

Алексеевна исследование нейрофизиологических механизмов последовательной слуховой маскировки в условиях сенсомоторной интеграции
На правах рукописи
___________
Павловская Марина Алексеевна
ИССЛЕДОВАНИЕ НЕЙРОФИЗИОЛОГИЧЕСКИХ МЕХАНИЗМОВ ПОСЛЕДОВАТЕЛЬНОЙ СЛУХОВОЙ МАСКИРОВКИ В УСЛОВИЯХ СЕНСОМОТОРНОЙ ИНТЕГРАЦИИ
03.00.13 - физиология
Автореферат
диссертации на соискание ученой степени
кандидата биологических наук
Ростов-на-Дону
2009
Работа выполнена на кафедре физиологии человека и животных Федерального государственного образовательного учреждения высшего профессионального обучения "Южный федеральный университет"
| Научный руководитель: | кандидат биологических наук, профессор Айдаркин Евгений Константинович |
| Официальные оппоненты: | доктор биологических наук, профессор Шульговский Валерий Викторович доктор биологических наук, профессор Владимирский Борис Михайлович |
Ведущая организация: Санкт-Петербургский государственный университет
Защита диссертации состоится "17" февраля 2009 г. в 14.00 час на заседании диссертационного совета Д.212.208.07 по биологическим наукам в Южном федеральном университете (344006, г. Ростов-на-Дону, ул. Большая Садовая, 105, ауд. 203).
С диссертацией можно ознакомиться в библиотеке Южного федерального университета (344006, г. Ростов-на-Дону, ул. Пушкинская, 148).
Автореферат разослан " 16 " января 2009 г.
Ученый секретарь диссертационного совета,
доктор биологических наук Колмакова Т.С.
Общая характеристика работы
Актуальность исследования. В психофизиологических исследованиях механизмов слухового восприятия наиболее актуальной является экспериментальная парадигма последовательной маскировки. Маскирующее влияние помехи приводит к повышению абсолютных (Ehmer, 1959; Green, Wier, 1984; Hawkins, Stivens, 1950) и дифференциальных порогов (Egan, 1948; Shatter, Moore, 1985), уменьшению субъективной громкости сигналов (Sharf, 1985), нарушению их распознавания (Kalokow, Stevens, 1977; Галунов, Королева,1988; Альтман, 1990; Crawley et al., 1994; Zeng, 1998; Zeng et al., 2001; Kovacs et al., 2005; Meese, 2004; Nishimura et al., 2004; Okamoto et al., 2005; Wojtczak, Viemeister, 2005; Robitaille, Jolicoeur, 2006) и скорости обработки информации (Miller et al., 1951). Сокращение маскировочного интервала (МИ) может приводить к полному подавлению восприятия тестового стимула, либо к ухудшению некоторых его признаков, что можно объяснить механизмами центральной и периферической маскировки.
Экспериментально показано, что при величине МИ от 0 до 50 мс господствуют закономерности периферической маскировки (Чистович, Иванова, 1959; Bekeshi, 1960; Levitt, Rabiner, 1967; Dealherage, Evans, 1969; Zwicker, Feldkeller, 1971; Zwicker, Shorn, 1978; Soderquist, Carstens, Frank, 1981; Гельфанд, 1984; Green, Wier, 1984; Галунов, Королева, 1988; Shamma, 1985), которые позволяют оценить вклад рецепторных систем в формирование слуховых образов за счет механизмов временной и пространственной суммации, латерального торможения, адаптации, рефрактерности и т.д. В диапазоне от 50 до 100 мс включаются механизмы сенсорной маскировки, основные эффекты которой базируются на принципах физиологической рефрактерности центральных анализаторных структур (Rabiner, 1967; Ward, 1967; Levitt, Dickson, 1983; Альтман, 1983; Shamma, 1985; Sheffers, 1984; Радионова, 1987). Именно в центральных отделах реализуются механизмы бинаурального взаимодействия, фильтрация и параллельная обработка сигналов с выделением различных их признаков (Галунов, Королева, 1988). При маскировочном интервале 200-300 мс возникает интерференция когнитивных процессов, связанных с анализом чередующихся целевого и маскировочного стимулов («когнитивная» маскировка), причиной которой могут быть нарушения кратковременного следа памяти о целевом стимуле (Невская, 1966, 1967; Глезер, 1975; Каменкович, 1975; Леушина, 1976, 1978; Борисова, 1983; Бахман, 1989), а также процессов переключения внимания с анализа целевого стимула на маскер и обратно.
В литературе влияние вариации стимулов в условиях маскировки на эффективность восприятия оценивается по параметрам времени реакции (ВР) (Lappin, Eriksen, 1964; Harrison, Fox, 1966; Mitchell, 1972, Bernstein et al., 1973; Matterson, Flaherty, 1976; Catlin, Neville, 1976; Кроль, Таненгольц, 1976; Eriksen, Schultz, 1978; Proctor et al., 1983) и связанных с событием потенциалов (ССП) (Kramer, Teas, 1982; Lusky, Rupert, 1982; Burkard, Hecox, 1987; Owen, Burkard, 1991; Вайтулевич, Кальницкая, Пак, 1997; Altman et al., 2000; Bekhterev et al., 2002; Petropavlovskaya, 2005).
Известно, что последовательность этапов сенсомоторной интеграции (СМИ) реализуется за счет вертикальных (корково-подкорковых) и горизонтальных (корково-корковых внутри- и межполушарных) возбудительных и тормозных влияний (Деглин, 1975; Павлова, 1980; Бианки, 1989; Брагина, Доброхотова, 1977; Жаворонкова, 2006), механизмы и особенности взаимодействия которых до конца не изучены. В настоящее время накапливается все больше свидетельств в пользу того, что асимметричная локализация корковых генераторов приводит к латерализации когнитивных функций, которая обеспечивается не только за счет особенностей кортикальных систем правого и левого полушарий, участвующих в их организации, но и за счет подкорковых структур, модулирующих уровень активации (Корсакова, Московичюте, 1988; Московичюте, 1998). Структурно-функциональная асимметрия мозга касается доминирования сенсорных и моторных процессов в левом полушарии, а их взаимодействия – в правом (Отмахова, Ковалев, 1988). При этом левополушарная асимметрия, возможно, реализуется при участии активирующей системы ствола мозга, а правополушарная – диэнцефальных и лимбических структур мозга (Брагина, Доброхтова, 1977).
Несмотря на имеющиеся многочисленные исследования феномена маскировки, общепризнанной концепции, объясняющей ее нейрофизиологические механизмы, особенно на «когнитивном» уровне, нет. Следовательно, наиболее актуальным в настоящее время является описание скорости сенсорной обработки и динамики когнитивных процессов, механизмов их интеграции на различных уровнях корково-подкоркового взаимодействия, обеспечивающих анализ целевых стимулов и маскера в условиях когнитивной маскировки, а также чувствительности различных стадий анализа целевого стимула (ожидание, восприятие, принятие решения и т.д.) к воздействию маскера.
Цель работы: Исследование нейрофизиологических механизмов последовательной центральной слуховой маскировки в условиях сенсомоторной интеграции по параметрам времени реакции и связанных с событием потенциалов.
Задачи исследования:
- Изучить зависимость ВР бинарного различения в условиях прямой (ПМ) и обратной (ОМ) последовательной маскировки от вероятности целевых стимулов и длительности МИ.
- Изучить динамику основных компонентов ССП в условиях ПМ и ОМ при равно- и разновероятном предъявлении целевых стимулов в зависимости от длительности МИ.
- Изучить динамику изопотенциальных карт ССП в условиях ПМ и ОМ при равно- и разновероятном предъявлении целевых стимулов в зависимости от длительности МИ.
- Изучить зависимость ритмических составляющих ССП в условиях ПМ и ОМ при равно- и разновероятном предъявлении целевых стимулов в зависимости от длительности МИ.
- Сформулировать представление о психо-физиологических механизмах восприятия целевого стимула в условиях последовательной слуховой маскировки.
Научная новизна результатов исследования.
- Показано, что скорость и асимметрия (ВР левой рукой меньше) распознавания целевого стимула пропорциональны его вероятности и больше в условиях ПМ (по сравнению с контролем и ОМ).
- Показано, что слуховое различение целевых стимулов в условиях бинарного выбора обеспечивается взаимодействием дельта колебаний, лежащих в основе CPV, CNV, НР и Р3б компонентов, и тета колебаний, участвующих в формировании N1, P2, N2 и P3а компонентов ССП. Ухудшение распознавания целевого стимула в условиях ОМ связано с воздействием маскера на постстимульные тета волны, а улучшение в условиях ПМ – с воздействием на достимульные дельта колебания.
- Показано, что динамика функциональной межполушарной асимметрии в условиях анализа целевых стимулов (контроль, ОМ) на стадиях ожидания (в центральной области) и различения (правой верхневисочной и левой нижнетеменной областях) характеризуется наличием двух негативных фокусов с характеристиками временного дельта диапазона колебаний с преобладанием правополушарного (особенно на редкий стимул). В условиях ПМ наблюдается левополушарная негативность в центральной области на стадии ожидания маскера с последующей инверсией асимметрии на этапе восприятия целевого стимула и последующем преобладании ее в правом полушарии, что обеспечивало более быстрое распознавание целевого стимула за счет демпфирующей функции левого полушария.
- Сокращение МИ в ситуациях ПМ и ОМ приводило к постепенному сдвигу CNV в теменную область, уменьшению асимметрии за счет слияния двух негативных фокусов, исчезновению Е-волны в центральной области, с последующим возникновением О-волны во фронтальном фокусе, что было связано с ухудшением процесса различения целевых стимулов, особенно в условиях периферической маскировки.
Научно-практическая значимость работы. В работе на основании изучения нейрофизиологических механизмов описано воздействие маскера на различные стадии анализа целевых стимулов при формировании и реализации СМИ. Так в условиях ОМ сокращение МИ приводит к постепенному нарушению стадий организации и реализации двигательной реакции (МИ 300 мс), принятия решения (МИ 200 мс), различения (МИ 100 мс) и обнаружения (МИ 50 мс), что отражалось в снижении скорости различения целевых стимулов. В процедуре ПМ маскер приходился на различные этапы достимульной подготовки (МИ 100-300 мс), что приводит к улучшению выполнения тестовой процедуры. Изменение скорости различения в экспериментальной парадигме последовательной слуховой маскировки связано с функциональной межполушарной асимметрией (ФМА) на различных стадиях СМИ, что выражалось в преобладании левого (ожидание маскера при ПМ) или правого (ожидание и различение целевого стимула при ОМ и ПМ) полушарий, а также от характера взаимодействия таламо-фронтальных (сенсорно-специфический анализ, отражающийся в развитии N1, P2, N2 и P3a компонентов ССП) и таламо-париетальных (основная функция которых состоит в модуляции заинтересованных структур мозга и определяющая развитие CPV, CNV, НР и P3б) систем внимания, деятельность которых отражается в активности тета- и дельта- колебаний, соответственно, лежащих в основе формирования указанных компонентов ССП.
Показана перспективность использования маскировочной парадигмы в качестве коррекционной процедуры воздействия на функциональное состояние человека-оператора в ходе рабочей смены, а также для оценки его помехоустойчивости.
Основные положения, выносимые на защиту.
- Скорость распознавания целевых стимулов в условиях последовательной маскировки определяется порядком следования маскера и целевых стимулов, их вероятностью и латерализацией двигательного ответа.
- В условиях ОМ воздействие маскера разрушает основные стадии СМИ, увеличивая скорость распознавания независимо от МИ. Маскер при ПМ, воздействуя только на стадию достимульной подготовки, повышает лабильность слуховой системы.
- Функциональная межполушарная асимметрия на ранних стадиях СМИ определяет высокую эффективность распознавания, с одной стороны, за счет преобладания негативных фокусов на стадиях ожидания и различения целевых стимулов в правом полушарии при активации физиологических механизмов внимания, с другой - за счет демпфирующей функции левого полушария, отражающейся в инверсии левополушарного доминирования негативного фокуса на стадии ожидания маскера в правополушарное – на стадии восприятия целевого стимула (при ПМ).
- Дельта колебания, обеспечивающие необходимый уровень возбудимости заинтересованных структур соответствуют развитию CNV, CPV, НР и Р3б компонентам ССП, а тета-волны, описывающие специфический анализ соответственно - N1, P2, N2, P3a компонентами ССП.
- Эффект последовательной слуховой маскировки связан с взаимодействием таламо-париетальной и таламо-фронтальной систем внимания, перераспределяющих его ресурсы при анализе целевых и маскировочных стимулов.
Апробация диссертационной работы. Результаты, полученные в данной работе, использовались при выполнении научно-исследовательских тем «Исследование нейрофизиологических механизмов взаимодействия зрительной, слуховой, вестибулярной и обонятельной систем при различных видах когнитивной деятельности» (№ государственной регистрации 10.1.06), «Исследование нейрофизиологических механизмов функциональной межполушарной асимметрии, лежащей в основе общей и локальной активации мозга в онто- и филогенезе» (№ гос. рег. 5.5.06), «Физиологическое, психологическое и кибернетическое изучение деятельности сигнальных систем организма в процессе выработки и реализации адаптивного поведения в проблемной ситуации» (№ гос. рег. 1.2.06).
Материалы диссертации представлены на VII-й, VIII-й и IX-й Всероссийских медико-биологических конференциях молодых исследователей «Человек и его здоровье» (Санкт-Петербург, 2004-2006), I-й и II-й ежегодных конференциях студентов и аспирантов базовых кафедр ЮНЦ РАН (Ростов-на-Дону, 2005, 2006), III-й и IV-й Межрегиональных научных конференциях студентов, аспирантов и молодых ученых «Молодежь XXI века – будущее Российской науки» (Ростов-на–Дону, 2005, 2008), 14-й Международной конференции по нейрокибернетике «Проблемы нейрокибернетики» (Ростов-на-Дону. 2005), XII-м Международном симпозиуме "Эколого-физиологические проблемы адаптации" (Москва, 2007), XIV-й и XV-й Международных конференциях студентов, аспирантов и молодых ученых "Ломоносов» (Москва, 2007, 2008), IX-й и XI-й Всероссийских медико-биологических конференциях молодых исследователей «Фундаментальная наука и клиническая медицина» (Санкт-Петербург, 2007, 2008), II-й и III-й Международных Междисциплинарных Конгрессах «Нейронаука для медицины и психологии» (Neuroscience for medicine and Psychophysiology Congress), (Украина, Судак, 2007, 2008), 14-ом Международном конгрессе психофизиологов (Санкт-Петербург, 2008), II-й Всероссийской научно-практической конференции «Функциональное состояние и здоровье человека» (Ростов-на-Дону, 2008), XII-й научной конференции молодых ученных по физиологии ВНД и нейрофизиологии (Москва, 2008), научной конференции «Актуальные вопросы функциональной межполушарной асимметрии и нейропластичности» (Москва, 2008).
Работа была апробирована на совместном заседании кафедры физиологии человека и животных биолого-почвенного факультета и УНИИ валеологии Южного федерального университета, на заседании Ученого Совета НИИ НК им. А.Б. Когана Южного федерального университета.
Публикации. По теме диссертации опубликовано 30 работ (из них 6 статей в журнале, рекомендованных ВАК РФ) и 1 патент на изобретение А61В 5/00, № 2007112370/14 от 20.12.2008); объемом 2,663 п.л.
Структура и объем диссертации. Диссертация изложена на 186 страницах машинописного текста, состоит из введения, 4 разделов (обзор литературы, методика, результаты, обсуждение), выводов и библиографического указателя, включающего 155 источников, из которых 126 – иностранных. Работа иллюстрирована 22 рисунками и 2 таблицами.
Материалы и Методы исследования
В исследовании принимали участие студенты и аспиранты ЮФУ в возрасте от 20 до 26 лет. В общей сложности было обследовано 27 человек, все правши.
В ходе электрофизиологического обследования испытуемый располагался в удобном кресле в звукоизолированной комнате, где выполнял ряд тестовых процедур. Слуховые стимулы предъявлялись одновременно с помощью двух динамиков, располагавшихся в 5 см от каждого уха. Для фиксации моторных реакций использовался кнопочный манипулятор «мышь».
Электрофизиологические методы исследования
Регистрация ЭЭГ, ВР и режим стимуляции реализовались при помощи компьютерного энцефалографа-анализатора «Энцефалан-131-03» (Таганрог, Россия). ЭЭГ-активность отводилась в 21 стандартном отведении (система 10-20) относительно объединенных ушных электродов и полосой пропускания 0,5-70 Гц. Индифферентный электрод располагался на лбу. Оцифрованная ЭЭГ и ВР экспортировались в MATLAB, где проводилась их дальнейшая обработка.
Схема психофизиологического тестирования
В качестве тестовых процедур были использованы ситуации ОМ и ПМ, в которых необходимо было дифференцировать два различных по частоте (1000 и 1200 Гц) тональных стимула интенсивностью 60 дБ и длительностью 30 мс. Согласно инструкции, на тестовый стимул частотой 1200 Гц необходимо было нажимать левой рукой на соответствующую клавишу манипулятора «мышь», а на стимул 1000 Гц – правой. В качестве маскера использовали тональный стимул 30 мс длительности, частотой 1100 Гц и интенсивностью 90 дБ. Для оценки влияния маскера на эффективность выполнения тестового задания использовался фиксированный ряд МИ 300, 200, 100 и 50 мс. В первой серии исследования вероятность целевых стимулов разной частоты была одинакова и составляла 0,5, во второй серии – вероятность одного из тестовых стимулов (1200 Гц) составляла 0,85, а другого (1000 Гц) – 0,15 («odd-ball» парадигма). Интервал между парами стимулов (тестовый стимул - маскер) составлял 4 с с девиацией 20%. В аналогичных контрольных сериях маскер отсутствовал.
Обработка данных
Вычислялись средние арифметические значения ВР, ошибка среднего, стандартное отклонение, доверительные интервалы с уровнем достоверности 0,95, а также процент правильных ответов для каждой руки. На основе анализа данных были построены графики зависимости среднего значения ВР от МИ.
Безартефактные участки ЭЭГ (режекция участков ЭЭГ с векодвигательными и ошибочными реакциями) подвергали полосовой фильтрации в диапазоне от 0,5 до 24 Гц. Частотные интервалы фильтрации для получения ритмических колебаний, соответствующих основным компонентам ССП, составляли 0,5-3 Гц для дельта, 4-7 Гц – тета, 8-13 Гц – альфа и 14-24 Гц- бета диапазонов. Эпоха анализа ЭЭГ для получения ССП составляла 1000 мс (100 мс до момента предъявления первого стимула в паре и 900 мс после). Количество усреднений ССП составляло не менее 60 реализаций.
Анализ ССП заключался в оценке амплитудно-временных показателей в исследуемом интервале. Амплитуда основных компонентов ССП измерялась относительно изолинии (средняя величина достимульного интервала длительностью 100 мс). Пиковый ЛП измерялся от начала предъявления стимула до ближайшей точки экстремума соответствующего компонента. Точность измерения временных параметров составляла 4 мс, а амплитудных – 0,04 мкВ. Для анализа были выбраны следующие компоненты звукового ССП: CNV, CPV (-200 - 0 мс), N1 (90 - 110 мс), P2 (160 - 180 мс), НР (160 - 180 мс), N2 (180 - 220 мс), P3 (320 - 350 мс) – для целевого и маскирующего стимулов. Метод изопотенциальных карт ССП и их ритмических компонентов заключался в построении динамики распределения мгновенных значений амплитуд компонентов ССП с шагом 20 мс, отражающих поэтапную обработку информации.
Для выяснения влияния исследуемых факторов на двигательные и электрофизиологические параметры использовался дисперсионный анализ ANOVA, где зависимыми переменными были амплитуда, ЛП компонентов ССП и ВР, а независимыми – отведение, испытуемый, вероятность целевых стимулов, тестовая процедура.
Результаты исследования
Исследование зависимости ВР от особенностей слуховой последовательной маскировки
Дисперсионный анализ (ANOVA) зависимости ВР от особенностей последовательной звуковой маскировки показал, что ВР при ПМ достоверно меньше, чем при ОМ (F=453,67, где р<0,000). ВР при МИ 300 мс было статистически значимо меньше ВР, полученном на МИ 100 и 50 мс (F=44,49, где р<0,000). На частый стимул (р=0,85) ВР было достоверно меньше, чем при вероятности 0,5 и 0,15 (F=458,52, где р<0,000).
Анализ скорости распознавания целевого стимула по ВР показал, что в контрольной серии статистически значимые изменения отсутствовали (рис. 1 А, В). Добавление маскирующего стимула привело к статистически значимому ухудшению выполнения сенсомоторной деятельности в условиях ОМ (рис. 1 А, В) и улучшению в случае ПМ (рис. 1 Б, Г), относительно контрольной серии бинарного различения.
Следует отметить эффект асимметрии ВР в условиях ПМ при равновероятном предъявлении целевых стимулов, что наблюдалось на всех исследуемых МИ и было связано с более коротким ВР левой руки. В условиях ОМ и в контроле преобладание ВР одной из рук не наблюдалось.
Для оценки значимости процессов произвольного и непроизвольного внимания была использована «odd-ball» парадигма (Наатанен, 1998; Айдаркин, 2007) при выполнении маскировочных тестовых процедур, которая позволила изменить их соотношение. Увеличение вероятности целевого стимула (р=0,85) приводило к сокращению ВР, выполняемой левой рукой в контрольной серии и при ОМ (Рис. 1 В) и, в большей степени, при ПМ (Рис. 1 Г), приближаясь к величинам, характерным для ПСМР (реакция обнаружения). В данных условиях асимметрия появляется при ОМ, а при ПМ усиливается, что связанно с доминированием левой руки. При предъявлении редкого (р=0,15) целевого стимула наблюдалось ухудшение выполнения деятельности.
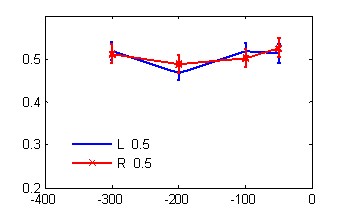
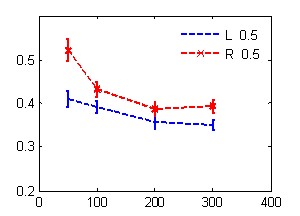

А Б

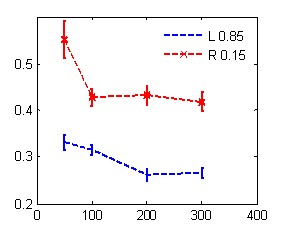

В Г
Рис. 1. Зависимость времени реакции от маскировочного интервала в условиях обратной (А, В) и прямой (Б, Г) маскировки с одинаковой (А, Б) и разной (В, Г) вероятностью предъявления целевых стимулов.
По оси абсцисс – МИ, мс; по оси ординат - время реакции, с. Линии с маркером - значения ВР при нажатии правой рукой, без маркера – левой рукой. Слева на графиках А, В показаны значения времени реакции, полученные в контрольной серии без маскера.
Сокращение МИ в условиях равно- и разновероятной стимуляции приводило к статистически значимому монотонному росту ВР.
Следовательно, скорость различения целевых стимулов повышается при увеличении МИ и вероятности их предъявления в ряду тестовых процедур – ОМ, контроль, ПМ, особенно при выполнении их левой рукой.
Анализ компонентов ССП в зависимости от особенностей слуховой последовательной маскировки
Анализ параметров ССП показал, что начальная стадия подготовки к анализу звуковых стимулов в условиях последовательной маскировки была связана с развитием негативной контингентной волны на целевой стимул (CNV) в центральной области, и/или позитивной волной (CPV) в лобной области при анализе маскера.
Стадия сенсорного анализа физических параметров стимула соответствовала развитию компонента N1. Сравнение его амплитудно-временных характеристик показало меньшую выраженность N1 в ответ на маскер по сравнению с N1 на целевой стимул.
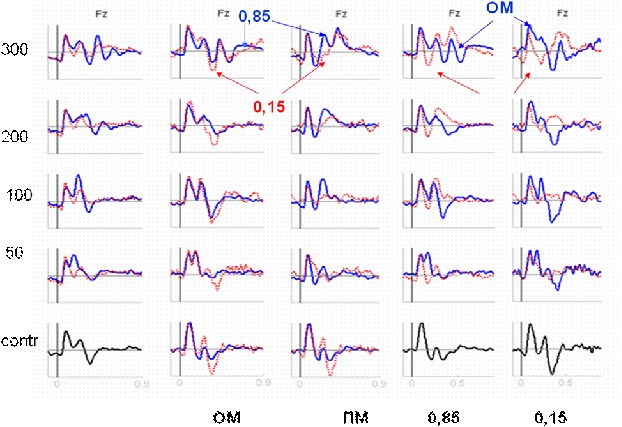

Рис. 2. Динамика компонентов ССП в условиях последовательной маскировки при изменении вероятности целевого стимула.
Слева указан МИ, мс. По оси абсцисс – время в с, по оси ординат – потенциал в мкВ. ОМ и ПМ – обратная и прямая маскировка; contr – контроль; 0,5, 0,85 и 0,15 – вероятности целевого стимула.
На этапе различения целевых стимулов формировалась негативность рассогласования (НР), развивавшаяся на заднем фронте N1. Увеличение выраженности НР при усложнении тестовых процедур приводило к редукции компонента Р2. Вслед за НР развивался негативный компонент N2, локализованный во фронтальной области.
В рамках «odd-ball» парадигмы анализ конфигурации ССП (рис. 2) показал дальнейший рост НР на редкий целевой стимул с последующим появлением мощной позитивной волны Р3б.
Сокращение МИ в условиях ОМ было связано с воздействием маскера на компонент Р3, связанного с организацией и реализацией двигательного акта (МИ 300 мс), на процесс принятия решения (МИ 200 мс), стадии различения (МИ 100 мс) и обнаружения (МИ 50 мс). В условиях ПМ (МИ 300-100 мс) маскер, попадающий на стадию ожидания целевого стимула, оптимизировал СМИ и обеспечивал сокращение времени выполнения теста.
Отмечено формирование медленной позитивно-негативно-позитивной волны (CPV-CNV-НР-Р3б), модулирующей основные стадии сенсорно-специфического анализа целевого стимула и маскера (N1, P2, N2). Уменьшение МИ при ОМ приводило к ослаблению медленных волн и деформации, указанных выше компонентов и, как следствие, к увеличению ВР.
Следовательно, парадигма маскировки представляет процедуру воздействия маскера на последовательные стадии СМИ начиная с достимульной подготовки (ПМ), стадии обнаружения, различения, принятия решения до организации и реализации двигательного акта (ОМ). Сокращение МИ приводило к интерференции компонентов ССП анализа целевого стимула с компонентами, развивающимися в ответ на маскер. Изменение вероятности целевых стимулов было связано с ростом выраженности модуляционных волн ССП с преобладанием позитивных процессов на этапе анализа маскировочного стимула и негативных компонентов на этапе анализа целевого стимула.
Исследование динамики изопотенциальных карт основных компонентов ССП в зависимости от особенностей слуховой последовательной маскировки
Метод построения изопотенциальных карт основных компонентов ССП (рис. 3, 4) позволяет рассматривать их как самостоятельные топологические структуры (процессы), мигрирующие по поверхности коры и отражающие изменение корково-подкорковых (вертикальные связи) и корково-корковых (горизонтальные связи) отношений (Айдаркин, 2007).
На стадии ожидания целевого стимула (на рис. 3 до 60 мс) обнаружено одновременное существование контингентной негативной волны, состоящей из двух ФМВ, локализованных в центральной области и позитивной волны в лобной области. Снижение вероятности целевого стимула в условиях последовательной маскировки приводило к ярко выраженному доминированию негативного фокуса в правом полушарии.
Сенсорная активация коры, соответствующая развитию компонента N1 была связана со слиянием двух негативных фокусов CNV и перемещением его из центрально-теменной области во фронтальную (рис. 3 от 80 до 120 мс).
Стадия распознавания целевых стимулов, представленная НР, сопровождалась расщеплением фронто-центральной негативности на два фокуса (рис. 3 от 140 до 180 мс), смещающихся в правую верхневисочную (доминирующей при разновероятной стимуляции) и левую нижнетеменную области. Оба фокуса находились в местах пересечения дорсальных и вентральных путей обработки слуховой информации (Hall, 2003).
Компонент Р2 ССП был локализован в фронто-центральных отведениях и хорошо выражен при анализе маскера (рис. 4 от 160 до 180 мс), конкурировал с НР на этапе анализа целевого стимула. Сокращение МИ приводило к его редукции.
Процедура подготовки к реализации СМИ отражалась в развитии волны Р3. Локализация данного компонента связана с формированием двух типов позитивности в лобно-центральной (Р3а, рис. 4) и теменной (Р3б, рис. 4) областях головного мозга, ФМВ которых были расположены сагиттально.
Анализ маскера связан с развитием позитивных процессов, отражающихся в развитии CPV (рис. 4 до 40 мс) и компонента Р2 (рис. 4 от 160 до 220 мс) при ПМ и интерференцией указанных элементов с Р3б при ОМ (рис. 3 от 400 до 560 мс).

Рис. 3. Динамика изопотенциальных карт основных компонентов ССП в условиях ОМ.
Светлым тоном показан максимум негативности, темным – позитивности. Цифры под картой указывают на время ее возникновения от момента подачи стимула, мс. Вероятность целевого стимула р=0,15, МИ составил 300 мс.

Рис. 4. Динамика изопотенциальных карт основных компонентов ССП в условиях ПМ.
Обозначения как на рис. 3.
Разновероятная стимуляция целевыми стимулами в условиях ОМ и ПМ привела к росту компонентов CNV, НР и Р3, что усиливало описанную выше ФМА, на соответствующих стадиях ожидания, различения и реализации СМИ.
Следовательно, анализ изопотенциальных карт основных компонентов ССП показал, что обработка целевых стимулов и маскера представлена динамикой взаимодействия таламо-париетальных, вносящих вклад в формирование CNV, НР и Р3б компонентов ССП и фронто-таламических, участвующих в формировании CPV, N1, N2, P2 и P3а компонентов, систем, регулирующих последовательные циклах организации и реализации СМИ. При этом этап ожидания и различения характеризовались правополушарным доминированием негативных фокусов в центральной области, обеспечивающих более эффективную реализацию задачи левой рукой.
Анализ зависимости изопотенциальных карт ритмических составляющих ССП от особенностей последовательной слуховой маскировки
Разложение (фильтрация) слуховых ССП на ритмические составляющие показало, что все основные их изменения связаны с дельта- (рис. 5) и тета-(рис. 6) колебаниями, суперпозиция изопотенциальных карт которых практически полностью совпадает с соответствующей картой целостного ССП (рис. 4).
Наиболее чувствительным ко всем указанным факторам экспериментальных парадигм были дельта-колебания (рис. 6), которые включали в себя последовательность медленных компонентов ССП, возникающих на следующих этапах СМИ: формирование (CNV), восстановление (НР) и завершение (Р3б).
Основная дельта-волна (рис. 5) ССП представляла собой комплекс двух асимметричных центрально-негативных колебаний, который возникал задолго до предъявления целевого стимула (CNV, Е-волна), перекрываясь с НР и сохраняясь в виде исходного комплекса, состоящего также из двух центрально-негативных ФМВ с правополушарным доминированием, и угнетался на этапе развития позитивно-теменной волны Р3б, восстанавливаясь после ее исчезновения. Данное восстановление CNV можно рассматривать в качестве варианта «постимперативной негативной волны».
Исследование особенностей тета-колебаний ССП (рис. 6) показало, что они имели лобное происхождение с локализацией ФМВ в областях, характерных для тета-ритма ЭЭГ, больше связаны с анализом пускового целевого слухового стимула и соответствовали развитию фазических экзогенных ответов в виде компонентов N1, Р2, N2 и Р3a. В условиях ОМ при сокращении МИ основные изменения были связаны с тета-колебаниями, определяющих развитие основных компонентов ССП, что приводило к нарушению соответствующих стадий СМИ: подготовка и реализация двигательного ответа (при МИ 200-300 мс), принятие решения (МИ 100 мс), обнаружение (МИ 50 мс).
ПМ в основном приводила к изменению дельта-колебаний ССП.

Рис.5. Динамика ритмических дельта компонентов ССП в условиях ОМ.
Вероятность целевого стимула р=0,15, МИ 300 мс, исходная динамика изопотенциальных карт основных компонентов ССП показана на рис. 3.

Рис. 6. Динамика ритмических тета компонентов ССП в условиях ПМ.
Вероятность целевого стимула р=0,15, МИ 300 мс, исходная динамика изопотенциальных карт основных компонентов ССП показана на рис. 4.
Появление маскера (МИ 100-300 мс) в условиях ПМ было связано с доминированием левого ФМВ в комплексе центральных негативных компонентов. При этом дальнейшее восстановление исходной правосторонней асимметрии наблюдалось на этапе развития НР в ответ на целевой стимул.
Сокращение МИ в обеих тестовых ситуациях приводило к постепенному сдвигу CNV в теменную область, уменьшению асимметрии за счет слияния двух фокусов, с последующей заменой центрально-теменной Е-волны, развивающейся после завершения компонента Р3б, на О-волну с ФМВ в лобной области.
Таким образом, нейрофизиологический механизм восприятия слухового стимула обеспечивается взаимодействием двух систем – тета и дельта ритмических колебаний, лежащих в основе формирования основных компонентов ССП. Механизм ОМ связан с нарушением цепочки тета-колебаний ССП, восстановление которой требует одинакового времени. В условиях ПМ воздействие маскера направлено на дельта-колебания ССП и выражается в изменении Е-волны, ФМВ которой в момент восприятия маскера смещается в левое полушарие, а на этапе анализа целевого стимула – в правое, восстанавливая исходную ФМА, характерную для ОМ и контроля.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Анализ экспериментальных результатов, полученных в условиях последовательной слуховой маскировки, показал, что увеличение ВР при ОМ, вероятно, свидетельствует о сокращении необходимых для опознания ресурсов внимания, что связано с отвлечением их на восприятие маскера. В условиях ПМ снижение ВР, скорее всего, происходит за счет предупреждающей функции маскера, перераспределяющей ресурсы внимания и оптимизирующей процесс различения целевых стимулов.
Увеличение вероятности целевого стимула особенно в условиях ПМ приводило к сокращению ВР, выполняемой левой рукой, что связано с доминированием процессов в правом полушарии, лежащих в основе активации непроизвольного внимания (Суворов, Шуваев, 2004) и организации соответствующей двигательной реакции, интерференция которых в одной гемисфере существенно улучшает выполнение СМР. Ухудшение различения при предъявлении редкого целевого стимула, возможно, происходит за счет усложнения реагирования и перехода от простой реакции обнаружения к сложной реакции различения, что связано с привлечением дополнительных ресурсов произвольного внимания, которые необходимы для восстановления образа редкого целевого стимула и реализации СМИ (Айдаркин, 2007). Сокращение МИ приводило к монотонному росту ВР, что связано с активацией аналогичных механизмов центральной и периферической маскировки.
Сенсорно-специфические процессы анализа стимулов (обнаружение, различение, принятие решения), сформированные тета-колебаниями и соответствующие компонентам N1, P2, N2 и P3а ССП, были локализованы в фронто-центральных отведениях, что подтверждает справедливость теории о циклической природе возникновения ощущения (Иваницкий, 1987). Для слуховой стимуляции тета-колебания ССП, вероятно, связаны с квантованием экзогенных (восприятие) и эндогенных (память) процессов (Симонов, 1979), что определяется в первую очередь влиянием на кору гиппокампальных структур. Предъявление маскера при ОМ приводит к нарушению цепочки последовательных стадий СМИ, в основе которых лежат однотипные циклические корково-гиппокампальные взаимодействия (Gaarder et al., 1966), для восстановления которых требуется приблизительно одинаковое время и снижению скорости различения по разным причинам (слияние физических признаков стимулов, нарушение сенсорного и моторного принятия решения, сенсорной рефрактерности). Развитие тета-компонентов ССП во фронтальной системе свидетельствует о том, что последовательность этапов СМИ контролируется из одного источника, связанного с таламо-фронтальной системой внимания.
Выше перечисленные этапы восприятия стимулов сопровождались модуляцией медленной дельта-волной позитивно-негативно-позитивной (в условиях ПМ формирующей цепочку компонентов CPV-CNV-НР-Р3б ССП, и негативно-позитивной (при ОМ CNV-НР-Р3б), отражая тонические (эндогенные) неспецифические процессы, связанные с концентрацией ресурсов произвольного внимания на анализе целевого стимула и повышающей пороги восприятия на маскер. Особое внимание в процессах модуляции сенсорных процессов обработки информации следует уделить двум морфофункциональным системам внимания: задней и передней (Posner, Petersen, 1990, 1994; Kimura, 1987, 2000; Davidson, 1993; Goldberg et al., 1994; Симонов, 1996; Вольф, 1998, 2000). Ключевыми структурами передней системы внимания (эндогенная, таламо-фронтальная) являются передняя цингулярная извилина и дорзолатеральная префронтальная кора, которые связаны с базальными ганглиями, лимбическими и таламическими ядрами, обеспечивая привлечение внимания к анализу стимулов за счет ретикулярного ядра таламуса (nucleus reticularis) (Watson et al., 1981; Mesulam, 1981, 1985; Weintrayb, Mesulam, 1989) и выполнению двигательной реакции с участием медиодорзального ядра таламуса (Мачинская, 2003). Задняя система (экзогенная, таламо-париетальная) включает заднюю теменную кору и, по мнению Мезулама (Mesulam, 1981, 1985), выступает интегратором сенсорной информации из различных областей мозга, обеспечивая сенсорное принятие решения.
Обе системы (дельта и тета) взаимодействуют через корково-корковые (горизонтальные, внутри- и межполушарные) и корково-подкорковые (вертикальные) связи (Деглин, 1975; Павлова, 1980; Бианки, 1989; Шарова, 1990; Жаворонкова, 2007).
Особый интерес представляет начальная стадия подготовки СМИ в условиях последовательной маскировки, которая отражается в одновременном существовании контингентной негативной волны в центральной области (Е-волна) и позитивной волны в лобной области (CPV). Негативная волна на стадии ожидания, сформированная билатерально асимметричными негативными колебаниями в центральной области с доминированием правого (анализ целевого стимула) или левого полушария (анализ маскера), связана с активацией таламо-париетальной системы внимания, концентрирующей ресурсы внимания для оптимизации процесса восприятия. Тогда как, возникающее одновременно позитивное колебание отражает процесс притормаживания таламо-фронтальной системы, контролирующей механизмы произвольного внимания, необходимые для оптимизации сенсорных и моторных процессов, предотвращая появление «ложных» тревог в случае ОМ и повышающей пороги восприятия маскера, обеспечивая игнорирование маскера при ПМ. Взаимодействие этих двух процессов образует первый цикл регуляции СМИ. Сокращение МИ в обеих тестовых ситуациях приводило к постепенному сдвигу CNV в теменную область, уменьшению асимметрии за счет слияния двух фокусов, с последующим замещением Е-волны, развивающейся после завершения компонента Р3б, О-волной с доминированием сагиттального фронтального фокуса. Данная трансформация, вероятно, была связана с ухудшением процесса восприятия, особенно в условиях периферической маскировки (МИ 50 мс), требующего больше ресурсов внимания к теменным ассоциативным структурам (Е-волна) для идентификации целевого стимула, сливающегося с маскером. Процесс облегчается О-волной, возникающей в последействии целевого стимула, являющимся «кондиционирующим» по отношению к следующей комбинации маскера и целевого стимула. Воздействие маскера в этот период в условиях ПМ приводил к оптимизации СМИ, обеспечивая сокращение времени выполнения теста.
Стадия сенсорного анализа физических параметров стимула соответствовала развитию переднего фронта компонента N1, связанного с активацией нейронов-детекторов (Соколов и др, 1963; Picton et al., 1970; Picton et al., 1981; Naatanen, Picton, 1987; Escera et al., 2002), а заднего - с дополнительной активацией механизмов произвольного внимания для целевого стимула (Hillyard et al., 1973; Schwent, Hillyard, 1975; Desmedt, Robertson, 1977; Стрелец, Клюева, 1978). Возникновение N1 связано со смещением фокуса негативности из центрально-теменной области во фронтальную, что свидетельствует о перемещении очага доминирования из таламо-париетальной системы внимания (стадия ожидания) в таламо-фронтальную – (этап сенсорного принятия решения). Наличие механизма активного игнорирования маскера снижало амплитуду N1 за счет произвольного повышения порогов восприятия.
Стадия различения целевых стимулов сопровождалась развитием НР, в виде двух негативных ФМВ в правой верхнее-височной (доминирующей при разновероятной стимуляции) и левой нижнетеменной областях мозга, в местах пересечения дорсальных и вентральных путей (Hall, 2003, 2006) обработки слуховой информации. Генерация НР в височной области коры предполагает независимое хранение и извлечение информации о звуковом стимуле (Naatanen, 1990), во фронтальной – этот компонент связан с автоматическим переключением внимания, обеспечивая активацию таламо-фронтальной системы внимания (Alho, 1995). Данный процесс может быть связан с восстановлением СМИ (Айдаркин, 1999-2008) на интервале между однотипными (Gonsalves et al., 2007), а не последовательными стимулами (Naatanen et al., 1990). Трансформация исходной правосторонней асимметрии в левостороннюю на этапе анализа маскера при ПМ, вероятно, и обеспечивалет более быстрое распознавание целевого стимула за счет демпфирующей функции (Деглин, 1975) левого полушария, что приводит к активному игнорированию маскера. Маскер, приходящийся на период развития НР приводит к нарушению процесса восстановления СМИ (Айдаркин, 1999-2007), сформированной в достимульный период, который был прерван процессом анализа физических признаков стимула, что отражается в увеличении ВР.
Одновременное развитие компонентов Р2, связанного с модуляционными процессами внимания, определяющими значимость стимула в задаче обнаружения (Vaughan et al., 1980; Crowley, Colrain, 2004) и N2, основная роль которого состоит в активации следующего звена обработки стимула, связанной с его идентификацией и опознанием (Иваницкий, 1978), процессов сравнения образов пришедшего и хранящегося в памяти образа предыдущего стимула (Наатанен, 1998), а также с запуском моторной составляющей (Vaughan, Costa, 1968; Syndulko, 1969; Deecke et al., 1973; Hoffman, 1990), вероятно, свидетельствует о наличии механизма корково-гиппокампальной модуляции сенсорного процесса с вовлечением ресурсов памяти, что представляет второй цикл регуляции СМИ.
Процедура подготовки к реализации СМИ, связанная с процессами уменьшения степеней свободы, отражалась в развитии Р3, локализованного в лобно-центральной (Р3а, тета-колебания ССП) и теменной (Р3б, дельта-составляющая ССП) областях с ФМВ в сагиттальных отведениях (Polich, 2007). Лобная волна (Р3а), вероятно, притормаживала процессы (Donchin, 1980), связанные с произвольным контролем со стороны таламо-фронтальной системы. Теменная позитивность в виде Р3б, скорее всего, соответствовала процессам деактивации таламо-париетальной системы в третьем цикле регуляции СМИ. Влияние маскера на данном этапе при ОМ (300 мс) приводило к взаимодействию позитивных процессов игнорирования маскера (таламо-фронтальная система, формирующая Р3а и Р2 компоненты анализа маскера) и деактивации таламо-париетальной системы (Р3б), ухудшая различение целевых стимулов.
Таким образом, реализация СМИ связана с взаимодействием таламо-фронтальной, проявляющейся в симметричных процессах развития тета- колебаний ССП (N1, P2, N2, P3а), и таламо-париетальной, отражающихся в асимметричных процессах развития дельта- колебаний ССП с доминированием правого полушария на этапах развития CNV и НР, систем внимания. В условиях ОМ маскер воздействовал, в первую очередь, на тета-колебания ССП, снижая эффективность деятельности фронтальной системы внимания, сохраняя правосторонний характер асимметрии дельта-компонентов ССП. В условиях ПМ маскер воздействует на дельта-волну, формируя доминирующий очаг париетальных систем внимания в левом полушарии, обеспечивая его демпфирование, а затем возвращается к исходной правополушарной асимметрии на этапе НР, обеспечивая оптимальный анализ целевого стимула фронтальной системой, что приводит к сокращению ВР. Преобладание левой руки связано с интерференцией проекций соответствующих двигательных компонентов и доминирующих центров непроизвольного внимания в правом полушарии. Увеличение вероятности стимулов увеличивает роль непроизвольного внимания, приводя к усилению доминирования правого полушария как на уровне париетальных, так и фронтальных систем внимания. В условиях ПМ этот процесс дополняется демпфирующей функцией левого полушария на этапе ожидания маскера. Сокращение МИ приводит к смене достимульной (Е-волна) активации париетальных систем внимания, на послестимульную (О-волна), запускающую фронтальную систему.
ВЫВОДЫ:
- Скорость распознавания целевого стимула в условиях центральной маскировки повышается в условиях ПМ и снижается при ОМ по сравнению с контрольной серией бинарного различения.
- Преобладание ответа левой рукой наблюдается при ПМ. Увеличение вероятности целевого стимула приводит к появлению асимметрии при ОМ и усилению ее при ПМ.
- Процесс сенсомоторного различения в условиях последовательной слуховой маскировки сопровождается развитием медленных дельта волн, позитивно-негативной-позитивной при ПМ и негативно-позитивной при ОМ. В обеих ситуациях негативная составляющая представлена волной ожидания и негативностью рассогласования. Позитивная волна в случае ПМ представлена волной игнорирования (CPV и Р2), а в условиях ОМ – суперпозицией следовой позитивизации - Р3б и CPV, предшествующей маскеру.
- Функциональная правополушарная асимметрия возникает на этапе ожидания (CNV) и восстановления (НР) СМИ по типу реципрокных и демпфирующих (особенно для маскера) отношений в виде негативных дельта колебаний с двумя асимметричными фокусами.
- Сокращение МИ приводит к воздействию маскера при ОМ на тета-колебания ССП, определяющие основные этапы анализа стимула, лежащие в основе развития N1, P2, N2 и P3а компонентов ССП и дельта модулирующую волну, сформированную CNV-НР-Р3б компонентами ССП, а также к смене достимульной (Е-волна), активирующей париетальную систему внимания, на фронтальную (О-волну), выраженную в постстимульный период.
Список основных работ, опубликованных по теме диссертации в изданиях, рекомендованные ВАК рф
- Патент 2341183 от 03.04.2007 РФ, МПК Ф61И 5/00 (2006.01). Способ оценки уровня внимания учащегося при компьютерном тестировании / Е.К. Айдаркин, Д.Н. Щербина, Н.В. Пахомов, М.А. Павловская, Е.С. Айдаркина.- № 2007112370/14; заявл. 03.04.2007; Опуб. 20.12.2008; Приоритет 03.04.2007. – 0.042 п.л., личный вклад 10%.
- Айдаркин Е.К., Павловская М.А. Исследование нейрофизиологических механизмов непроизвольного внимания в условиях центральной маскировки слухового стимула // Валеология, №1, 2007, C. 65-74. – 0.21 п.л., личный вклад 50%.
- Бабенко В.В., Кульба С.Н., Павловская М.А. Временная суммация амплитудно-модулированного шума в слуховой системе человека // Психологический журнал, №1, 2007, C. 100-106. – 0.098 п.л., личный вклад 30%.
- Айдаркин Е.К., Павловская М.А. Исследование нейрофизиологических механизмов взаимодействия произвольного и непроизвольного внимания в условиях последовательной слуховой маскировки // Валеология, №3, 2007, C. 103-113. – 0.231 п.л., личный вклад 50%.
- Айдаркин Е.К., Павловская М.А. Исследование ритмических составляющих связанных с событием потенциалов в условиях слуховой последовательной маскировки» // Валеология, №4, 2008, C. 63-79. – 0.29 п.л., личный вклад 40%.
- Айдаркин Е.К., Павловская М.А., Одинцова Н.А. Нейрофизиологические закономерности последовательной маскировки в зрительной и слуховой системах // Валеология, №4, 2008, C. 63-81. – 0.168 п.л., личный вклад 40%.
Список работ, опубликованных по теме диссертации
- Павловская М.А. Сравнительная эффективность on - и off - механизмов на временной и частотной шкалах в слуховой системе человека // Сб. материалов VII Всероссийской медико-биологической конференция молодых исследователей «Человек и его здоровье». Тезисы. - Санкт-Петербург, 2004. с - 206-207. – 0,084 п.л., личный вклад 100%.
- Павловская М.А. Зависимость порога обнаружения амплитудной модуляции шума от частоты и числа периодов модуляции // Физиология и медицина: Сб.материалов Всерос.конф.молодых исследователей; 14-16 апр. 2005г., г.С.-Петербург.- СПб.: [Б.и.],2005.-С.86.- (Вестн.молодых ученых. Сер.Науки о жизни. 2005.Прил.). – 0,042 п.л., личный вклад 100%.
- Павловская М.А. Исследование слуховых механизмов временной интеграции амплитудно-модулированного шума. Сб. материалов I ежегодной конференции студентов и аспирантов базовых кафедр ЮНЦ РАН. Тезисы.; Ростов на дону: Изд-во ЮНЦ РАН, 2005. С. 334-335. – 0,084 п.л., личный вклад 100%.
- Павловская М.А. Исследование закономерностей группирования звуковых сигналов на частотной и временной шкалах в слуховой системе человека // Сб. материалов III Межрегиональной научной конференции студентов, аспирантов и молодых ученых «Молодежь XXI века – будущее Российской науки» / РГУ секция «Психология»; Тезисы. - Ростов - на –Дону, ЦВВР, 2005 г. С. 267-268. – 0,084 п.л., личный вклад 100%.
- Бабенко В.В., Кульба С.Н., Павловская М.А. Закономерности восприятия амплитудной модуляции шума в слуховой системе человека // Мат. 14-й Международной конференции по нейрокибернетике «Проблемы нейрокибернетики». Том 1. Ростов-на-Дону. Издательство ООО «ЦВВР». 2005. с. 204-206. – 0.042 п.л., личный вклад 30%.
- Павловская М.А. Исследование нейрофизиологических механизмов непроизвольного внимания человека при изменении соотношения физических параметров звукового сигнала // Мат. IX Всероссийской медико-биологической конференция молодых исследователей «Человек и его здоровье». Тезисы. - Санкт-Петербург, 2006. - С. 252 - 253. – 0.084 п.л., личный вклад 100%.
- Павловская М.А. Влияние физических параметров стимула на конфигурацию слуховых вызванных потенциалов мозга человека // Мат. II ежегодной научной конференции студентов и аспирантов базовых кафедр ЮНЦ РАН. Тезисы.- Ростов-на-Дону: Изд-во ЮНЦ РАН, 2006. – С. 35 - 36. – 0.084 п.л., личный вклад 100%.
- Старостин А.Н., Павловская М.А. Исследование влияния динамики межстимульного интервала на функциональное состояние человека // Материалы XII Международного симпозиума "Эколого-физиологические проблемы адаптации" (Москва, 30-31 января 2007).- М., 2007.- С. 420-421. – 0.042 п.л., личный вклад 50%.
- Павловская М.А. Исследование нейрофизиологических механизмов последовательной маскировки // Мат ХХ съезда Физиологического общества им. И.П. Павлова. Тезисы докладов. - М.: Издательский дом «Русский врач», 2007.- С. 366-367. – 0.084 п.л., личный вклад 100%.
- Павловская М.А. Исследование взаимодействия механизмов произвольного и непроизвольного внимания в условиях последовательной звуковой маскировки [Текст] // Материалы международной конференции студентов, аспирантов и молодых ученых "Ломоносов-2007". - Москва, 2007. - C.52. – 0.042 п.л., личный вклад 100%.
- Павловская М.А. Изучение механизмов непроизвольного внимания в условиях последовательной маскировки звукового сигнала // Мат. IX Всероссийской медико-биологической конференция молодых исследователей «Фундаментальная наука и клиническая медицина». Тезисы. - Санкт-Петербург, 2007. –С.332-333. – 0.084 п.л., личный вклад 100%.
- Айдаркин Е.К., Павловская М.А. Исследование нейрофизиологических механизмов непроизвольного внимания в условиях последовательной маскировки по параметрам ССП // Мат. III Конгресса «Нейронаука для медицины и психологии». Тезисы.- Москва, 2007.- С. 33. – 0.021 п.л., личный вклад 50%.
- Aydarkin E.K., Pavlovskaya M.A. The neurophysiologic mechanisms of the involuntary attention in condition successive auditory masking by ERP`s components // Math. III-th Neuroscience for medicine and Psychophysiology Congress.- M.: МАКС Пресс, 2007.- P.- 34. – 0.021 п.л., личный вклад 50%.
- Павловская М.А. Время в условиях последовательной маскировки [Текст] // Материалы XV Международной конференции студентов, аспирантов и молодых ученых "Ломоносов-2008"; секция «Биология». - М.: МАКС Пресс, 2008. - C.173-174. – 0.084 п.л., личный вклад 100%.
- Павловская М.А. Нейрофизиологические механизмы формирования сенсомоторной интеграции в условиях последовательной звуковой маскировки // Мат. XI Всероссийской медико-биологической конференция молодых исследователей «Фундаментальная наука и клиническая медицина». Тезисы. - Санкт-Петербург, 2008. –С.276-277. – 0.084 п.л., личный вклад 100%.
- Павловская М.А. Психофизиологические механизмы центральной последовательной слуховой маскировки // Мат. III-го Международного междисциплинарного Конгресса «Нейронаука для медицины и психологии».- M.: МАКС Пресс, 2008.- С. 222-223. – 0.084 п.л., личный вклад 100%.
- Pavlovskaya M.A. Psychophysiological mechanisms of central successive auditory masking // Math. III-th Neuroscience for medicine and Psychophysiology Congress.- M.: МАКС Пресс, 2008.- P. 223-224. – 0.084 п.л., личный вклад 100%.
- Pavlovskaya M.A. Successive auditory masking and sensomotor integration // In. J. Psychophysiology. New York, NY : Cambridge University Press. V. 69/3, 2008. P. 305-306. – 0.084 п.л., личный вклад 100%.
- Айдаркин Е.К., Лепешкова Н.В., Павловская М.А. Использование влияния маскировочного стимула для коррекции эффективности деятельности оператора // Мат. II Всероссийской научно-практической конференции «Функциональное состояние и здоровье человека» - Ростов-на-Дону, 2008.- С.65-67. – 0.063 п.л., личный вклад 50%.
- Павловская М.А. Динамика механизмов локальной активации мозга в условиях последовательной слуховой маскировки // Мат. II Всероссийской научно-практической конференции «Функциональное состояние и здоровье человека» - Ростов-на-Дону, 2008.- С. 93-96. – 0.168 п.л., личный вклад 100%.
- Павловская М.А. Механизм корково-подкорковой регуляция различных этапов восприятия в условиях последовательной звуковой маскировки // Мат. IV Межрегиональной научной конференции студентов, аспирантов и молодых ученых «Молодежь XXI века – будущее Российской науки» / ЮФУ секция «Биология»; Ростов - на –Дону, 2008 г. С. 215. – 0,042 п.л., личный вклад 100%.
- Павловская М.А., Ан А.С. Исследование механизмов чередования прямой и обратной последовательной слуховой маскировки // Мат. XII Научной конференции молодых ученных по физиологии ВНД и нейрофизиологии.- Москва, ИВНД и НФ, 2008. С.- 53. – 0.038 п.л., личный вклад 90%.
- Павловская М.А., Одинцова Н.А. Нейрофизиологические закономерности последовательной маскировки в зрительной и слуховой системах // Мат. XII Научной конференции молодых ученных по физиологии ВНД и нейрофизиологии.- Москва, ИВНД и НФ, 2008. С.- 52. – 0.038 п.л., личный вклад 90%.
- Айдаркин Е.К., Павловская М.А. Роль функциональной межполушарной асимметрии в формировании сенсомоторной интеграции в условиях зрительной и слуховой последовательной маскировки // Мат. Научной конференции «Актуальные вопросы функциональной межполушарной асимметрии и нейропластичности» (Материалы Всероссийской конференции с международным участием). – М.: Научный мир, 2008. С. 9-14. – 0.125 п.л., личный вклад 50%.
Список научных сокращений
ВР – время реакции
ЛП – латентный период
МИ – маскировочный интервал
ОМ – обратная маскировка
ПМ – прямая маскировка
ПСМР – простая сенсомоторная реакция
СМИ – сенсомоторная интеграция
СМР – сенсомоторная реакция
ССМР – сложная сенсомоторная реакция
ССП – связанный с событием потенциал
ФМА – функциональная межполушарная асимметрия
ФМВ – фокус максимальной выраженности
ЭЭГ – электроэнцефалограмма