

Молекулярно-генетическое и экологическое разнообразие рода chionomys на кавказе
На правах рукописи
Сижажева Аминат Мухамедовна
МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКОЕ
И ЭКОЛОГИЧЕСКОЕ РАЗНООБРАЗИЕ
РОДА CHIONOMYS НА КАВКАЗЕ
03.02.14 – Биологические ресурсы
АВТОРЕФЕРАТ
Диссертации на соискание ученой степени
кандидата биологических наук
Владикавказ-2012
Работа выполнена в ФГБОУ ВПО «Кабардино-Балкарский государственный университет им. Х.М. Бербекова» на кафедре общей биологии, экологи и природопользования
Научные руководители: Заслуженный деятель науки КБР,
доктор биологических наук, профессор
Дзуев Руслан Исмагилович
кандидат биологических наук
Банникова Анна Андреевна
Официальные оппоненты: доктор биологических наук, профессор
Чопикашвили Лидия Васильевна
доктор биологических наук, профессор
Шахмурзов Мухамед Музакирович
Ведущая организация: ФГБОУ ВПО «Ставропольский
государственный университет»
Защита диссертации состоится 30 апреля 2012 г.. в 10 ч. на заседании диссертационного совета Д 220.023.04 при ФГБОУ ВПО «Горский государственный аграрный университет» по адресу: 362040, РСО-Алания, г. Владикавказ, ул. Кирова, 37, Горский ГАУ, зал заседаний диссертационного совета. Тел./факс: (8672) 53-99-26; E-mail: [email protected]
С диссертацией можно ознакомиться в библиотеке ФГБОУ ВПО «Горский государственный аграрный университет», с авторефератом на официальном сайте www.gorskigau.ru
Текст объявления о защите диссертации и автореферат диссертации отправлены в Минобрнауки РФ по адресам: vak2.ed.gov.ru и [email protected] 29 марта 2012 г.
Автореферат диссертации разослан 29 марта 2012 г.
Ученый секретарь диссертационного совета,
кандидат биологических наук, доцент С.А. Гревцова
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы. Многообразие ландшафтных условий Кавказа, его животное население и растительный мир издавна вызывают живой интерес как отечественных, так и зарубежных исследователей. Более того, отдельные группы млекопитающих и растений региона служили объектом изучения и предметом дискуссий. Очевидно, это связано с тем, что внутривидовая изменчивость параметров вида (морфологическая, цитогенетическая, зоогеографическая и т.д.), весьма сложная, что, по мнению Р.И. Дзуева (1995), обусловлено длительной самобытной историей формирования, пестротой ландшафтного покрова и трехмерным положением ареала в пространстве.
На протяжении последних двух-трех десятилетий в изучении териофауны Кавказа видное место занимают различные аспекты географической, популяционной, цитогенетической изменчивости, а в последние годы и молекулярно-генетических параметров вида, в связи с биологическим эффектом взаимодействия горных и равнинных экосистем.
Стремительное развитие и усовершенствование в последние десятилетия молекулярно-генетических методов филогенетики и систематики, привели к заметному увеличению числа работ, в которых рассматриваются родственные отношения самых разных групп животных, так как появилась возможность тестировать трудные или неразделимые на морфологическом уровне макро- и микроэволюционные гипотезы. Пристальное внимание при этом уделяется сейчас вопросам как внутривидовой, так и межвидовой генетической изменчивости и их таксономических категорий (Банникова, 2004; Абрамсон, 2007; Банникова и др., 2010; Lebedev et al., 2007; Castiglia et al., 2009). Несомненно, на млекопитающих это отразилось в наибольшей степени.
Исследования внутривидовой генетической изменчивости позволяют не только решать внутривидовую филогению и систематику, но и получить объективные данные о центре возникновения видов и формирования ареалов, что в свою очередь приводит к лучшему пониманию геологической истории регионов, а также биоресурсного потенциала исследуемой территории.
Наиболее полные данные о филогеографической структуре видов снежных полевок в настоящее время имеются для Chionomys nivalis Martins, 1842 (Castiglia et.al., 2009). Таких материалов по Chionomys gud Satunin, 1909 и Chionomys roberti Thomas 1906 в доступной научной литературе найти не удалось. Между тем хорошо известно, что без аналогичных данных по роду Chionomys Кавказа фактически невозможно решить дискуссионные вопросы филогении и о систематико-таксономическом положении представителей этих групп полевок. В связи, с чем представляется весьма актуальным исследование существующего состава рода Chionomys на Кавказе, что явилось причиной предпринятых впервые нами исследовательских работ по анализу митохондриального ДНК (cytb).
Цель работы – исследование генетической изменчивости, филогенетических отношений и дивергенции снежных полевок на Кавказе на основе анализа митохондриальной ДНК, цитогенетических параметров и составление на этой основе кадастровых карт ареалов.
Для достижения этой цели были поставлены следующие задачи:
- определить первичные нуклеотидные последовательности (далее – последовательности) и нуклеотидный состав гена cytb исследуемых представителей снежных полевок;
- получить гаплотипы митохондриального гена b Ch. gud, Ch. roberti из разных географических точек на Кавказе и на этой основе сделать заключение о внутривидовой генетической дифференциации и филогенетическом положении изученных видов в составе рода по митохондриальным и ядерным генам;
- изучить внутривидовую молекулярную дифференциацию Ch. nivalis на основе, главным образом, данных Генбанка по цитохрому b, а также положение вида в системе рода по оригинальным данным;
- выяснить, как соотносятся уровни внутри- и межвидовой дивергенции у исследуемых представителей различных таксонов снежных полевок;
- провести сравнительный анализ филогении рода Chionomys Кавказа, полученных с использованием различных митохондриальных маркеров, цитогенетических и морфологических данных;
- составить кадастровые карты ареалов видов Chionomys на Кавказе.
Научная новизна работы. Впервые получены гаплотипы митохондриального гена b для Ch. gud и Ch. roberti из разных географических точек Кавказа. На основе этих данных выявлена внутривидовая генетическая дифференциация представителей рода Chionomys. С помощью молекулярно-филогенетических методов установлены эволюционные взаимосвязи между видами рода Chionomys. Выдвинута гипотеза о более раннем отделении Ch. nivalis от общего предкового ствола на основании изученных видовых параметров.
На молекулярно-генетически датированном материале составлены кадастровые карты распространения видов рода Chionomys. Снежные полевки Кавказа предложены как модельная группа для исследования внутри- и межвидовой изменчивости в горах.
Теоретическая и практическая значимость работы. Исследование носит теоретический характер, оно будет содействовать развитию систематики и филогении всего рода Chionomys и в целом подсемейства Arvicolinae Евразии, выявлению общебиологических закономерностей изменчивости и микроэволюции в горах, совершествованию теоретических основ охраны уникального генофонда фауны Кавказа.
Представленные генетические маркеры cytb могут быть применены для идентификации видов и внутривидовых форм снежных полевок в таксономических, эволюционных и природоохранных целях, а также использованы в Международном Генбанке по штрих кодированию серых полевок на основе ДНК.
Результаты работы могут применяться в практических мероприятиях, использующих данные таксономических исследований и сведения по инвентаризации фауны (при составлении кадастров, разработке стратегии и тактики охраны цено- и генофонда Кавказа, проведении экологического мониторинга и т.п.). Карты распространения, составленные отдельно по видам, могут быть использованы при обосновании необходимости совершенствования сети охраняемых территорий региона.
Апробация работы. Материалы диссертационной работы докладывались и обсуждались на ежегодных научно-практических семинарах кафедры общей биологии, экологии и природопользования и научно-исследовательской лаборатории горной экологии КБГУ (Нальчик, 2007, 2008, 2009, 2010, 2011), XI Международной конференции «Биологическое разнообразие Кавказа» (Магас, 2009); Всероссийской научной конференции «Целостность вида у млекопитающих: изолирующие барьеры и гибридизация» (Петергоф, 2010); XII Международной конференции «Биологическое разнообразие Кавказа» (Махачкала, 2010); XIII Международной конференции «Биологическое разнообразие Кавказа» (Грозный, 2011); Международной научной конференции студентов, аспирантов и молодых ученых «Перспектива-2011» (Нальчик, 2011).
На защиту выносятся следующие научные положения и основные результаты исследований:
- результаты, полученные при анализе митохондриального гена b для Ch. gud и Ch. roberti из разных географических точек Кавказа;
- данные о внутривидовой генетической дифференциации по митохондриальному гену цитохрома b снежных полевок Кавказа;
- показатели, характеризующие филогенетические отношения видов рода Chionomys;
- цитогенетические показатели (хромосомные наборы) снежных полевок;
- особенности распространения, экологии и биологии Ch. nivalis, Ch. gud, Ch. roberti.
Публикации результатов исследований. По материалам диссертации опубликовано 9 научных работ, в том числе одна монография и 3 работы в рекомендуемых ВАК РФ изданиях.
Объем и структура диссертации. Диссертация изложена на 150 страницах машинописного текста и состоит из введения, обзора литературы, материала и методов исследований, результатов исследований и их обсуждения, выводов, библиографии. Работа иллюстрирована 9 таблицами, 20 рисунками, в том числе 6 фотографиями. Список литературы включает 218 наименований, в том числе 76 иностранных источников.
Благодарности. Я благодарна моим научным руководителям: Руслану Исмагиловичу Дзуеву, чьи идеи и научные планы стали основой этой работы, многие исследования проводились и проводятся под его непосредственным руководством, без его постоянного внимания работа не была бы завершена; и Анне Андреевне Банниковой, которая способствовала освоению мною молекулярно-генетического метода исследований, а также анализу и интерпретации полученных нами материалов. Кроме того, часть материалов, использованных в диссертации, она любезно предоставила нам для дальнейших анализов.
Искренне признательна за поддержку, советы и всестороннюю помощь при выполнении работы сотрудникам кафедры общей биологии, экологии и природопользования КБГУ к.б.н., доцентам: В.Н. Кануковой, М.М. Хуламхановой, а также моим коллегам, сотрудникам научно-исследовательской лаборатории горной экологии КБГУ: М.А. Хашкуловой, А.Х. Чеченовой, С.М. Казиевой, Р.В. Курашиновой, коллективу кабинета молекулярной зоологии при кафедре зоологии позвоночных МГУ.
Выражаю глубокую признательность к.б.н., с.н.с. ЗИН РАН Ф.Н. Голенищеву; к.б.н., с.н.с. ЗИН РАН В.Г. Маликову за предоставленные образцы Сh. gud и Ch. roberti; к.б.н., с.н.с. В.С. Лебедеву за помощь во время обработки коллекционного материала ЗМ МГУ; к.б.н., доценту кафедры ботаники АГУ Л.В. Хасановой, с которой проводила полевые исследования во время научных экспедиций по Кавказу. Искреннюю благодарность выражаю к.б.н., с.н.с. Ин-та зоологии АН Азербайджана Г.Н. Кулиеву, который предоставил для исследования микропрепараты хромосом Chionomys из Закавказья.
Я очень благодарна моим родителям, сестре, а также моим друзьям за безграничное терпение и поддержку. Также хочется поблагодарить водителя Кармокова Ч.Х., без которого не были бы возможны экспедиционные выезды по сбору материала.
Я признательна руководству КБГУ за предоставленную мне возможность заниматься научной работой как в условиях научно-исследовательской лаборатории горной экологии, так и в медико-биологическом центре.
ОСНОВНОЕ СОДЕРЖАНИЕ РАБОТЫ
Глава i. Филогения, систематика и экология снежных полевок
Глава содержит литературные сведения, систематизированные по нескольким направлениям: I – Вклад зоологических исследований в изучение териофауны Кавказа, охватывающий конец XVIII – начало XIX века до наших дней, II – Краткая история описания представителей рода Chionomys, (география, известные палеонтологические данные, общие проблемы таксономии и изменчивости), III – Морфологическая и морфометрическая характеристика и положение снежных полевок в системе Arvicolinae, IV – Экология и морфофизиология представителей рода Chionomys на Кавказе, V – Молекулярно-генетическая изученность снежных полевок.
Глава ii. Материалы и методы исследования
В основу настоящей работы положены материалы, собранные соискателем в различных районах Кавказа с 2004 по 2011 гг., составляющие 487 экз. тушек, шкурок, черепов и полных скелетов, в том числе: Ch. gud – 238 экз., Ch. roberti – 196 экз. и Ch. nivalis – 53 экз. За время исследования отработано более 1500 ловушко-суток.
Кроме того, в работе были использованы коллекционные материалы ряда зоомузеев РФ: Кабардино-Балкарского госуниверситета (сборы сотрудников кафедры общей биологии, экологии и природопользования, научно-исследовательской лаборатории горной экологии с 1967-го по 2011 г.), Московского госуниверситета (Ch. gud – 63 тушки и 57 черепов; соответственно Ch. roberti – 51и 58 и Ch. nivalis 62 и 60), Зоологического института РАН (Ch. nivalis 32 и Ch. roberti 35 экз.).
Учеты численности исследуемых видов снежных полевок мы проводили по общепринятой методике А.П. Кузякина (1962).
Нам также была предоставлена возможность изучить уникальные материалы (коллекции) с.н.с. кафедры зоологии позвоночных животных МГУ А.А. Банниковой по митохондриальной и ядерной ДНК представителей подсемейства полевочьи (Arvicolinae) Евразии.
Значительное внимание в нашей работе уделялось полевым исследованиям снежных полевок Кавказа. Обследование проводилось в основном по методике, предложенной Г.А. Новиковым (1953) и Р.И. Дзуевым и др. (2000).
Пробы для анализа митохондриального и ядерного ДНК, фиксировали в 96о этиловом спирте (Булатова и др., 2009).
Всего для анализа митохондриального и ядерного ДНК нами были отловлены и исследованы 42 особи трех видов снежных полевок, в том числе: Ch.roberti – 12, Ch.gud – 25, Ch. nivalis – 5.
Геномную ДНК выделяли из фиксированных этанолом мышц, печени, почек и семенников стандартным методом фенол-хлороформной депротеинизации после обработки гомогената тканей протеиназой К (Sambrook et al., 1989).
Реконструкция филогенетических деревьев была выполнена с помощью нескольких широко используемых алгоритмов. Филогенетический анализ по методам максимального правдоподобия (maximum likelihood, ML), ближайшего связывания (neighbor-joining, NJ) и максимальной экономии (maximum parsimony, MP) выполнен в программах пакета MEGA 4.0.0.4083. Реконструкции ML деревьев выполнена с помощью программы пакета Treefinder, version October 2008 (Jobb, 2008). Реконструкции на основе метода ML предварялась определением оптимальной модели эволюции последовательности с помощью программы Modeltest версия 3.7 (Posada, Crandall, 1998) на основе критерия BIC.
Для определения степени различий между типами мтДНК и расчетах генетических дистанций использовали некорректированную p-дистанцию, – наблюдаемую пропорцию нуклеотидов, неодинаковых у двух последовательностей и выраженную в частях от единицы, либо в процентах.
Анализ внутривидового генетического разнообразия по результатам секвенирования cytb включал оценку гаплотипического (H) и нуклеотидного () разнообразия, которое проводили с помощью программы Arlequin 2.000 (Schneider et al., 2000), и построение сетевых деревьев в программе Network 4.5.0.0 (Bandel et al., 1999).
Хромосомные препараты приготовлены из костного мозга по общепринятой методике (Ford, Hamerton, 1956; Орлов, Булатова, 1983; Дзуев, 1998). Изучение структурного гетерохроматина проводили по методике, предложенной А. Самнером (Sammner, 1972) с некоторой модификацией, предложенной Р.И. Дзуевым (1998) для мелких млекопитающих.
Сознавая важность создания кадастровых карт, а также их значение для понимания хромосомной и молекулярно-генетической изменчивости, систематики и филогении рода Chionomys на Кавказе, мы нанесли на схематическую карту все известные нам места добычи трех видов снежных полевок на территории исследуемого региона, привязав их к названиям урочищ и населенных пунктов. Таких точек на картах распространения снежных полевок на Кавказе насчитывается 122.
При составлении карт мы руководствовались методическими рекомендациями Н.В. Тупиковой (1969), Х.Х. Шхашамишева (1992).
Глава III. Кариотипическая характеристика снежных полевок (Chionomys) Кавказа
Кариология млекопитающих – одно из бурно развивающихся направлений териологических исследований. Библиография работ в этой области в исследуемом регионе к настоящему времени насчитывает более 250 названий, причем подавляющее большинство их относится к последним 15–25 годам, и выполнено в основном местными териологами – цитогенетиками.
Бурное развитие кариологии млекопитающих объясняется, во-первых, методическим прогрессом, позволяющим получать надежные фактические данные, и, во-вторых, эффективностью этих данных в решении проблем териологии и общей биологии в целом.
В нашем случае, поскольку у представителей рода Chionomys хромосомные наборы описаны всего лишь по небольшому числу особей (исключение составляет Ch. gud), на наш взгляд, встает необходимость увеличения как количества исследованных особей видов рода, так и количества изученных популяций, особенно значительно изолированных от видовых ареалов. Это необходимо для определения эволюционной роли хромосомного полиморфизма, природы селективных преимуществ, которые дают отдельной популяции и виду в целом меж- и внутрипопуляционная вариабельность числа и морфологии хромосом.
Вышеизложенное положение определило программу последовательного изучения кариотипа трех видов снежных полевок (Ch. nivalis, Ch, gud, Ch. roberti) на Кавказе.
3.1. Хромосомный набор снежной полевки (Ch. nivalis)
Как видно из рис. 1, диплоидный набор снежной полевки содержит 54 хромосомы. Число плеч аутосом (NFa) равно 52, а основное число плеч хромосом составляет 56. Аутосомы представлены акроцентриками. За исключением первой пары акроцентрических аутосом, которая несколько больше по размерам, остальные составляют по размерам постепенно и равномерно убывающий ряд.

Рис. 1. Кариотип снежной полевки – Chionomys nivalis.
2n = 54, NFa = 52
Половые хромосомы заметно гетероморфные, легко идентифицируются. По морфологии Х-хромосома относится к субметацентрическому типу и по величине она приравнивается к первой паре аутосом. У-хромосома представлена мелким акроцентриком. Хромосомный набор Ch. nivalis на протяжении видового ареала (от Западной Европы до Кавказа включительно) не обнаруживает оптически выявляемого полиморфизма как по количеству, морфологии аутосом и гетерохромосом, так и дифференциально окрашенных метафазных хромосом (Todorovic et al., 1971; Pechev et all., 1979; Кулиев, 1979; Guardia et al., 1981; Яценко, 1982 и наши данные).
3.2. Кариотип полевки Роберта (Ch. roberti)
Как видно из рис. 2, в диплоидном наборе у всех исследованных полевок содержится 54 хромосомы, NFa = 54, NF = 57 у самцов и 58 у самок. Аутосомы представлены акроцентрическими элементами за исключением 26-пары, которая имеет двуплечую структуру. Среди акроцентрических аутосом первая пара самая крупная, остальные так резко не отличаются и образуют отдельные группы уравненных по размеру хромосом с постепенным уменьшением их величины.
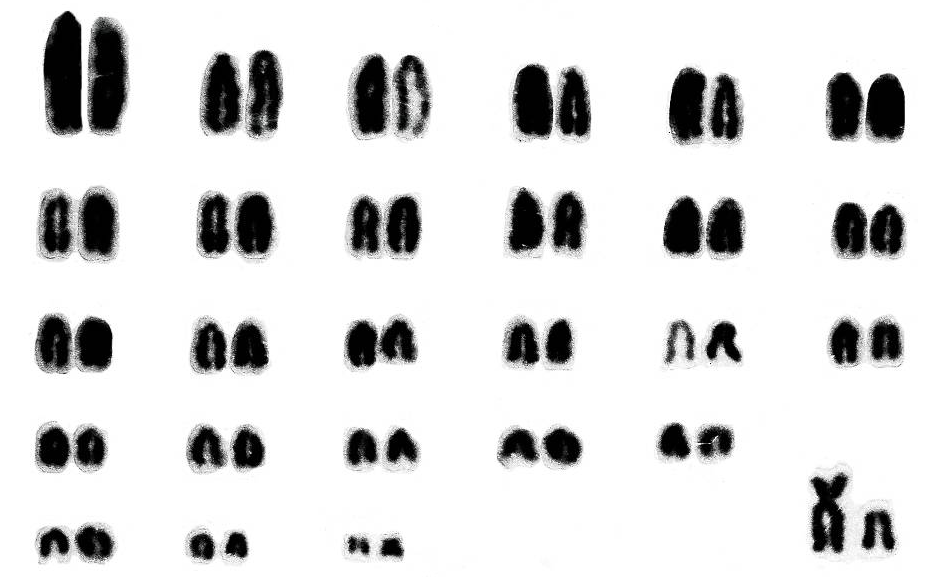
Рис. 2. Кариотип полевки Роберта – Chionomys roberti.
2n = 54, NFa = 54
Половые хромосомы идентифицируются довольно четко: Х-хромосома субметацентрическая, приравниваемая по размерам к первой паре самых крупных аутосом; Y-хромосома – акроцентрическая, занимающая по величине промежуточное положение между 23–24-й парами аутосом.
Кариотип этого вида также исследован методом дифференциальной окраски с использованием G-полос и С-окраски (Саблина и др., 1988; наши данные).
Как видно из изложенного материала и литературных сведений, хромосомный набор Ch. roberti не обнаруживает оптически видимого полиморфизма как по морфологии хромосом, строению G-полос и С-окраски, так и по количеству хромосом и основному числу плеч аутосом и гетерохромосомного комплекса.
3.3. Кариотип гудаурской полевки (Ch. gud)
Как видно из рис. 3, кариотип в диплоидном наборе содержит 54 хромосомы, число плеч аутосом (NFa) равно 54, а основное число плеч хромосом у самцов составляет 57, у самок – 58.
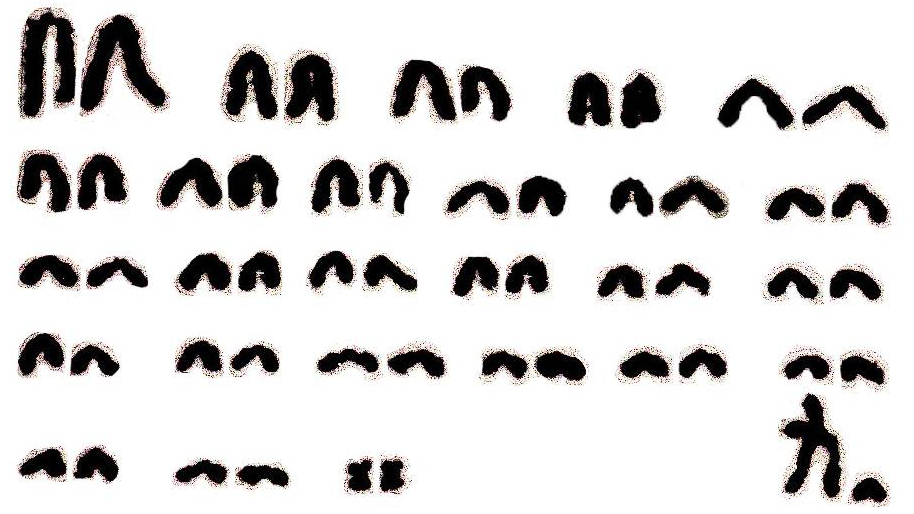
Рис. 3. Кариотип гудаурской полевки – Chionomys gud.
2n = 54, NFa = 54
Набор аутосом состоит из 25 пар акроцентрических элементов и одной пары метацентриков. Акроцентрические аутосомы по размерам, как и у выше приведенных представителей этого рода, заметно гетероморфны: одна пара занимает по величине первое место в наборе и выделяется от остальных акроцентрических элементов по размерам, а другие не отличаются резко и составляют плавно убывающий ряд. Единственная пара метацентрических аутосом по величине занимает последнее место в кариограмме.
Гетерохромосомы легко идентифицируются. По морфологии Х-хромосома относится к субметацентрическому типу и по величине приравнивается к первой паре аутосом. Y-хромосома представлена мелким акроцентриком, по размерам занимает промежуточное положение между 22–23-й парами аутосом.
Хромосомный набор гудаурской полевки на всем протяжении ареала стабильный как по числу хромосом в наборе, так и по морфологии аутосом и гетерохромосом. Таким образом, кариологические исследования представителей рода Chionomys Кавказа, еще раз подтвердили их цитогенетическую видоспецифичность.
Особенно важно это для снежной (Ch. nivalis) и гудаурской (Ch. gud) полевок – видов, систематическое положение которых остается дискуссионным до настоящего времени. Полученные нами и коллегами данные по кариотипам этих двух морфологически фактически не различимых представителей рода Chionomys свидетельствуют о том, что они четко отличаются по этому параметру, что еще раз подтверждае т их видовую самостоятельность.
Глава IV. Молекулярно-генетическая характеристика биологического разнообразия рода Chionomys Кавказа
Использование молекулярных маркеров в систематике и экологии сегодня стало неотъемлемой частью современных зоологических исследований. В таксономии молекулярный подход привел к пересмотру филогении и систем классификаций многих групп животных и растений, в т.ч. млекопитающих, где привлечение данных по секвенированию митохондриальных и ядерных генов перевернуло большую часть прежних представлений о филогении отрядов и надотрядных группировок (Stanhope et al., 1998; Murphy et al., 2001; Nikaido et al., 2003 a,b). В экологии развитие молекулярно-генетических исследований обусловило качественный скачок в популяционной биологии, в основе которой теперь прочно стоит генетика популяций. Применению молекулярных маркеров и проблемам их использования и интерпретации получаемых результатов посвящено немало обзорных статей (Гречко, 2002; Банникова, 2004; Edwards et al., 2005; Delsuc et al., 2005 и др.).
4.1. Характеристика последовательностей и нуклеотидный состав у исследуемых видов рода Chionomys
Конечная матрица, используемая в анализе, включала 68 последовательностей трех распознаваемых видов Chionomys, 26 последовательностей было взято из Генбанка. Конечное выравнивание составило 1140 п.н. Неполные последовательности длиной более 1000 п.н. (21 последовательность, из которых 18 из Генбанка), а также три коротких сиквенса длиной 573 п.н. (Ch. rob 1–10),
553 п.н. (ZIN73189) и 468 п.н. (Ch. gud A) также были использованы в анализе.
Митохондриальная ДНК (cytb) и ядерные гены (10-й экзон GHR и 11-й экзон BRCA1). Для выяснения филогенетических взаимоотношений видов снежных полевок и их внутривидовой генетической дивергенции на основе митохондриального ДНК и ядерных генов были построены деревья с использованием трех разных филогенетических алгоритмов (NJ, MP, ML).
На дереве ядерных генов, которое построено методом максимального правдоподобия (ML), указаны поддержки, полученные в парсимониальном (MP) и дистанционном (NJ) анализах.
На ML-дереве митохондриального гена cytb Ch. gud и Ch. roberti группируются вместе с бутстрэп-поддержкой (bv>50%) (рис. 4).
На всех деревьях в составе рода Chionomys выявляются 3 основные группы гаплотипов, которые имеют высокие поддержки бутстрэпа и отстоят друг от друга на существенные генетические дистанции. Эти группировки соответствуют трем видам: Ch. gud, Ch. roberti и Ch. nivalis.

Рис. 4. Филогенетическое ML-дерево видов рода Chionomys по результатам анализа
1140 п.н. митохондриального гена cytb. Числа вблизи ветвей – показатели поддержек
(>50%) в бутстрэп-анализе (1000 реплик)
На рис. 5 приведено суммарное дерево, полученное в результате анализа комбинированной последовательности из двух ядерных генов.
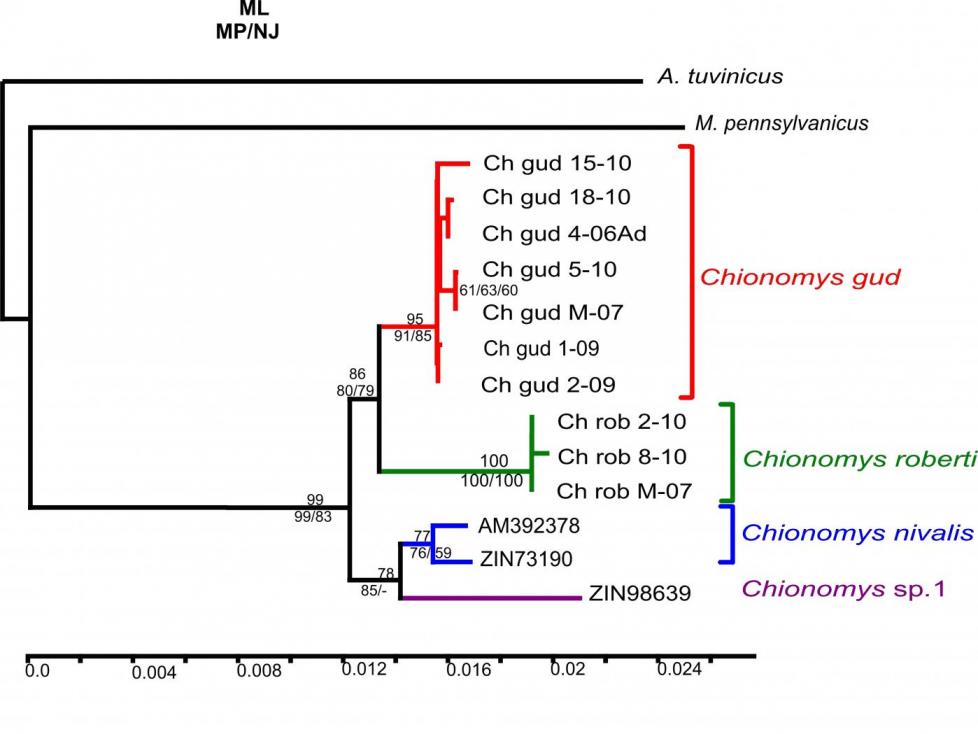
Рис. 5. Филогенетическое ML-дерево рода Chionomys по результатам анализа комбинированной последовательности 10-го экзона GHR1 и 11-го экзона BRCA1. Бутстрэп-поддержки (>50 %) в ML, NJ и MP анализах приведены вблизи узлов в последовательности
ML/MP/NJ соответственно (1000 реплик)
Филогенетический анализ комбинированной последовательности ядерных экзонов GHR и BRCA1 подтвердил наличие в анализируемой выборке образцов четырех дивергированных клад, три из которых соответствуют распознаваемым ранее видам Ch. gud (Кавказ), Ch. roberti (Кавказ) и Ch. nivalis (Западная Европа), а одна относится к образцу Ch. nivalis из Турции (Chionomys sp.1), Ch. gud группируется с Ch. roberti, а Ch. nivalis из Западной Европы – с Ch. nivalis из Турции (Chionomys sp.1).
4.2. Внутривидовое генетическое разнообразие по результатам анализа мтДНК представителей рода Chionomys Кавказа
Помимо основных 5 групп на ML-дереве cytb хорошо различается множество подгрупп, которые относятся к тому или иному виду и отражают внутривидовую структуру снежных полевок. В составе Ch. gud выявлено не менее 3–4 таких подгрупп гаплотипов, для Ch. roberti – 2–3, для Ch. nivalis – 5–6. Эти гаплогруппы в большинстве случаев соответствуют определенным географическим локалитетам и, следовательно, определяют наличие у видов рода Сhionomys довольно четкой филогеографической структуры.
Филогеографическая структура Ch. gud. Использование любого из алгоритмов результировалось в появлении на дереве четырех групп в составе Ch. gud, которые обнаруживают прямую корреляцию с географической принадлежностью образцов: «Приэльбрусье», «Железноводск», «Безенги» и «Адыгея». Наиболее разнородной внутри себя оказываются выборки «Безенги» и «Адыгея», наиболее гомогенной – «Железноводск». Возможно, это может быть связано с возрастом данных популяций, «Безенги» и «Железноводск» характеризуются наиболее высокой генетической дистанцией, отделяющей их от остальных выборок. Для «Железноводска» это может объясняться относительной изолированностью популяции снежных полевок горы Развалка, что объясняет также и ее генетическую гомогенность.
Гаплотипы, полученные в результате выравнивания последовательностей cytb Ch. gud были связаны сетью пошаговых мутационных преобразований с помощью алгоритма медианных сетей гаплотипов в программе Network 4.5.0.0 (Bandelt et al., 1999).
Представленная на рис. 6 парсимониальная сеть наглядно демонстрирует существование четырех группировок гаплотипов. Всего в нашей выборке гудаурской полевки найдено 25 митохондриальных гаплотипов.
Размеры кружков в сети пропорциональны числу экземпляров, имеющих данный митохондриальный гаплотип. Расстояние между гаплотипами пропорционально количеству нуклеотидных замен, отличающих один гаплотип от другого (число этих замен приведено на интернодах). В междоузлиях сети располагаются рассчитанные программой гипотетические предковые гаплотипы.
Анализ медианной сети не только подтверждает результаты филогенетического анализа с использованием трех различных алгоритмов построения деревьев, но и наглядно показывает большую близость железноводских и безенгийских групп гаплотипов между собой, а также значительную удаленность обеих этих гаплогрупп и адыгейской гаплогруппы от гаплотипов Приэльбрусья (табл. 1).
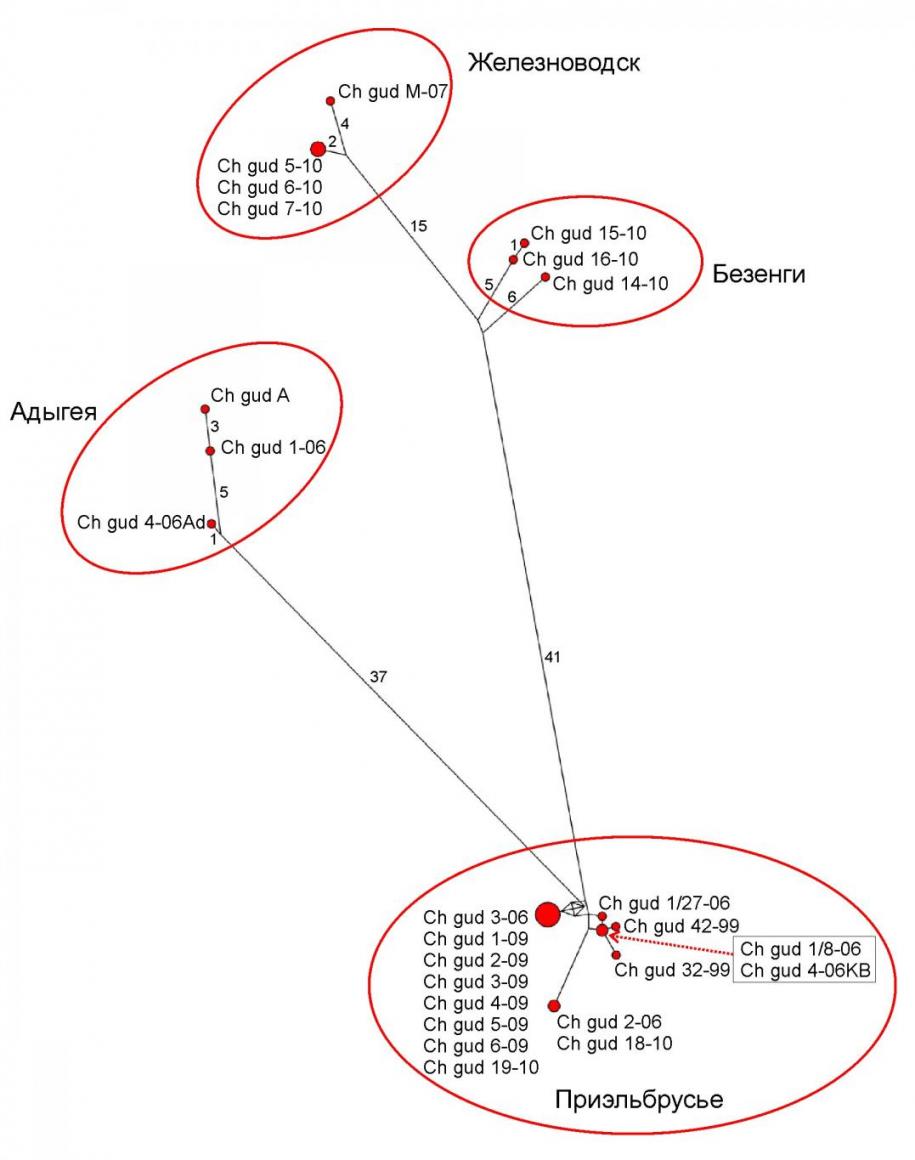
Рис. 6. Медианная сеть 25 гаплотипов cytb Ch. gud.
Размеры кружков пропорциональны числу образцов с соответствующим гаплотипом, длины интернод пропорциональны количеству разделяющих гаплотипы нуклеотидных
замен (числа на интернодах)
Таблица 1
Генетическое расстояние между географическими популяциями Ch. gud
Кавказа и средние значения внутрипопуляционных генетических дистанций
(над диагональю – некорректированная p-дистанция; под диагональю –
net-дистанция; на диагонали жирным шрифтом выделены средние значения
внутрипопуляционных генетических дистанций)
| p-/net-дист. %)±S.E. | Приэльбрусье | Безенги | Железноводск | Адыгея |
| Приэльбрусье | 0.39±0.11 | 3.93±0.52 | 4.12±0.54 | 3.59±0.60 |
| Безенги | 4.50±0.57 | 0.76±0.20 | 1.54±0.35 | 3.88±0.60 |
| Железноводск | 4.45±0.57 | 2.05±0.38 | 0.27±0.10 | 4.27±0.62 |
| Адыгея | 4.20±0.61 | 4.63±0.64 | 4.78±0.63 | 0.76±0.30 |
4.3. Анализ генетического разнообразия у исследуемых видов снежных
полевок
Ch. gud. Для анализа географической изменчивости Ch. gud выборка
(n = 25) была поделена на 4 подвыборки (условно географических популяций) в соответствии с географической принадлежностью гаплотипов: «Приэльбрусье», «Железноводск», «Безенги» и «Адыгея». Для оценки внутривидовой митохондриальной изменчивости Ch. gud в программе Arlequin 2.000 (Schneider et al., 2000) был произведен расчет показателей нуклеотидного () и гаплотипического (H) разнообразия. Результаты анализа представлены в табл. 2.
Таблица 2
Гаплотипическая и нуклеотидная характеристика популяций (приведены показатели только для выборок, содержащих не менее 3-х образцов).
H – гаплотипическое разнообразие, (%) – нуклеотидное разнообразие
| Выборка | Число образцов | Число гаплотипов | H | (%) | Tau |
| Приэльбрусье | 15 | 6 | 0.75 | 0.0045±0.0033 | 8.6621 |
| Безенги | 3 | 3 | 1.00 | 0.0087±0.0071 | 13.8125 |
| Железноводск | 4 | 2 | 0.50 | 0.0030±0.0022 | – |
| Адыгея | 3 | 3 | 1.00 | 0.0039±0.0032 | 4.4922 |
Из табл. 2 следует, что образцы с г. Развалка (окр. Железноводска) мало отличаются между собой по последовательности cytb, а наиболее сильно различающиеся между собой гаплотипы характерны для Безенги (=0.0087). Выборка из Безенги, несмотря на очень ограниченную территорию по сравнению с выборками из Адыгеи и Приэльбрусья, имеет также самый высокий показатель Tau.
Показатель Tau – это время, измеряемое в числе нуклеотидных замен на длину гена. Tau = 13.8 для популяции Безенги означает, что время происхождения данной популяции ~13.8 нуклеотидных замен назад, т.е. это время перехода данной популяции к стабильной численности. Если принять во внимание, что скорость замен в cytb у ближайших к снежным полевкам видам рода Microtus составляет не менее 30% за 1 млн лет (для самых недавних дивергентных событий) (Bannikova et al., 2010), то тогда возраст безенгийской популяции составляет ~40 000 лет (т.е. попадает в период Каргинского межледниковья), в то время, как возраст приэльбруской популяции (Tau = 8.7) из этих же расчетов составляет примерно 25 000 лет, т.е. относится к периоду последнего доледниковья, а адыгейская популяция еще моложе.
Ch. roberti. На дереве максимального правдоподобия (рис. 4), также в составе Ch. roberti различаются три клады, которые соответствуют географическому группированию образцов: (1) Грузия и Турция; (2) Адыгея-Лаганаки; (3) Адыгея-Гузерипль и Абхазия. Первая клада является базальной, в то время как две другие составляют сестринскую группу. Генетические дистанции между ними приведены в табл. 3.
Таблица 3
Генетическое расстояние между географическими популяциями Ch. roberti
и средние значения внутрипопуляционных генетических дистанций
(над диагональю – некорректированная p-дистанция; под диагональю –
net-дистанция; на диагонали жирным шрифтом выделены
средние значения внутрипопуляционных генетических дистанций)
| p-/net-дистанция (%)±S.E | Адыгея-Лаганаки | Абхазия+Гузерипль | Грузия+Турция |
| Адыгея-Лаганаки | 0.16±0.11 | 0.89±0.25 | 2.89±0.50 |
| Абхазия+Гузерипль | 1.02±0.25 | 0.18±0.08 | 2.45±0.44 |
| Грузия+Турция | 3.37±0.50 | 3.29±0.50 | 0.79±0.28 |
Ch. nivalis. Генетические дистанции между гаплогруппами Ch. nivalis приведены в табл. 4. Генетическое расстояние (net-дистанция) между образцами, относящимися к разным горным системам (Пиренеи, Альпы, Апеннины, Закарпатье, Понтийские горы при исключении образца из ЮЦ Турции) изменяется от 1.22±0.33 (Испания/Зап. Альпы и Апеннины) до 2.81±0.47 (Карпаты/Ближний Восток).
Как видно из выше изложенного материала, филогенетический анализ митохондриального гена cytb и двух ядерных экзонов выявил в проанализированной выборке снежных полевок пять четких филогенетических линий. Митохондриальный ген cytb и 11 экзон ядерного гена BRCA1 показывают на более раннее отделение Ch. nivalis от общего ствола Chionomys по сравнению с Ch. gud и Ch. roberti.
Таблица 4
Генетическое расстояние между гаплогруппами Ch. nivalis: (над диагональю –
некорректированная p-дистанция; под диагональю – net-дистанция, %).
Приведены дистанции между кластерами на рис. 6 (NJ) с бутстрэп-поддержкой
не менее 75 %. L1 – Ближний Восток, L2 – Балканы, L3 – Словакия, L4 – Альпы и Апеннины, L5 – Испания, L6 – Зап. Альпы и Апеннины, L7 – Карпаты, L8 – Турция
| p-/net-дист. (%)± S.E | L1 | L2 | L3 | L4 | L5 | L6 | L7 | L8 |
| L1 | 1.29±0.25 | 2.46±0.43 | 2.62±0.44 | 2.25±0.42 | 2.18±0.43 | 1.92±0.39 | 2.81±0.47 | 8.46±0.79 |
| L2 | 3.55±0.48 | 0.89±0.19 | 2.73±0.46 | 1.94±0.38 | 2.08±0.42 | 1.62±0.34 | 1.78±0.38 | 8.46±0.83 |
| L3 | 3.27±0.48 | 3.18±0.47 | - | 2.15±0.41 | 2.67±0.48 | 2.12±0.41 | 3.02±0.47 | 9.39±0.78 |
| L4 | 3.30±0.46 | 2.79±0.44 | 2.55±0.45 | 0.81±0.18 | 1.92±0.38 | 1.24±0.29 | 2.27±0.42 | 8.96±0.79 |
| L5 | 3.61±0.48 | 3.32±0.47 | 3.46±0.51 | 3.11±0.44 | 1.58±0.36 | 1.22±0.33 | 2.67±0.46 | 9.18±0.83 |
| L6 | 3.28±0.42 | 2.78±0.40 | 2.83±0.46 | 2.36±0.36 | 2.72±0.40 | 1.43±0.29 | 2.07±0.42 | 8.87±0.82 |
| L7 | 3.46±0.50 | 2.23±0.40 | 3.02±0.47 | 2.68±0.44 | 3.46±0.49 | 2.78±0.44 | – | 10.12±0.86 |
| L8 | 9.11±0.85 | 9.83±0.85 | 9.44±0.83 | 9.37±0.86 | 9.97±0.85 | 9.59±0.88 | 10.12±0.90 | – |
Внутривидовая генетическая изменчивость Ch. gud. Номинальный подвид Ch. g. gud, занимающий центральные части Главного Кавказского хребта, включает две группы гаплотипов – «Приэльбрусье» и «Безенги», причем они оказываются относительно не родственны друг от друга, поскольку «Безенги» при высокой поддержке образует сестринскую группу с гаплотипами реликтовой популяции Железноводска (подвид для этой части ареала не описан). К подвиду из западной части Главного Кавказского хребта, Ch. g. nenjukovi, относится гаплогруппа «Адыгея».
Порядок дивергенции четырех найденных гаплогрупп Ch. gud дает основание полагать, что приэльбрусская популяция является наиболее молодой по своему происхождению, в то время как возраст железноводской и безенгийской популяций наибольший.
Внутривидовая генетическая изменчивость Ch. roberti. Взаимоотношения трех найденных гаплогрупп Ch. roberti и порядок их размещения на дереве свидетельствуют в пользу более древнего происхождения закавказских популяций. Гаплотипы из северо-центральной Турции и Грузии занимают самое базальное положение. Гаплогруппы Абхазии (оз. Рица) и Адыгеи (Гузерипль и Лаганаки) могли бы сформировать монофилетические группировки, если бы не образец 3–06 из Гузерипля, гаплотип которого обнаруживает сходство с абхазскими гаплотипами. Своеобразие адыгейской популяции Лаганаки у Ch. roberti менее заметно, чем в случае с Ch. gud. Но ясно, что у обоих видов митохондриальные гаплотипы Адыгеи сильно отличаются от центрально-кавказских и закавказских. Это говорит о том, что истории одних и тех же географических популяций Кавказа могли быть сходными у разных видов со сходными экологическими требованиями.
В целом для всех снежных полевок можно определенно сказать, что филогеографическая структура их видов и генетическая дифференциация популяций определяются степенью обособленности горных систем и регионов, к которым они приурочены.
Глава V. Особенности пространственной организации видового населения представителей рода Chionomys на Кавказе и ее антропогенные изменения
Имеющиеся в научной литературе палеозоологические и современные данные свидетельствуют о том, что млекопитающие, в том числе представители рода Chionomys, на Кавказе составляют один из древних элементов природных экосистем региона. Остатки современных видов млекопитающих, в том числе рода Chionomys, известны с плейстоцена (Верещагин, 1959). С тех пор развитие орографии, формирование ландшафтной структуры, пространственной организации видового населения и ареалов отдельных видов и форм протекали как единый эволюционный процесс. В итоге сложились тесные и очень тонкие связи между ландшафтной структурой и современным размещением, внутривидовой изменчивостью и экологией млекопитающих Кавказа. По нашему мнению, без учета прошлых и современных ландшафтных условий трудно понять многие стороны жизни и организации исследуемых нами видов снежных полевок, особенно цито- и молекулярно-генетические особенности Ch.nivalis, Ch.roberti и Ch.gud, их изменения в ходе эволюции.
5.1. Распространение и биотопическая приуроченность снежной полевки
на Кавказе
Снежная полевка – Ch. nivalis по всем имеющимся у нас данным и литературным сведениям (Кулиев, 1978, 1979; Курятников, 1978; Дзуев, 1989, 1995) широко распространена в горах Закавказья. Она населяет горные леса и луга Месхетинского, Триалетского, Рагинского, Карталинского хребтов до горы Шахдаг включительно (рис. 7). Далее, на юго-востоке Малого Кавказа, снежная полевка прослежена в поясах темнохвойных лесов и субальпийскх лугов до Муровдагского хребта. Кроме того, она широко встречается по всей горной системе Малого Кавказа от Джавахетско-Армянского нагорья до Шахдагского и Карабахского хребтов на северо-западе, а на юго-востоке до Зангезурского хребта включительно.
В горно-степном поясе места обитания снежной полевки занимают весьма обширные площади и связаны в основном с каменистыми россыпями. В пределах всех остальных биотопов она живет не постоянно, а лишь заходит сюда, видимо, при кормежке и миграциях.
В субальпийском поясе этого региона она заселяет различные биотопы: субальпийские луга с каменистыми осыпями, злаковые, разнотравно-злаковые и отчасти разнотравные луга, очень редко субальпийское высокотравье с каменистыми осыпями. И, наконец, в альпийском поясе – альпийские луга, альпийские ковры, заросли рододендрона кавказского и т.д.
5.2. Структура ареала полевки Роберта на Кавказе
Как видно из рис. 7 на Западном Кавказе, где мезофильные луга и леса сохранились более или менее и занимают всю горную область, она широко распространена. Начиная с запада от верховья реки Белой, этот зверек прослежен нами на восток до реки Малой Лабы, Архыза и Теберды. В пределах Кавказского заповедника и прилегающих территорий Ch. roberti встречается в районе горы Абаго в субальпийском поясе на высоте 2000м, на Кордоне Умпырь на высоте 900 м. В окрестностях населенных пунктов Абадзехская, Даховская, Хамышки, Никель на высоте 400–500 м отмечена нами как многочисленный вид. В Тебердинском заповеднике малоазийская снеговая полевка добыта в поясе лесов и на границе леса в субальпике на высотах 800–2000 м.
Район Пятигорье-Эльбрус характеризуется значительным повышением сухости и континентальности климата во всех высотных ландшафтах, начиная от равнины и заканчивая альпийским поясом. На этой территории ни нами, ни предыдущими исследователями Ch. roberti не была отмечена.
В юго-восточной части северного макросклона Большого Кавказа она обнаружена лишь в верховьях рр. Аварское и Андийское Койсу на высотах от 1500 до 3000 м в лесистой части субальпийского пояса (Лавровский и Колесников, 1956; Верещагин, 1959). В Закавказье она имеет более широкое распространение. Встречается от Бзыбского хребта по Черноморскому побережью до Батуми, занимает горы Месхетинского, Лихского хребтов и южный макросклон Западного Кавказа.
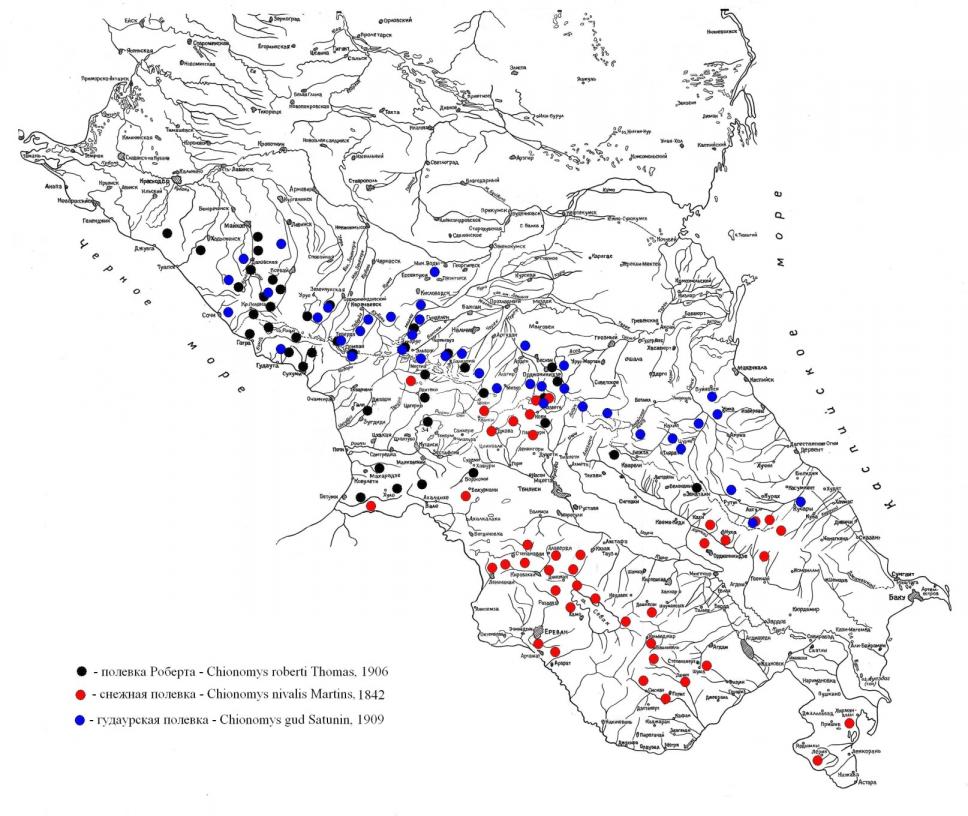
Рис. 7. Распространение снежных полевок на Кавказе
Как видно из вышеизложенного и литературных источников (Туров, 1926; Шидловский, 1948, 1958; Хасанова, 1990; Дзуев, 1995 и др.), полевка Роберта считается типичным представителем лесной фауны, размещение которой приурочено к кустарниковым зарослям берегов различных водоемов. По нашим данным и литературным сведениям, ареал этого зверька носит реликтовый характер на Кавказе.
5.3. Распространение и биотопическая приуроченность гудаурской полевки
Гудаурская полевка – Ch. gud свойственна лесному, субальпийскому и альпийскому поясам Северного Кавказа (рис. 7). На северном макросклоне Большого Кавказа юго-восточная граница распространения совпадает с Самурским хребтом, выступающим здесь хорошим рубежом между сухими ландшафтами северо-западного Закавказья и более или менее мезофильными горными лесами бассейна Самур. На южном макросклоне Большого Кавказа в связи с резкой ксерофитизацией горных ландшафтов, гудаурская полевка фактически не встречается как здесь, так и на всем протяжении Закавказья, Малого Кавказа и Талыша.
На Северном Кавказе гудаурская полевка привязана в каждом ущелье к определенным биотопам. Например, в междуречье Кумы и Подкумка она встречается в лесостепном поясе на высоте 500–600 м н.у.м. В долине реки Малка местообитания, хотя и привязаны к каменистым биотопам, но в разных высотных поясах заметно отличаются.
На ксерофитизированных лугах горного Дагестана гудаурская полевка сохраняет мезофильные черты своей экологии. Нами и другими исследователями (Гептнер и др., 1941; Верещагин, 1959; Дзуев, 1995 и др.) она замечена на поляне субальпийского пояса и у ручейков в окр. с. Аракани, по верхней границе букового леса, в увлажненных депрессиях, лощинах на субальпийских лугах в окр. с. Тлярота.
Снежные полевки – это ландшафтные виды более или менее мезофитных горнолуговых и горнолесных биоценозов. Оптимум ареала отдельных представителей этого рода находится в различных ландшафтных условиях. Ареал гудаурской полевки (Ch. gud) занимает северный макросклон Большого Кавказа с оптимумом ареала в субальпийском поясе. Распространение полевки Роберта (Ch. roberti) охватывает северный и южный макросклоны Большого Кавказа, а оптимум ареала находится в пределах лесного пояса с более или менее мезофитными биотопами. За пределом кавказской части ареала, по данным Н.К. Верещагина (1959), она встречается на Северных склонах восточного Тавра и Малой Азии. Современный ареал снежной полевки (Ch. nivalis) наиболее обширный и охватывает Средиземноморье – от Пиренеев до Балкан, она встречается в Сирии, Палестине, Малой Азии, в западном Копет-Даге. На Кавказе она достоверна по кариологически датированным данным от Бзыбского хребта на западном Закавказье до Талыша включительно, с оптимумом ареала на территории Малого Кавказа. Представители рода Chionomys на Кавказе четко различаются по морфологии, кариотипу, молекулярно-генетическому материалу, экологии, зоогеографии и т.д. Наиболее ксерофильный вид Ch. nivalis, мезофильный Ch. roberti, а Ch. gud занимает промежуточное положение по занимаемым экологическим нишам на Кавказе.
ВЫВОДЫ
- На основе комплексного изучения показана реальность существования на Кавказе 3 видов Chionomys: Ch. nivalis Martins, 1842; Ch. roberti Thomas, 1906; Ch. gud Satunin, 1909. Степень морфологической и молекулярно-генетической дивергенции, репродуктивной изоляции между ними существенно различна.
- Филогенетические отношения снежных полевок, установленные в результате анализа митохондриального гена cytb, позволяют различить в про-анализированной выборке пять четких филогенетических линий: Ch. nivalis, Ch. gud, Ch. roberti, Ch. nivalis ZIN2737 из Центрального Тавра и Ch. gud EU700087 из Ардахана (Турция).
- Анализ ядерных экзонов GHR и BRCA1 и комбинированной последовательности из обоих генов поддерживает найденные клады и общую топологию митохондриального дерева.
- Митохондриальный ген cytb и 11-й экзон ядерного гена BRCA1, а также комбинированная последовательность из ядерных генов показывают на более раннее отделение Ch. nivalis от общего ствола по сравнению с Ch. gud и Ch. roberti, что согласуется с известными морфологическими и кариологическими данными.
- Молекулярно-генетическое разнообразие рода Chionomys видового и внутривидового уровня выше предполагаемого до сих пор на основе морфо-логии и строения кариотипов.
- Особенности кариотипов снежных полевок на Кавказе стойкие и видоспецифичные: у Ch.nivalis 2n = 54, NF = 56; Ch.roberti 2n = 54, NF = 58 и Ch.gud 2n = 54, NF = 58.
- Пространственная организация видового населения и экология снежных полевок на Кавказе различны. Ареал Ch.nivalis ограничен закавказской частью региона, Ch.gud – занимает Северный Кавказ, а ареал Ch.roberti наиболее обши-рен, но носит реликтовый дизъюнктивный характер; изолированные поселения в наиболее мезофильных ландшафтах, как на Северном Кавказе, так и в Закавказье. Основным лимитирующим фактором распространения снежных полевок на Кавказе является наличие характерных биотопов, т.е. ландшафтные условия.
ПУБЛИКАЦИИ ПО МАТЕРИАЛАМ ДИССЕРТАЦИИ
- Банникова А.А., Маликов В.Г., Голенищев Ф.Н., Лебедев В.С., Сижажева А.М., Хуламханова М.М., Дзуев Р.И. Новые данные по разнообразию полевок рода Chionomys (Rodendia, Arvicolinae). Целостность вида у млекопитающих: изолирующие барьеры // Материалы конференции. – Петергоф, 2010. – С. 8.
- Дзуев Р.И., Сижажева А.М., Хашкулова М.А. Население млекопитающих горных экосистем центрального Кавказа (на примере Приэльбрусья) // XII Международная конференция «Биологическое разнообразие Кавказа». – Махачкала, 2010. – С. 308–310.
- Дзуев Р.И., Хашкулова М.А., Сижажева А.М. Эколого-географическая характеристика населения млекопитающих на Центральном Кавказе (на примере КБР) // XII Международная конференция «Биологическое разнообразие Кавказа». – Махачкала, 2010. – С. 315–317.
- Дзуев Р.И., Хуламханова М.М., Сижажева А.М. Молекулярная систематика и эколого-биологические особенности гудаурской полевки (Chionomys gud Satunin,1909) на Кавказе. – Махачкала: Эко-пресс, 2011. – 208с.
- Сижажева А.М., Хашкулова М.А., Башаева А.Х. Млекопитающие в Минераловодских лакколитах (на примере «вековой мерзлоты» горы Развалка) // Материалы международной научной конференции студентов, аспирантов и молодых ученых «Перспектива-2011. – Нальчик: Каб.-Балк. ун-т, 2011. – С. 62–66.
- Дзуев Р.И., Сижажева А.М. Пространственная структура ареалов снеговых полевок (Chionomys) на Кавказе // XIII Международная конференция «Биологическое разнообразие Кавказа». – Грозный, 2011. – С. 198–201.
- Сижажева А.М., Дзуев Р.И. Основные тенденции в распространении и численности мелких млекопитающих Кавказа, обусловленные динамикой природно-климатических факторов (на примере рода Chionomys). Юг России: экология, развитие. – Махачкала, 2011. – № 3. – С. 75–82.
- Сижажева А.М., Дзуев Р.И., Хашкулова М.А., Канукова В.Н. Хромосомный набор и эколого-биологические особенности гудаурской полевки (Chionomys gud Satutin, 1909) в условиях горы Развалка Ставропольского края // Труды Кубанского государственного аграрного университета. – Краснодар, 2011. – № 5(32). – С. 118–122.
- Дзуев Р.И., Сижажева А.М., Банникова А.А.. Внутривидовая генетическая структура кавказской снеговой полевки Chionomys gud Satunin, 1909 по результатам анализа митохондриальной ДНК // Известие КБГУ. – Нальчик: Каб.-Балк. ун-т, 2011. – T. 1. – № 4. – С. 35–42.
Лицензия ИД № 00003 от 27.08.99
Подписано в печать 02.03.12.
Формат 60х84 1/16. Бумага офсетная. Гарнитура Таймс.
Усл. печ. л. 1,1. Тираж 100 экз.
Издательство М. и В. Котляровых
(ООО «Полиграфсервис и Т»)
360051, КБР, г. Нальчик, ул. Кабардинская, 19 Тел./факс: (8662) 42-62-09