно-злаковых ценозов (рис. 1).
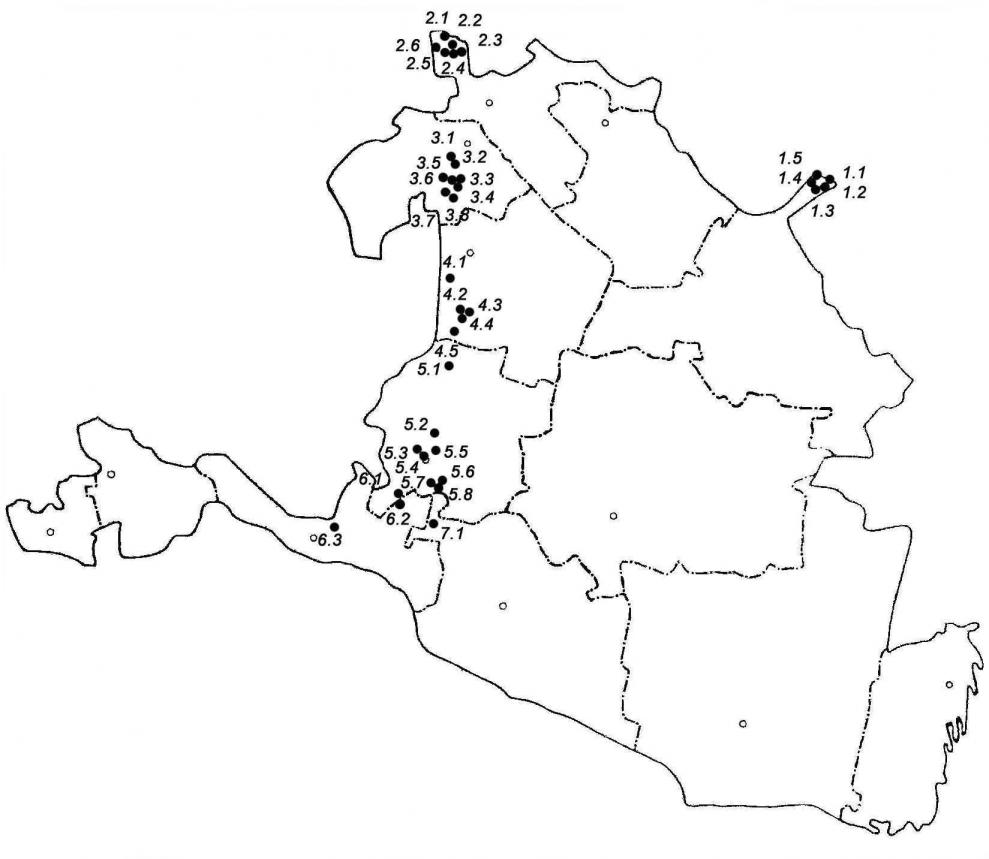
Рис. 1 Карта-схема мест исследования
Для идентификации собранного материала были просмотрены гербарные коллекции Ботанического института им. В.Л. Комарова РАН. Оценка численности видов, а также доля участия солодки в травостое, определялась в соответствии со шкалой, предложенной О. Друде (Воронов, 1973). Видовые названия устанавливались по определителям флоры Северо-Западного Кавказа и Предкавказья (Косенко, 1970), флоры Европейской части СССР (Станков, 1949) и флоры средней полосы европейской части России (Маевский, 2006). Латинские названия растений и их номенклатура приведены по сводке С.К. Черепанова (1995).
Анатомические исследования проводили по методикам, предложенным И.А. Наумовым, В.И. Козловым (1954) и Г.Г. Фурстом (1979). Исследования проводились на микроскопе МБИ-3. Рисунки с препаратов изготавливались с микроскопа с помощью рисовального аппарата РА-1.
Фенологические наблюдения вели по методике И.Н. Бейдеман (1974). Деление растений на жизненные формы осуществляли по классификации И.Г. Серебрякова (1962). При экологическом анализе использовалась классификационная система, основанная на принципах А.П. Шенникова (1950), но с выделением комбинированных подгрупп растений.
Урожайность (плотность запасов сырья) солодки определялась методом модельных экземпляров (Буданцев, Харитонова, 1999; Фармакогнозия, 2006). Урожайность установлена для 10 наиболее крупных ЦП.
Наличие биологически активных веществ (сахаров и флавоноидов), а также макроэлементов (азота, фосфора, калия (NPK)) определяли по: Методам биохимического исследования растений (1987); Н.И. Гринкевич, Л.Н. Сафронич (1983). Количественное определение содержания глицирризиновой кислоты проводилось титриметрически (Муравьев, 1964; Государственная Фармакопея, 1968; Толстиков, 1969). Содержание кальция, магния, железа, марганца, меди, цинка в надземной и подземной массе солодки определяли методом атомно-абсорбционной спектроскопии на ААS-1 (Карл Цейс, Германия) (Ринькис, 1987). Кормовые достоинства травы солодки (протеин, клетчатка), а также определение гигровлаги и зольности проводили по П.Т. Лебедеву, А.Т. Усовичу (1969). Для агрохимической характеристики почв использовались следующие виды анализа: потенциометрическое определение рН водной вытяжки (Аринушкина, 1970), NPK и анионный (HCO3-, Cl-, SO42-) состав водных вытяжек, содержание кальция, магния, железа, марганца, меди, цинка в надземной массе солодки также проводили методом атомно-абсорбционной спектроскопии (Методы определения микроэлементов в почвах, растениях и водах (1974)).
На учетных площадках, закладываемых при проведении геоботанических исследований, учитывалось наличие или отсутствие видов; затем эти данные обрабатывались с помощью таблиц 2x2 (Василевич, 1972). В анализ сопряженности были включены виды, имеющие встречаемость от 10% и выше. Вычисление коэффициентов межвидовой сопряженности проводилось по формуле Бравэ-Пирсона (КВ) (Нешатаев, 1987). Для сравнения видового состава солодковых ценопопуляций использовали коэффициент общности Серенсена-Чекановского (КSC).
Состояние особи вида изучалось методом установления эколого-ценотического градиента (экоклина) с использованием показателей (жизненности) растений в ценопопуляциях по размерному спектру особей (Ишбирдин, Ишмуратова, 2004).
Статистическая обработка данных и построение диаграмм проводились в программе Excel.
3. ЭКОЛОГО-ЦЕНОТИЧЕСКИЕ И АНАТОМИЧЕСКИЕ ОСОБЕННОСТИ СОЛОДКИ ГОЛОЙ В РЕСПУБЛИКЕ КАЛМЫКИЯ
В РК солодка распространена спорадически, в основном в поймах рек, по балкам Ергенинской возвышенности. Здесь она является мезоксерофитом и мезогалофитом. Ее популяции приурочены к подзоне настоящей (разнотравно – дерновиннозлаковой) степи и нередко связаны с луговыми почвами средней степени засоления.
Исследования показали, что у большинства растений РК начало вегетации сильно зависит от температурного фактора (рис. 2).
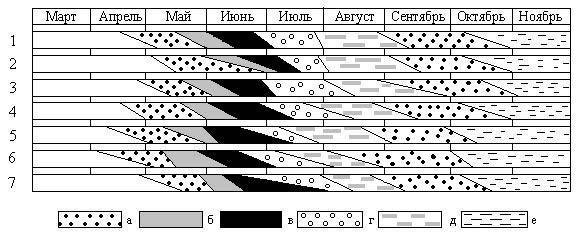
Рис. 2 Фенологические спектры солодки голой
а – вегетация, б – бутонизация, в – цветение, г – плодоношение, д – созревание плодов, е – отмирание, 1 – ЦП5.6 (2003 г.), 2 – ЦП5.7 (2004 г.), 3 – ЦП6.3 (2004 г.), 4 – ЦП6.3 (2005 г.), 5 – ЦП3.5 (2005 г.), 6 – ЦП2.4 (2004 г.), 7 – ЦП1.2 (2005 г.)
В целом по республике, в районах, неоднородных по климатическому режиму, можно проследить следующий ряд начала вегетации и цветения: Малодербетовский и Сарпинский Целинный и Приютненский Кетченеровский и Ики-Бурульский Юстинский. Начало вегетации зависит от возрастного состояния самих растений. Так, в Малодербетовском районе (самом северном) в ценопопуляциях, где основную часть составляют генеративные растения, наблюдаются более ранние сроки наступления бутонизации и цветения. Повышенная влажность и медленное нарастание температуры в Юстинском районе, в пойме р. Волги, существенно сдвигает наступление фенофаз у солодки.
В исследуемых ценопопуляциях собрано и определено 262 вида дикорастущих сосудистых растений, относящихся к 48 семействам и 168 родам. Анализ систематической структуры ведущих семейств показывает, что наиболее представительными являются шесть семейств: Asteraceae (15.99% от общего числа родов), Poaceae (11.24%), Brassicaceae (6.51%), Fabaceae (5.92%), Lamiaceae (5.33%), Apiaceae (4.74%). В целом, соотношение соотношение видов и родов, а также спектр ведущих семейств, входящих в состав ценопопуляций солодки голой, соответствует таковым флоры РК (Бакташева, 2000).
Показательным для характеристики флористического состава солодковых ценопопуляций является количественный состав родов. Крупных родов пять, они включают более шести видов: Astragalus (10 видов), Potentilla (9), Artemisia (7), Carex (7), Achillea (6). На долю этих родов приходится 39 видов, что составляет 14,89% от всего видового состава.
В сложении солодковых ценопопуляций большую роль играют многолетние травянистые растения, которые представлены 165 видами (рис. 3). Их процентное соотношение (62.98%) сохраняется и варьирует в небольших пределах.
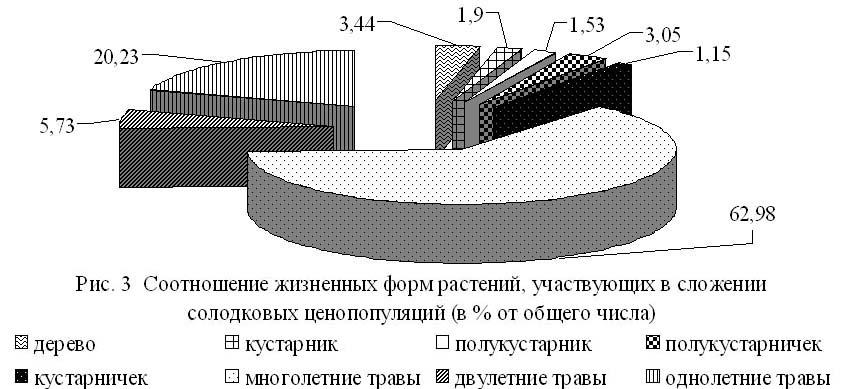
Основными лимитирующими факторами для солодки голой, как и для большинства растений республики, являются дефицит влаги и засоленность. Проведенный экологический анализ показал, что ведущая роль в сложении солодковых ценопопуляций принадлежит 4 экологическим группам растений: эвксерофитам, эвриксерофитам, мезоксерофитам и эвмезофитам (рис. 4). Необходимо отметить, что их процентное отношение в изученных ценопопуляциях хотя и меняется под влиянием неодинаковых климатических и эдафических условий природных районов республики, но изменения незначительны.
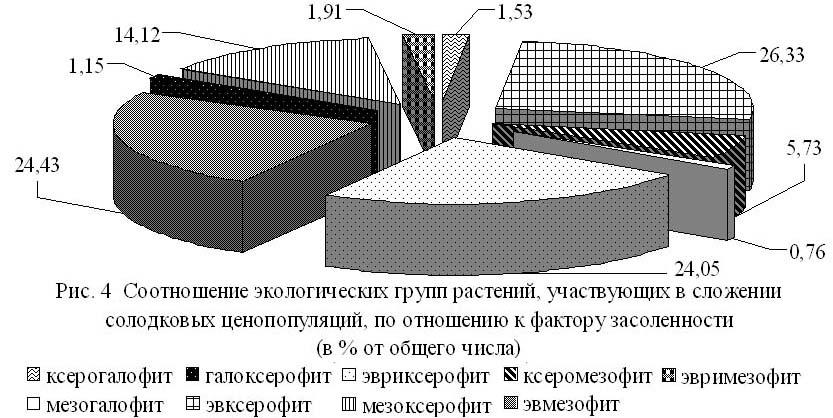
Отмечено, что солодка, произрастающая по протяжинам балок Ергеней и в поймах рек, в условиях среднего увлажнения, активно осваивает целинные равнинные территории. Сначала это единичные экземпляры, затем небольшие пятна, которые постепенно увеличиваются и все больше теряют связь с «балочными» растениями. Крупные ценопопуляции солодки отмечаются и в подзоне сухой степи, где в сложении ценозов основная роль приходится на эвксерофиты. Это можно объяснить тем, что здесь солодка не испытывает такого сильного прессинга со стороны плотнодерновинных злаков, стержнекорневых, корнеотпрысковых и корневищных многолетников. Незначительная доля, от всего состава растений, участвующих в сложении ценопопуляций, приходится на ксеро–, мезогалофиты и галоксерофиты, что свидетельствует о слабой засоленности исследуемых территорий.
Применение коэффициента общности Серенсена-Чекановского (KSC) позволило выявить 30 видов растений общих для всех 36 солодковых ценопопуляций. Тридцать видов растений объединены в 2 группы; коэффициенты общности находятся в пределах от 32 до 76%. Ниже приведен фрагмент дендрограммы, отражающей сходство флористического состава солодковых ценопопуляций (рис. 5). Самый высокий коэффициент связи (76%) отмечен для солодки голой и пырея ползучего.
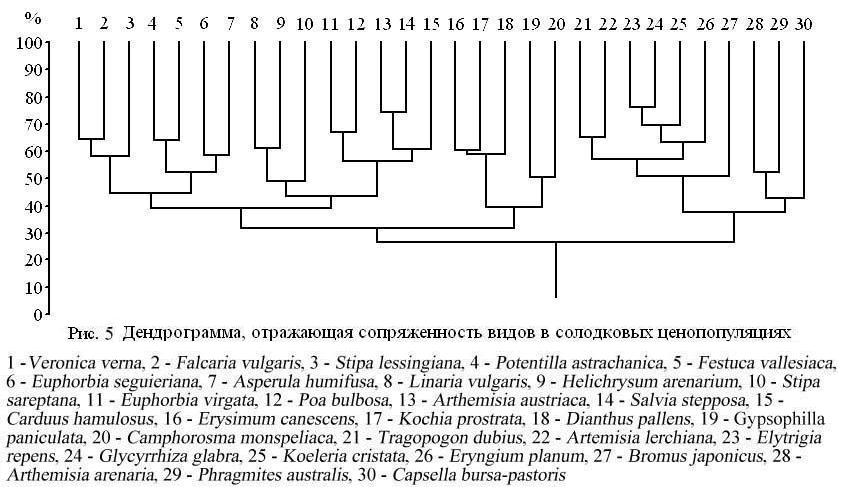
Они составляют ядро первой группы, в состав которой входят по убывающей: K. cristata, E. planum, T. dubius, A. lerchiana, B. japonicus, A. arenaria, Ph. australis, C. bursa-pastoris. Вторая группа состоит из 3 подгрупп, связанных друг с другом более тесно, чем с предыдущей группой. Она объединяет 20 видов растений, которые хотя и встречаются во многих растительных группировках, но все же предпочитают солодковые ценопопуляции.
Полученные результаты были подтверждены и уточнены с помощью метода межвидовых сопряженностей, с применением коэффициента Бравэ-Пирсона. Корреляционная матрица, составленная для перечисленных выше 30 видов, показала наличие 151 отрицательного коэффициента сопряженности и 242 положительных. Отношение числа достоверно отрицательных сопряженностей к числу достоверно положительных связей матрицы равно 0.62 и указывает на высокую склонность видов образовывать сообщество с участием солодки.
Объектом анатомического исследования послужили растения, произрастающие на территории Целинного района (ЦП5.7 и ЦП5.8) и отличающиеся морфологически. Солодка из первой ценопопуляции характеризовалась типичными «голыми» бобами (далее упоминается как форма typica), из второй – большим количеством шипиков на перикарпии плодов, «шиповато-железистыми» бобами (далее упоминается как форма glandulifera). Сравнению были подвергнуты анатомическое строение стебля, листа корневища, корня.
Стебель G. glabra покрыт однослойным эпидермисом. Клетки эпидермиса незначительно вытянуты и имеют прямые стенки. На эпидермисе у солодки голой есть простые толстостенные волоски с многоклеточным основанием, железистые чешуйки, которые состоят из короткой ножки, переходящей в многоклеточную полушаровидной формы головку, и шипики (рис. 6). Стебли typica покрыты большим количеством шипов. Внутреннее строение стебля обычно для двудольных растений (Шамсувалиева, 1999). Особенностью строения солодки является наличие во всех частях стебля клеток идиобластов, содержащих слизи и пигменты. Колленхима пластинчатого типа; у glandulifera развита лучше. Внутрь от колленхимы располагаются овальные и округлые паренхимные клетки первичной коры, содержащие хлоропласты. Среди паренхимных клеток в большом количестве имеются более крупные идиобласты, заполненные, как и клетки колленхимы, слизями. У typica идиобласты крупные. Во многих паренхимных клетках обнаружен ассимиляционный крахмал. Проводящая система пучкового типа. Флоэма состоит из ситовидных трубок небольшого диаметра. Камбиальное кольцо выражено слабо и состоит из 1-2 слоев клеток прозенхимного типа с заостренными окончаниями. Древесина состоит из первичной и вторичной ксилемы. Вторичная ксилема развита сильнее. У glandulifera ксилема существенно толще, чем у typica. Диаметр сосудов у typica солодки очень мал. Сердцевинные лучи многочисленны. Тяжи древесной паренхимы состоят из 2-5 клеток. Перимедулярная зона состоит из клеток паренхимного типа. К перимедулярной зоне примыкают идиобласты, более удлиненные клетки, иногда с заостренными концами. Центральную часть стебля занимает сердцевина, клетки которой имеют округло-многоугольную форму (рис. 7). В этой части у солодки голой находится много идиобластов. У glandulifera число клеток идиобластов в центре сердцевины незначительно. Они располагаются в основном в непосредственной близости от первичной ксилемы проводящих пучков.
Лист. Листочки сложного листа солодок имеют сходную структуру (рис. 8). Однако толщина листовой пластинки и кутикулы у typica была больше, чем у glandulifera. На нижнем эпидермисе имеются железистые чешуйки, устьица и простые волоски. Клетки палисадной и губчатой паренхимы очень сходны и трудно различимы. Проводящие пучки листьев закрытые коллатеральные.
Корневище солодки голой длинное, в поперечнике округлое, покрытое шелушащейся коричневой перидермой. Перидерму образуют толстостенные прямоугольной формы клетки пробки, лежащие строго радиальными рядами (рис. 9). Пробка состоит из 5-10 слоев клеток. Корневище glandulifera имеет хорошо развитую пробку. К пробке примыкают крупные клетки феллодермы; они имеют округлую форму. В ее клетках имеются жиры и крахмальные зерна. Механическая ткань хорошо развита. Тяжи лубяных волокон имеют кристаллоносную обкладку. Во вторичной коре корневища хорошо выражены сердцевинные лучи, пересекающие кольцо флоэмы в радиальном направлении. Древесина корневища рассеянно-сосудистого типа, сосуды образуют радиальные ряды, чередующиеся с довольно широкими сердцевинными лучами.
Сосуды членистые бочонковидные, между ними расположены небольшие группы волокон либриформа. Древесная паренхима вазицентрическая. Тяжи древесной паренхимы состоят из 2-5 клеток. Сердцевинные лучи корневища представлены 1-6 рядами. Их много у glandulifera.
| Рис. 6 Поперечный срез стебля (ув. 400) G. glabra glandulifera. 1 – железки, 2 – волоски, 3 – кутикула, 4 – клетки эпидермиса, 5 – идиобласты, 6 – первичная кора, 7 – механическая ткань, 8 – флоэма, 9 – камбий, 10 – ксилема, 11 – сердцевина Рис. 7 Сердцевина стебля. I – G. glabra typica, II – G. glabra glandulifera Рис. 8 Участок поперечного среза листа солодок (ув. 280). I – G. glabra typica, II – G. glabra glandulifera, 1 – верхний эпидермис, 2 – железистая чешуйка, 3 – столбчатый мезофилл, 4 – губчатый мезофилл, 5 – нижний эпидермис, 6 – простой волосок Рис. 9 Поперечный срез корневища G. glabra (ув. 280). 1 – пробка, 2 – механическая ткань, 3 – флоэма, 4 – жиры, 5 – крахмал, 6 – идиобласты, 7 – камбий, 8 – ксилема, 9 – сосуды, 10 – волокна либриформа, 11 – сердцевинные лучи Рис. 10 Схема радиального сечения корня (ув. 56). 1 – пробка, 2 – флоэма, 3 – механическая ткань, 4 – камбий, 5 – волокна либриформа, 6 – сердцевинные лучи, 7 – сосуды |
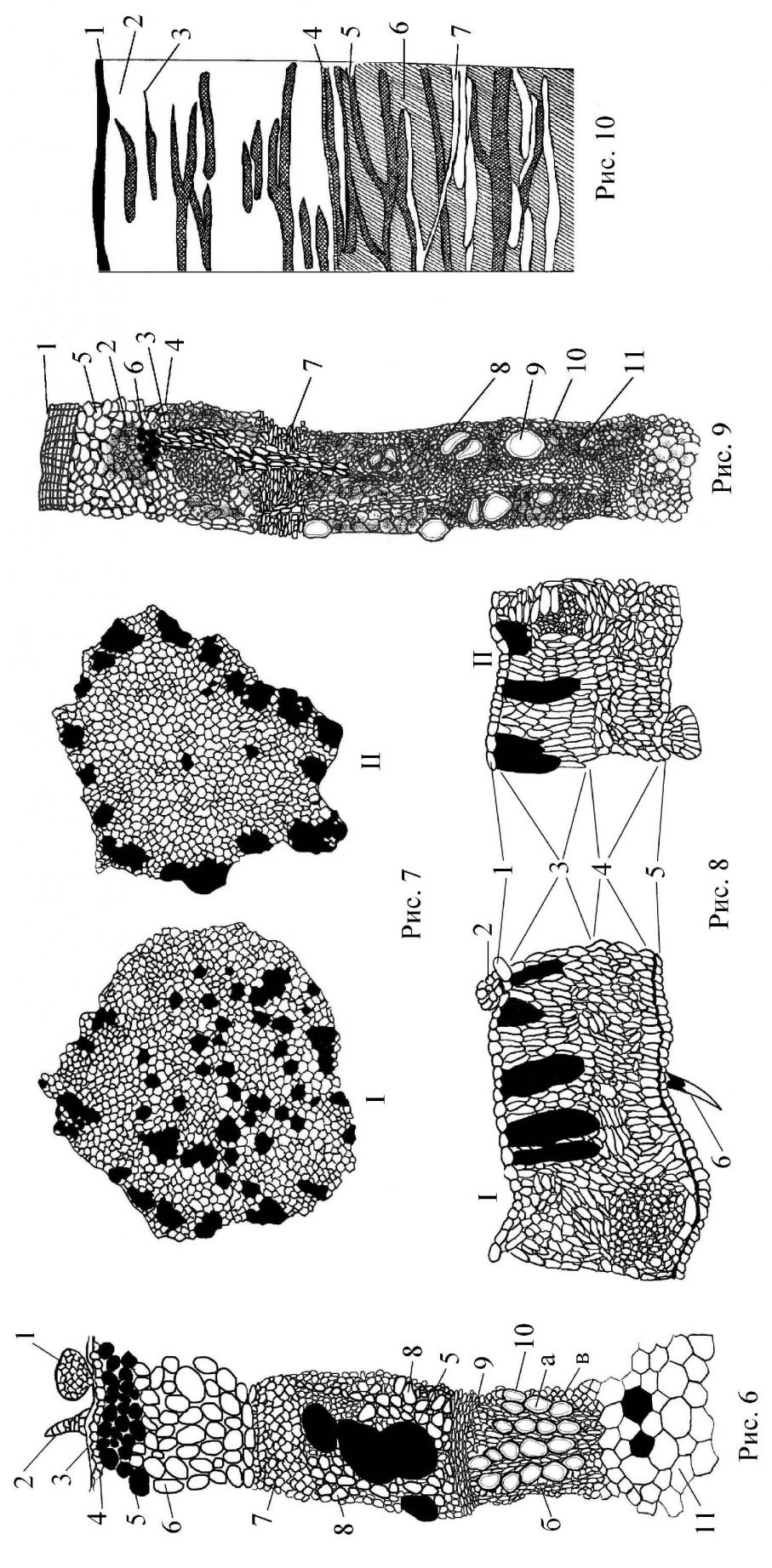
Центральную часть корневища занимает сердцевина, состоящая из клеток округло-многоугольной формы. Среди клеток сердцевины имеются вытянутые по длине корневища идиобласты. Отмечено, что typica содержит больше идиобластов, чем glandulifera. В клетках сердцевины накапливаются крахмал и жиры. Однако у последней, запасные питательные вещества содержатся в основном в клетках феллодермы и лубяной паренхимы.
Корень солодки голой с наружной стороны покрыт пробкой. Имеются чечевички. Во флоэме находятся тяжи лубяных волокон, расположенные параллельными рядами. Участки флоэмы в радиальном направлении чередуются с хорошо развитыми сердцевинными лучами (рис. 10). Ситовидные трубки тонкостенные, имеют скошенные ситовидные пластинки. Камбий состоит из 5-6 слоев клеток. Древесина представляет сплошной цилиндр. Корни солодки полиархные. Годичные кольца не выражены. Древесная паренхима хорошо развита. Стенки сосудов и волокон либриформа одревесневевшие, а древесной паренхимы и лубяных волокон клетчатковые. Сердцевинные лучи древесины заполнены крахмалом (у glandulifera). Идиобластов в корне у растений из обеих ценопопуляций обнаружено не было.
Крупные заросли G. glabra, произрастающие на территории Юстинского района, сосредоточены на небольшом участке Волго-Ахтубинской поймы. Общее проективное покрытие составляет 60-100%. В составе ценопопуляций встречаются: Bromus mollis, Anthoxanthum odoratum, Dactylis glomerata, E. planum, Inula britanica, Butomus umbellatus, Alisma plantago – aguatica и др.
Малодербетовский район характеризуется развитым сельским хозяйством, сравнительно высокой степенью распаханности земель. Проективное покрытие составляет 70-90%, в ряде мест – 100%. Доля участия солодки в травостое – 40-50%. Доминантами и субдоминантами выступают St. sareptana, A. austriaca, Achillea millefolium. В составе разнотравья отмечены лапчатка, шалфей, горошек, зопник, гвоздика и др.
Ценопопуляции солодки голой в Сарпинском районе произрастают в самых различных условиях. Общее проективное покрытие – 65-80%. Солодка соседствует с Calophaca wolgarica, Amygdalus nana, Iris pumila, Delphinium puniceum, Linaria genistifolia и др. Из злаков присутствуют: костры, типчак, пырей, мортуки и др.
Проективное покрытие ценопопуляций солодки в Кетченеровском районе – 50-60%. Встречается малочисленными популяциями. Территория испытывает сильное влияние антропогенного фактора. Травостой разреженный. В составе разнотравья встречаются: Stipa pulcherrima, Artemisia taurica, Descurainia sophia, Cachrys odontalgica, C. bursa-pastoris, Astragalus vulpinus, Helichrysum arenarium.
На территории Целинного района G. glabra встречается в основном по балкам Ергеней. Проективное покрытие составляет 70-80%. Доля участия солодки в травостое – 35-50%. В формировании травостоя в основном участвуют злаки: St. lessingiana, P. bulbosa, Puccinellia distans, E. repens, Agropyron pectiniforme, Eremopyrum triticeum.
На территории Приютненского района солодка произрастает большей частью на светлокаштановых солонцеватых почвах. Многочисленны St. lessingiana, K. cristata, K. prostrata и Carex sp.
Маршрутные исследования не выявили крупных ценопопуляций солодки на территории Ики-Бурульского района.
Значения индексов виталитета (IVC), составленные для пяти районов республики, показали, что в наилучших условиях находятся ценопопуляции солодки голой Юстинского района, а в наихудших – Кетченеровского (табл. 1). Индекс размерной пластичности (ISP) солодки голой равен 1.4, что характеризует ее как S-стратега. S-виды отличаются повышенной выживаемостью при ограниченности экологических ресурсов и имеют малую фитомассу (Grime, 1979).
Таблица 1
Индексы жизненности ценопопуляций солодки голой
и размерной пластичности вида в РК
| Признаки | Отношение сред. знач. признака в ЦП к сред. знач. признака для всех ЦП (Xi/ | Сред. знач. признака для всех ЦП ( | ||||
| Целинный | Кетченер. | Сарпинск. | Малодерб. | Юстинск. | ||
| Высота растений, см | 66.4+5.4 | 62.2+4.3 | 60.2+6.9 | 67.3+5.8 | 90.4+8.1 | 69.3 |
| Число сложн. листьев, шт. | 23.4+3.1 | 22.3+2.2 | 22.7+4.2 | 28.9+3.5 | 36.3+2.2 | 26.72 |
| Длина сложн. листа, см | 10.2+1.1 | 10.1+1.0 | 11.3+1.5 | 10.9+1.2 | 12.1+0.6 | 10.92 |
| Ширина сложн. листа, см | 5.7+0.5 | 4.7+0.8 | 6.0+0.5 | 5.5+0.2 | 6.9+0.3 | 5.76 |
| Число листочков сложн. листа, шт. | 7.9+0.5 | 7.6+1.2 | 8.9+1.5 | 10.9+1.2 | 9.7+1.3 | 9 |
| Длина листочка, см | 3.4+0.2 | 3.1+0.3 | 3.3+0.1 | 3.1+0.1 | 3.3+0.2 | 3.24 |
| Ширина листочка, см | 1.8+0.2 | 1.7+0.5 | 1.6+0.1 | 1.5+0.1 | 1.5+0.1 | 1.62 |
| Длина цветоноса, см | 10.1+1.1 | 10.1+1.5 | 10.5+0.5 | 10.6+1.8 | 10.8+0.6 | 10.42 |
| Число цветков, шт. | 20.0+2.7 | 25.5+1.3 | 34.9+1.7 | 26.5+5.0 | 35.6+1.5 | 28.5 |
| Длина цветка, см | 1.1+0.1 | 1.1+0.1 | 1.1+0.1 | 1.1+0.1 | 1.1+0.1 | 1.1 |
| Число бобов, шт. | 19.3+1.2 | 17.8+1.9 | 30.7+8.6 | 24.3+4.0 | 34.5+2.4 | 25.32 |
| Длина боба, см | 2.1+0.3 | 2.1+0.2 | 2.2+0.1 | 2.1+0.1 | 2.2+0.1 | 2.14 |
| Сумма (Xi/ | 11.2 | 10.8 | 12.3 | 12.0 | 15.3 | |
| IVC | 0.93 | 0.91 | 1.02 | 0.99 | 1.28 | |
| ISР=IVCmax/IVCmin | 1.4 | |||||
| Экоклин | Юстинский – Сарпинский – Малодербетовский – Целинный – Кетченеровский | |||||
Значения индексов виталитета, а также составленный на их основе экоклин (Юстинский – Сарпинский – Малодербетовский – Целинный – Кетченеровский), свидетельствуют о том, что флористический состав ценозов играет не самую большую роль в жизненности исследуемых ценопопуляций. Здесь на первое место, по нашим наблюдениям, выступают эдафические условия и отсутствие прессинга со стороны антропогенного фактора.
4. БИОХИМИЧЕСКИЙ СОСТАВ СОЛОДКИ ГОЛОЙ
По данным литературы (Саидов, Шамсувалиева, 1979), глицирризиновая кислота накапливается в коровой паренхиме корней и корневищах. Условия произрастания, возраст, фазы вегетации существенно влияют на определение максимума её содержания (Кустова, 1950; Муравьев, 1957; Гельбахиане, 1957; Мучник, 1975; Саурамбаев, 1983).
Для выяснения динамики содержания глицирризиновой кислоты, сахаров, флавоноидов в подземных органах солодки голой отбиралось различное сырье (ошкуренное (о/ш), неошкуренное (н/ш), собранное по 3 фазам вегетации (табл. 2). Исследования показали, что осенние корни и корневища солодки голой того же диаметра содержат больше глицирризиновой кислоты (14.76%), чем летние (10.82%). При определении суммы сахаров, нами было выявлено, что их содержание коррелирует с глицирризиновой кислотой. Так, летом, эта зависимость обратная: сахаров много (9.19%), а глицирризиновой кислоты - мало (10.82%), осенью наблюдается прямая зависимость: сахаров мало (6.65%), а глицирризиновой кислоты - много (14.76%).
Таблица 2
Динамика содержания глицирризиновой кислоты, сахаров, флавоноидов в подземных
органах солодки голой в зависимости от фазы вегетации, % от абс. сух. массы
| Фаза развития | Сырье | Биологически активные вещества | ||
| глицир.к-та | сахара | флавоноиды | ||
| Отрастание | н/ш | 7.67+1.30 | 7.80+2.12 | 1.15+0.50 |
| о/ш | 8.49+1.35 | 4.92+1.50 | 3.60+0.22 | |
| Цветение | н/ш | 10.82+1.81 | 9.19+1.05 | 3.41+0.85 |
| о/ш | 6.85+1.60 | 5.64+1.00 | 0.92+1.00 | |
| Плодоношение | н/ш | 14.76+0.50 | 6.65+1.40 | 2.62+0.46 |
| о/ш | 10.96+2.00 | 8.59+0.35 | 1.05+0.35 | |
Установлено, что содержание сахаров летом во флоэме увеличивается, а осенью часть их часть превращается в запасные питательные вещества и локализуется в ксилеме, другая часть идет на образование сапонинов, и содержание глицирризиновой кислоты увеличивается. Отмечено, что содержание флавоноидов минимально в фазу отрастания (1.15%), максимально в фазу цветения (3.41%), а в фазу плодоношения их содержание снижается.
Анализ минерального состава подземных органов солодки голой проводился по двум фазам вегетации и в различном сырье (табл. 3). Было отмечено, что содержание азота увеличивается в фазу цветения в неошкуренном сырье (3.38%), а в ошкуренном – его мало (2.67%). Содержание фосфора, калия и кальция увеличивается в фазу плодоношения в ошкуренном сырье (0.38%, 0.39%, 14.02 г/кг соответственно). Максимумы содержания магния, меди, марганца, цинка в неошкуренном сырье приходятся на летний период (5.22 г/кг, 9.63 мг/кг, 23.50 мг/кг, 20.63 мг/кг).
Таблица 3
Содержание минеральных элементов в солодковом корне,
в пересчете на абс. сух. массу (усредненные данные)
| Фаза развития | Сырье | N | P | K | Ca | Mg | Cu | Mn | Zn |
| % | г/кг | мг/кг | |||||||
| Цветение | н/ш | 3.38 | 0.23 | 0.32 | 12.37 | 5.22 | 9.63 | 23.50 | 20.63 |
| о/ш | 2.67 | 0.23 | 0.39 | 12.64 | 4.54 | 6.13 | 11.50 | 14.50 | |
| Плодоношение | н/ш | 3.08 | 0.32 | 0.35 | 13.62 | 1.91 | 5.56 | 14.75 | 20.50 |
| о/ш | 2.88 | 0.38 | 0.39 | 14.02 | 0.84 | 4.69 | 9.63 | 11.25 | |
Для определения наилучшего по содержанию биологически активных веществ (БАВ) и минеральных элементов лекарственно сырья был проведен анализ подземных органов солодки из наиболее крупных десяти ценопопуляций: 1.1, 1.2, 1.4, 2.1, 2.5, 2.6, 3.5, 3.7, 5.7, 6.2 (Табл. 4).
Таблица 4
Содержание разных веществ в подземных и надземных органах солодки голой, в пересчете на абс. сух. массу
| Цено- популяция | Орга- ны | Глицир. к-та | Сахара | Флаво- ноиды | Клет- чатка | Про- теин | Зола | N | P | K | Ca | Mg | Cu | Zn | Mn | Fe |
| % | г/кг | мг/кг | ||||||||||||||
| 1.1 | п | 12.12 | 7.04 | 2.78 | - | 12.37 | 8.39 | 1.98 | 0.090 | 0.48 | 10.27 | 6.21 | 5.35 | 14.04 | 38.52 | 717 |
| н | - | 11.20 | 1.70 | 21.4 | 17.38 | 5.67 | 2.78 | 0.200 | 0.81 | 13.39 | 6.15 | 4.72 | 18.90 | 22.08 | 151 | |
| 1.2 | п | 13.72 | 12.89 | 2.21 | - | - | 11.00 | 1.54 | 0.080 | 0.85 | 12.53 | 9.07 | 3.73 | 12.15 | 28.40 | 648 |
| 1.4 | п | 8.80 | 8.90 | 3.41 | - | 9.28 | 9.53 | 1.49 | 0.120 | 0.72 | 11.66 | 6.91 | 3.24 | 13.50 | 40.50 | 691 |
| н | - | 8.15 | - | 13.5 | 15.67 | 6.69 | 2.51 | 0.140 | 1.17 | 16.13 | 9.05 | 3.20 | 19.07 | 45.78 | 112 | |
| 2.1 | п | 8.60 | 9.30 | 1.30 | - | - | 10.39 | 1.70 | 0.054 | 0.72 | 21.17 | 12.42 | 13.09 | 15.52 | 35.46 | 475 |
| 2.2 | п | - | - | - | - | 11.37 | 12.38 | 1.82 | 0.110 | 0.97 | 18.83 | 9.42 | 9.63 | 12.04 | 42.80 | 695 |
| н | - | 11.20 | 1.40 | 15.1 | 24.75 | 8.42 | 3.96 | 0.310 | 1.24 | 10.27 | 5.27 | 11.37 | 48.82 | 55.64 | 407 | |
| 2.3 | п | 7.67 | 9.63 | 1.55 | - | 10.63 | 10.10 | 1.70 | 0.032 | 0.92 | 19.87 | 8.96 | 6.08 | 20.25 | 28.08 | 270 |
| н | - | 6.80 | 1.30 | 18.9 | 16.37 | 11.44 | 2.62 | 0.220 | 1.49 | 15.55 | 5.29 | 7.56 | 23.62 | 62.60 | 356 | |
| 2.5 | п | 7.70 | 7.10 | 1.60 | - | 9.62 | 13.52 | 1.54 | 0.054 | 0.27 | 22.90 | 11.66 | 7.16 | 15.52 | 29.16 | 421 |
| н | - | 10.72 | 1.37 | 20.4 | 16.37 | 10.30 | 2.62 | 0.180 | 1.15 | 11.23 | 6.80 | 9.45 | 20.52 | 70.20 | 238 | |
| 2.6 | п | 6.92 | 9.72 | 1.42 | - | 9.96 | 11.50 | 1.59 | 0.054 | 0.31 | 15.98 | 9.83 | 10.93 | 18.22 | 21.60 | 194 |
| н | - | 8.87 | 1.65 | 23.7 | 16.75 | 17.40 | 2.68 | 0.220 | 1.58 | 9.61 | 6.23 | 15.25 | 39.43 | 56.75 | 442 | |
| 3.2 | п | 4.10 | 7.80 | 1.30 | - | 12.71 | 7.15 | 2.03 | 0.070 | 0.30 | 9.84 | 10.37 | 16.98 | 27.42 | 32.10 | 214 |
| н | - | 10.60 | 1.20 | 12.9 | 24.98 | 8.57 | 4.00 | 0.290 | 1.06 | 14.26 | 7.77 | 11.61 | 50.62 | 104.70 | 173 | |
| 3.5 | п | 10.76 | 6.29 | 1.80 | - | - | 9.98 | 1.42 | 0.130 | 1.24 | 10.70 | 7.17 | 5.49 | 13.37 | 34.24 | 546 |
| 3.7 | п | 5.83 | 5.17 | 1.38 | - | - | 13.91 | 1.47 | 0.110 | 0.94 | 15.41 | 8.56 | 9.63 | 12.71 | 55.64 | 803 |
| 4.2 | п | - | - | - | - | 11.71 | 11.90 | 1.87 | 0.110 | 0.43 | 18.83 | 10.30 | 8.03 | 17.39 | 30.17 | 663 |
| н | - | 7.80 | 1.30 | 12.9 | 25.65 | 11.20 | 4.10 | 0.430 | 1.84 | 12.10 | 6.05 | 7.70 | 39.15 | 75.60 | 486 | |
| 5.7 | п | 9.50 | 11.80 | 1.60 | - | - | 9.50 | 1.25 | 0.053 | 0.66 | 15.93 | 9.82 | 8.34 | 14.60 | 33.73 | 528 |
| 6.1 | п | 7.90 | 11.50 | 1.50 | - | 7.86 | 10.3 | 1.26 | 0.280 | 0.53 | 19.26 | 7.70 | 10.56 | 25.41 | 35.31 | 460 |
| н | - | 11.50 | 1.40 | 12.8 | 23.96 | 6.87 | 3.83 | 0.270 | 1.16 | 10.80 | 5.94 | 8.24 | 36.45 | 100.4 | 151 | |
| 6.2 | п | 9.82 | 8.93 | 1.57 | - | - | 7.77 | 1.26 | 0.110 | 0.94 | 10.27 | 5.46 | 6.83 | 26.07 | 46.01 | 423 |
Параллельно проводился агрохимический анализ почв в изучаемых ценопопуляциях, который показал отсутствие засоления почти во всех образцах почвы.
Отмечено высокое содержание БАВ в подземных органах (п) солодки голой из ценопопуляций Юстинского района. Есть все основания растения из этих ценопопуляций рекомендовать в культуру. Наиболее высокое содержание сахаров отмечено у растений из ценопопуляций 1.2 и 5.7, наименьшее – в 3.7.
Проведенный анализ показал, что зольность образцов, а, следовательно, суммарное количество микроэлементов, из ценопопуляций Малодербетовского района существенно выше остальных. Высокое содержание азота отмечено для корней и корневищ солодки из ценопопуляций Юстинского и Малодербетовского районов. Для большинства растений из этих районов характерно наличие многочисленных клубеньков. Причем с увеличением увлажненности почвы возрастало их количество.
Анализ питательности, кормовой ценности надземной фитомассы солодки голой, собранной в фазу цветения в исследуемых районах, показал ее высокую значимость. Надземная часть (н) солодки голой имеет высокую питательность. В ней много протеина (15.67 – 25.65%), а также клетчатки (12.8 – 23.7%). Суммарное количество сахаров в изучаемых ценопопуляциях колеблется в пределах от 6.8 до 11.5%, наименьшее их содержание отмечено в ЦП2.3, наибольшее – в ЦП6.1. Содержание сахаров и флавоноидов в надземных органах выше, чем в подземных, что объясняется их защитной ролью. Сумма флавоноидов в районе исследования варьирует в пределах от 1.2 до 3.41%. Наибольшее их содержание в надземной фитомассе отмечено в ЦП1.1, подземной – в ЦП1.4.
В надземной массе солодки голой отмечено большее количество марганца, в частности в П3.2, П6.1 (104.7, 100.4 мг/кг соответственно), поэтому мы рекомендуем ее силосовать и добавлять в сено, с люцерной, а также в смеси с другими растениями. В ценопопуляциях, произрастающих в Малодербетовском районе, отмечено высокое содержание меди, что подтверждает отмеченную Т.А. Волхонской (2000) связь между содержанием в растениях БАВ и концентрацией в них макро- и микроэлементов. Минимумы меди, цинка, марганца, железа приходятся на надземную фитомассу солодки, произрастающей в Юстинском районе. В надземной фитомассе солодки голой аккумулируется больше макро- и микроэлементов, чем в подземной. Однако ряд биологического поглощения микроэлементов остается тем же: Fe>Mn>Zn>Cu. Различия отмечены в поглощении магния и кальция: в надземных органах больше накапливается магния, а в подземных – кальция.
5. СОВРЕМЕННОЕ СОСТОЯНИЕ И ОХРАНА ПОПУЛЯЦИЙ СОЛОДКИ ГОЛОЙ
Анализ данных, полученных методом установления эколого-ценотического градиента, а также многолетние наблюдения, позволили нам провести ранжирование исследованных ценопопуляций по современному состоянию на три группы: хорошее, удовлетворительное, критическое. При этом учитывалась занимаемая площадь, численность особей в популяции, процент генеративных побегов и другие признаки.
Хорошее состояние имеют ценопопуляции, произрастающие на территории Юстинского, Малодербетовского и Сарпинского районов. Удовлетворительное состояние ценопопуляций солодки отмечается преимущественно в Приютненском районе и в ряде ценопопуляций Целинного района. Малочисленные критические популяции, находящиеся на грани исчезновения, находятся в Кетченеровском районе.
Для того чтобы получить логические выводы и сравнить их с эмпирическими, изложенными выше, на основании коэффициента сходства Серенсена-Чекановского установлено наличие 30 видов, входящих в состав всех анализируемых ценопопуляций. Эти виды, по нашему мнению, составляют модельную ЦП (МЦ) (табл. 5). Для полного абстрагирования от количества видов, введенных в сопряженный анализ, учитывались только частные от коэффициента Бравэ-Пирсона (КВ) и их суммы (А – «–»КВ /, В – «+»КВ /, С – «0»КВ / ) в ценопопуляциях: 1.2, 1.4. 2.5, 3.3, 4.1, 4.5, 5.2, 5.4. Наблюдаемые состояния (приватное мнение) указаны в таблице под условным названием Группа1, а вычисляемые состояния (А<(В+С) – хорошее состояние, А=(В+С) – удовлетворительное, А>(В+С) – неудовлетворительное) показывают принадлежность ценопопуляции к определенному состоянию, обозначаемому как Группа2.
Таблица 5
Состояние ЦП солодки голой из различных районов РК
| Параметр | 1.2 | 1.4 | 2.5 | 3.3 | 4.1 | 4.5 | 5.2 | 5.4 | МЦ |
| Группа1 | хор. | хор. | хор. | неуд. | уд. | неуд. | неуд. | неуд. | – |
| Кол-во «–» КВ | 140 | 62 | 59 | 111 | 59 | 155 | 58 | 72 | 151 |
| Кол-во «+» КВ | 131 | 101 | 52 | 79 | 50 | 134 | 57 | 48 | 239 |
| Кол-во «0» КВ | 29 | 27 | 25 | – | 11 | 11 | 5 | – | 45 |
| 300 | 190 | 136 | 190 | 120 | 300 | 120 | 120 | 435 | |
| А | 0.467 | 0.137 | 0.434 | 0.584 | 0.492 | 0.517 | 0.483 | 0.6 | 0.347 |
| В | 0.437 | 0.532 | 0.382 | 0.416 | 0.417 | 0.447 | 0.476 | 0.4 | 0.549 |
| С | 0.097 | 0.142 | 0.184 | – | 0.092 | 0.037 | 0.039 | – | 0.103 |
| < = > | < | < | < | > | = | > | = | > | < |
| Группа2 | хор. | хор. | хор. | неуд. | уд. | неуд. | уд. | неуд. | хор |
Проведенные исследования показали сходство эмпирических выводов с логическими. Так, были правильно определены состояния всех, кроме 5.2, солодковых ценопопуляций. Ценопопуляция 5.2 Целинного района резко сокращает свою площадь из-за развернувшейся на окраине г. Элисты стройки. Несмотря на это связи между растениями еще достаточно высоки и данная ценопопуляция относится ко второй группе, состояние которой расценивается как удовлетворительное. К этой же группе также относится и ЦП4.1 Кетренеровского района, где возрастающий прессинг со стороны антропогенного фактора еще не нарушил связи между видами.
Наиболее часто в качестве лимитирующих факторов для большинства солодковых ценопопуляций выступают перевыпас скота, сенокошение, бессистемные заготовки, реже – ценотические взаимоотношения, где доминанты – плотнодерновинные или корневищные злаки угнетающе действуют на молодые растения. Угнетение популяций, по нашим наблюдениям, происходит и под действием различных групп фитофагов, которые в жаркую погоду активно поедают надземную массу солодки, а во время плодоношения откладывают яйца в бобы.
Ресурсы солодки голой в исследуемых районах отличаются значительно (табл. 6). Наибольший объем возможных заготовок (с периодичностью через 3-5 лет на одном месте) возможен в Юстинском (ЦП1.1, ЦП1.4) и Малодербетовском районах (ЦП2.5). Эксплуатационный запас в этих ценопопуляциях составляет 26.5 – 35.3 т. Объем заготовок не должен превышать 10% от эксплуатационного запаса (Триль, Гранкина, 1986), что составляет 2.65 – 3.53 т.
Необходимо отметить, что в пойменных ценопопуляциях Юстинского района солодка формирует многоярусную корневую систему, за счет которой восстановление корней и корневищ идет быстрее. Это дает нам право утверждать, согласно С.С. Сабоиеву и Х.С. Мастоншоевой (1991), что для заготовок лучше использовать пойменные заросли.
Таблица 6
Общие запасы и объемы возможных заготовок корней и корневищ солодки голой
| Участок | Урожайность, г/м2 | Площадь, га | Эксплуатационный запас, т | Объем возможных заготовок, т |
| 1.1 | 651.2+52.2 | 5 | 27.3 | 2.73 |
| 1.4 | 678.6+74.6 | 5 | 26.5 | 2.65 |
| 1.5 | 576.2+28.8 | 2 | 10.4 | 1.04 |
| 2.3 | 740.2+76.0 | 1.5 | 8.8 | 0.88 |
| 2.5 | 1943.8+90.0 | 2 | 35.3 | 3.53 |
| 3.2 | 645.6+80.4 | 3 | 14.5 | 1.45 |
| 3.8 | 1045.0+108.0 | 1 | 8.3 | 0.83 |
| 5.8 | 1165.2+163.2 | 2 | 16.8 | 1.68 |
В целом, объем возможных заготовок в исследуемых районах незначительный и заготавливать сырье в промышленных масштабах для удовлетворения потребностей по линии аптекоуправления нецелесообразно. Заготовки возможны лишь при введении солодки голой в культуру с учетом изложенного выше.
Несомненно, что при сохраняющемся прессинге со стороны антропогенного фактора, который нарушает естественное восстановление ценопопуляций, солодка голая станет уязвимым видом и перейдет из третьей (Бакташева, 2000) во вторую категорию редкости. Для организации охранных мер по сохранению солодковых ценопопуляций необходимо проводить мониторинг за структурой ценопопуляций G. glabra в РК, с последующей экологической экспертизой раз в 3-5 лет.
Проведенные наблюдения и анализ литературных сведений позволяют рекомендовать для восстановления естественных ценопопуляций солодки голой и соответствующей ей степной растительности метод «агростепи», предложенный Д.С. Дзыбовым (1996). Этот же метод можно рекомендовать для активного воспроизводства – биологической рекультивации нарушенных после выкопки корней и корневищ земель, а также для оздоровления ландшафта.
ВЫВОДЫ
- В РК солодка голая распространена спорадически, в основном по балкам Ергенинской возвышенности. Она входит в состав разнотравных, разнотравно-злаковых и злаковых ценозов. Наибольший урожай подземных органов солодки (1943.8+90 г/м2) отмечен в составе разнотравных ценозов.
- В исследованных ценопопуляциях выявлено 262 вида дикорастущих сосудистых растений, относящихся к 48 семействам и 168 родам. Процентное соотношение многолетников составляет 62.98%. Ведущая роль в сложении ценопопуляций принадлежит четырем экологическим группам (эвксерофитам, эвриксерофитам, мезоксерофитам и эвмезофитам). В целом, состояние большинства ценопопуляций солодки голой оценивается как «удовлетворительное».
- У солодки голой встречаются два типа перикарпия бобов, характеризующихся отсутствием экзогенных (эпидермальных) образований («голые бобы») и наличием шипов с железкой в верхней части («шиповатые бобы»).
- Установлено высокое содержание БАВ в подземных органах солодки голой из пойменных ценопопуляций Юстинского района. Выявлена локализация глицирризиновой кислоты в перидерме, а сахаров и флавоноидов в центральном цилиндре подземных органов. В фазе цветения надземная фитомасса солодки содержит много протеина (15.7 – 25.7%), клетчатки (12.8 – 23.7%), сахаров (6.8 – 11.5%). Отмечена высокая способность солодки аккумулировать такие минеральные элементы, как Mn и Fe.
- Оптимальными сроками заготовки солодкового корня в РК являются летний и осенние периоды.
СПИСОК ЛИТЕРАТУРЫ
* - публикация в печатном издании перечня ВАК РФ
- Алексеева Т.Б., Бакташева Н.М., Сангаджиева Л.Х. Сравнительный анализ отдельных компонентов химического состава лекарственного аптечного сырья солодки голой (Glycyrrhiza glabra L.) c местным, произрастающим на территории Целинного района республики Калмыкии // Сб. науч. тр. молодых ученых, аспирантов и студентов Калмыцкого государственного университета. Вып.1. – Элиста, 2003. – С. 4-5.
- Бакташева Н.М., Алексеева Т.Б. Обсуждение возможности применения растений флоры Калмыкии в составе тибетских лекарственных средств // Ботанические исследования в азиатской России: Матер. XI Съезда Русского Ботанического общества.Т.3 – Барнаул, 2003 – С. 3-4.
- Алексеева Т.Б. Анализ содержания глицирризиновой кислоты в подземных органах солодки голой в условиях Калмыкии // Матер. VIII молодежной конф. ботаников. Санкт-Петербург. – СПб., 2004. – С. 117.
- Алексеева Т.Б., Бакташева Н.М. Фитоценотический анализ популяции солодки голой в районе села Максимовка // Проблемы сохранения и рационального использования биоразнообразия Прикаспия и сопредельных регионов: Матер. 3 Международной заочной научной конф. / Ассоциация университетов Прикаспийских государств. Выпуск 2. – Элиста, 2005. – С. 7-9.
- Бакташева Н.М., Алексеева Т.Б. О сходстве систематической структуры флоры Калмыкии и Северного Прикаспия // Экологические проблемы и биоразнообразие Северного Прикаспия: Материалы Международной научно-практической конференции, посвященной 100-летию со дня рождения профессора В.В. Иванова– Уральск, 2005. – С. 42-45.
- Алексеева Т.Б., Бакташева Н.М., Унканжинов Г.Д. Особенности биохимического состава солодки голой // Роль особо охраняемых природных территорий в сохранении биоразнообразия: Матер. международной научно-практической конф., посвященной 10-летию Государственного природного заповедника «Ростовский». – Ростов-на-Дону, 2006. – С. 173 – 174.
- Алексеева Т.Б. Динамика содержания биологически активных веществ в подземных органах солодки голой в условиях Калмыкии // Матер. I (IX) международной конф. молодых ботаников. Санкт-Петербург. – СПб., 2006 – С. 232.
- Алексеева Т.Б., Бакташева Н.М. Эколого-биологическая характеристика растительных сообществ с участием солодки голой (Glycyrrhiza glabra L.) на примере северо-западных популяций Республики Калмыкия // Степи Северной Евразии: Матер. IV международного симпозиума. Оренбург – Оренбург, 2006. – С. 56-59.
- *Алексеева Т.Б. Состояние популяций солодки голой в Калмыкии // Научная мысль Кавказа. Спецвыпуск №5. – Ростов-на-Дону, 2006. – С. 83-86.
- Алексеева Т.Б. Продуктивность и биохимический состав солодки голой из популяций Волго-Ахтубинской поймы // Экология биосистем: проблемы изучения, индикации и прогнозирования. Матер. международной научно-практической конф., посвященной 75-летию Астраханского государственного университета. – Астрахань, 2007. – С. 3-5.
Подписано в печать 20.09. 2007. Формат 6084 1/16.
Усл. печ. л. 1,16. Тираж 110 экз. Заказ № 949
Издательство Калмыцкого университета.
358000 Элиста, ул. Пушкина, 11