

Н. С. БАКШЕЕВ, И. К. ПАДЧЕНКО
МОЧЕПОЛОВОЙ ТРИХОМОНОЗ У ЖЕНЩИН
ИЗДАТЕЛЬСТВО «МЕДИЦИНА» МОСКВА — 1971
РЕФЕРАТ
Книга посвящена весьма распространенному заболеванию — мочеполовому трихомонозу. Авторы обобщили данные отечественной и зарубежной литературы по различным разделам мочеполового трихомоноза. В книге приводятся результаты собственных экспериментальных, лабораторных и клинических исследований. Книга написана на современном научном уровне и дает достаточно полное представление об учении о трихомонозе.
Работа представляет большой интерес для акушеров-гинекологов, так как в ней подробно излагаются вопросы эпидемиологии трихомоноза, его патогенеза, клиники, диагностики н лечения. Значительное внимание уделено таким важным вопросам, как профилактика и организация борьбы с мочеполовым трихомонозом.
ВВЕДЕНИЕ
Неустанная забота Советского правительства и Коммунистической партии Советского Союза об охране здоровья трудящихся обязывает медицинских работников всемерно улучшать организацию медицинского обслуживания насе ления, изучать сущность заболеваний, шире внедрять в повседневную жизнь профилактические мероприятия Усовершенствовать методы лечения.
К числу широко распространенных, но еще недостаточно изученные инвазий относится трихомоноз — болезнь, вызываемая трихомонадами. Необходимо отметить, что болезни, вызываемые трихомонадами у человека, составляют весьма важную проблему, имеющую как социальное, так и народнохозяйственное значение.
Данная инфекция у человека относится к группе так называемых негонококковых воспалительных заболеваний мочеполовых органов.
Установлено, например, что негонококкавые уретриты протекают клинически относительно тяжелее, чем гонорея, и вызывают больший процент осложнений со стороны по ловых органов человека. Это связано с недостаточным знанием вопросов эпидемиологии, несовершенствам диагностики и лечения, а также отсутствием официально узаконен ной системы учета больных данными ипфекциями, (трихо моноз, бактериальные, микотические уретриты и др.)
Изучение и анализ данных отечественной, и зарубежной литературы позволяют сделать заключение, что воспали-тельные заболевания мочеполовых органов нарушают тру доспособность человека. Нередко эти заболевания являют ся одной из причин, снижающих работоспособность жен щин и ее квалификацию, вследствие неблагоприятного влияния на состояние здоровья и возникновения в отдельных случаях бесплодия. Наличие трихомоноза нередко способствует возникновению у женщин тяжёлых воспали тельных заболеваний септической этиологии.
Результаты многочисленных наблюдений показывают, что в группе так называемых негонококковых воспали тельных заболеваний мочеполовых органов одно из ведущих мест принадлежит трихомонозу. В настоящее время считается установленным, что от 6,6 до 54,8% (в среднем от 15 до 30%) негонококковых уретритов вызываются трихомонадами.
В последние годы наблюдается тенденция увеличения всех видов негонококковых уретритов (трихомонадных, микробных, микотических и др.) по отношению к острой гонорее. Изменяется также соотношение между гонореей и негонококковыми уретритами в пользу последних (И. М. Порудоминский, 1960; И. И. Ильин, 1962; Siboulet, I960, и др.). Негонококковые уретриты превышают заболеваемость гонореей, как указывает И. М. Порудоминский (1963), в 1,5 —2 раза.
Некоторое представление о роли трихомоноза, занимающего одно из ведущих мест в группе так называемых негонококковых воспалительных заболеваний мочеполовых путей, можно составить хотя бы на основании того факта, что на долю больных трихомонадными кольпитами приходится до 20—40% всех обращений в гинекологическую и до 50—60% — в венерологическую амбулаторию (М. Ф. Елистратова, 1937; Jirovec, 1958, 1962, и др.). Среди первичных больных кабинета мужской гонореи число лиц, страдающих трихомонадной инфекцией, колеблется от 7,7 до 17,5% (А. М. Корчемкин, 1960). Согласно нашим наблюдениям, свыше трети всех процедур, производимых в течение года в манипуляционном кабинете женских консультаций, назначается по поводу мочеполового трихомоноза.
Установлено, что у инфицированных трихомонадами женщин наблюдаются значительные морфологические изменения воспалительного характера со стороны слизистой оболочки влагалища и шейки матки (Arboli, 1963). Имеются указания на возможность более частого возникновения рака шейки матки у данной группы больных (Pundel, Schwachtgen, 1957).
В данной работе предпринята попытка обобщить имеющиеся в отечественной и зарубежной литературе сведения по различным разделам трихомонадной инфекции и поделиться результатами собственных экспериментальных, лабораторных и клинических исследований. Особое внимание при этом уделено разработке вопросов эпидемиологии, лечения, профилактики и организации борьбы с трихомонозом мочеполовых органов человека. Мы надеемся, что наши предложения будут положительно восприняты практическими врачами и органами здравоохранения и окажут им значительную помощь в борьбе с этой широко распространенной инфекцией.
ГЛАВА I
КРАТКИЕ ИСТОРИЧЕСКИЕ СВЕДЕНИЯ
О МОЧЕПОЛОВОМ ТРИХОМОНОЗЕ
И ЕГО ВОЗБУДИТЕЛЕ
Первые сведения о трихомонозе женских мочеполовых органов появились в литературе в 1836 г., когда Donne обнаружил в гнойном влагалищном содержимом однокле точного жгутикового паразита животной природы, которого он назвал Trichomonos vaginale. Спустя 2 года, в 1938г., это определение протиста Ehrenberg изменил на Trichomonos vaginalis, которое и было утверждено интернациональной комиссией по зоологической номенклатуре в 1926 г.
Следует, однако, отметить, что Donne не считал обнаруженного им паразита возбудителем какого-либо заболевания. Описывая в 1855 г. морфологию трихомонад, Kolliker и Scanconi также считали их безвредными обитателями половых органов. Между тем наиболее благоприятные ус-
ловия для развития этих паразитов создавались, по мнению этих авторов, в случае воспаления слизистой оболочки влагалища.
В 1870 г. наш соотечественник, профессор акушерской клиники в Харькове И. П. Лазаревич, подтвердил наличие трихомонад во влагалищном содержимом, подробно описал их морфологию, частично изучил физиологию и впервые научно доказал наличие связи между этими протистами и воспалением слизистой оболочки влагалища и шейки матки. В монографии «Паразиты женских половых органов» в
, 1870 г. он писал, что у пораженных трихомонадами лиц наблюдается разрыхление и изъязвление слизистой оболочки, а влагалищное содержимое становится гноевидным, пенистым, реакция его — слабокислой или нейтральной.
По мнению И. П. Лазаревича, заражение женских половых путей происходит при попадании возбудителей извне и чаще всего наблюдается в весеннее время, Онi трихомонадная инфекция женских мочеполовых помимо чисто локальных измедений в тканях, оказывает также влияние на общее состояние организма. В доказательство последнего принципиальна важного утверждения он приводит выписки из историй болезни двух больных, страдавших одновременно истерией и трихомонозом.
После излечения этих больных от трихомоноза припадки
истерии становились слабее и затем полностью прекратились.
И. П. Лазаревич указывает также надо обстоятельство, что иногда наличие трихомонад во влагалище может вовсе не вызывать болезненных явлений, т. е. инфекция способна протекать в ряде случаев в виде так называемого бессимптомного носительства этих паразитов.
Для борьбы с заболеванием он предложил ряд профилактических и лечебных мероприятий, указав на необходимость в каждом отдельном случае лечить также сопровождающие трихомоноз болезненные состояния.
К. Ф. Славянский в 1888 г., описывая различные клинические формы воспаления слизистой оболочки влагалища, указывал, что в случаях затянувшейся болезни, особенно у беременных, в вагинальных выделениях нередко можно обнаружить трихомонады. При этом он обратил внимание на то, что в хронических случаях заболевания жалобы на боли и бели могут совершенно отсутствовать.
Спустя 46 лет после выхода в свет монографий И. П. Лазаревича Hoehne (1916) описал 12 случаев трихомонадного кольпита, указав, что его причиной было попадание трихомонад во влагалище. Исчезновение грихомонад из влагалища способствовала излечению кольпита.
Таким образом, открытие возбудителя трихомонадной инфекций принадлежит Donne, который впервые обнаружил протиста. Связь между наличием трихомонад и воспалительным процессом слизистой оболочки влагалища и шейки матки впервые установили отечественные авторы И. П. Лазаревич и К. Ф. Славянский. Этот факт опровергают имеющиеся в литературе указание Jirovec, Peter, И. А. Евнина, Н. А. Лапидуса (1939), Б. В. Вертинского (1958), Б. А. Теохарова (1959) и некоторых других авторов, приписывающих установление указанной связи Hoehne.
Морфологически схожего с Trichomonas vaginalis жгутиконосца, паразитирующего в кишечнике человека, изве ныне под названием trichomonaa intestmalis, вперые выделил из испражнений холерного больного и описал в литературе французский паразитолог Devalue в 1854 г, В последующие годы этот паразит описывался многими авторами под самыми различными названиями, чему немало способствовало его внешнее сходство с другим ви дом, обитающим в кишечнике человека и именуемым в в настоящее время Chilomastix mesnili. Трихомонаде, паразитирующей в содержимом кишечного тракта человека, окончательно было дано название Trichomonas intestinalis, s. hominis Davaine (1854) —кишечная трихомонада.
В 1862 г. русский исследователь О. Штейнберг открыл и описал похожего на Trichomonas vaginalis жгутикового протиста, паразитирующего в полости рта. По своим морфологическим особенностям данный паразит так же, как и кишечная трихомонада, был отнесен к роду Trichomonas (трихомонад),
Впоследствии эту трихомонаду Wenyon (1926) назвал Trichomonas elongata, Steinberg (1862)—ротовая трихомонада.
Вопрос о самостовтельности видов трихомонад, обнаруживаемых в мочеполовых органах,киишечнике и полости рта человека, остается до сих пор окончательно не решенным.
Первое сообщение об обнаружении трихомонад в выделениях из половых органов девочек было сделано. Hausmann в 1870 г. В более поздний период факт обитания трихомонад в половой сфере девочек был подтвержден результатами многочисленных исследований (М. Ф. Елистратова, 1937; П. Д. Давыдов, 1951; Г. Р. Робачев-ский, 19&2, 1954; В. А. Сапожкова, 1956; М, А. Тимохина, 1958,;1959; А, А, Радионченко-Кулакова, 1960; В. И.Рыбников, 1964; Feo, W56; Komorowska, Liniecka, Kurnatowska, 1961, 1962, и др.) Однако далеко еще не все авторы допускают возможность паразитирования трихомонад у девочек в различные периоды их жизни. В частности, Peter (1945), Jirovec (I960) и др. считают, что для этих протистов не имеется подходящих условий для развития во влагалище девочек в, период так называемого покоя половых органов (в возрасте от 1 года до 10 лет) ввиду отсутствия в их организме эстрогенов. Напротив, Feo (1958) путем культивирования трихомонад на питательных средах, лишенных мальтозы, установил, что гликоген эпителия вагины, появляю щийся в pesyjjbtaTe эстрогенной стимуляции, не является необходимым факторам роста для этих паразитов. По его •мнению, трихомонады способны обитать как во влагалище девочек и девушек, так и женщин различного возраста, Накопленные за последние годы в отечественной и зару* бежной литературе многочисленные фактические данные и результаты наших исследований позволяют считать, что это положение является в настоящее время окончательно доказанным.
Об обнаружении трихомонад в моче мужчин впервые сообщили в 1894 г. Marchand и Miura. В 1927 г, чешский венеролог Capek сообщил о двух случаях трихомонадного уретрита у мужчин, возникшего вследствие полового общения с женщинами, страдавшими трихомоладиым кольпитом.
Этим самым он первым указал на венерический характер болезни, вызываемой трихомонадами. Ему также принадлежит мысль о 'необходимости лечения при этой инфекции обоих супругов. В последующие годы в содержимом уретры мужчин трихомонад выявляли А. Д. Ротнищшй (1938), В. Н. Матвеев (1939) и др. Особенно большое число сообщений fo значительной распространенности трихомоноза среди мужчин появилось после 1944—1945 гг. (Б. В. Вер-шинский, 1958; Б. А. Т^охарощ, 1959; А. М. Корчемкин, 1960; М. С. Месинев, 1960; С. Л. Козин, 1961; А. Ф. Аникин, 1964; Feo, 1944; Lanceley, 1955; Coutts, 1955; Hoffmann, Malyszko, 1962, и др.).
Короткий перечень лишь некоторых работ, выполненных в послевоенные годы отечественными и зарубежными исследователями по различным вопросам, касаГющимся проблемы мочеполового трихомоноза мужчин, является вполне убедительным доказательством того факта, что трихомонады приспособились в процессе эволюции паразитировать в мочеполовых органах не только женщин, но и мужчин.
В настоящее время считается установленным, что у мужчин трихомонады могут локализоваться, кроме уретры, в предстательной железе, семяпроводах, мочевом пузыре, куперовых железах и других-отделах мочеполовых органов (И. М. Исмаил-Заде, 1948; А. Ф. Аникин, 1964; Lanceley, 1955; Б. Гофман и Е. Малышко, 1962; Preisler, 1960; Palazzetti, 1959, и др.).
У женщин этих протистов находили на коже бедер, на наружных половых органах, в бартолиновых железах,
влагалище, цервикальном канале, уретре, скеновых железах, околоплодной жидкости, моче, взятой из мочевого пузыря катетером. Т. Б. Альбицкая (1955) обнаруживала трихомонад в маточном секрете и маточных трубах. Становится очевидным, что трихомонады локализуются у мужчин и женщин в различных отделах как половой сферы, так и мочевыводящих путей.
Исходя из этого, некоторые авторы (И. М. Исмаил-За* де, 1948; Б. С. Пойзнер, 1960; Roth, 1944; Palazzetii, 1959, и др.) считают, что термин Trichomonas vagmalis, присвоенный возбудителю мочеполового трихомоноза еще в прошлом веке, является устаревшим и поэтому подлежит замене.
При этом Roth (1944) предлагает называть этих протестов Trichomonas genitalis, а И. М. Исмаил-Заде (1948) — Trichomonas urogenitalis, т. е. мочеполовыми или урогени-тальными трихомонадами.
Результаты изучения и анализа литературных материал лов, а также собственных наблюдений убеждают в том, что принятый в 1957 г. 'на Международном симпозиуме bl Реймсе термин «влагалищная трихомонада» действительно не дает полного представления как о свойствах самого возбудителя, так и о характере и клинических формах проявления вызываемой им болезни. Поэтому есть основание согласиться с мнением тех авторов, которые рекомендуют считать его устаревшим, а следовательно, и подлежащим замене.
Из числа упоминавшихся терминов ^следует остановиться, по нашему мнению, на том, который наиболее полно характеризует основные свойства этих протистов. Этим требованиям больше всего соответствует, как ;это следует из литературных материалов, характеризующих действительное состояние настоящего вопроса, название, данное И. М. Исмаил-Заде (1948), «урогенитал^ная трихомонада» (Trichomonas urogenitalis). На этом же основании следует согласиться и с мнением Palazzetti (1959), рекомендующим называть ч вызываемую этими протестами болезнь урогенитальным, или мочеполовым, трихомонозом.
Таким образом, резюмируя полученные данные, можно заключить, что трихомонады обитают в мочеполовых органах (Trichomonas vaginalis; Donne, 1836), кишечнике (Trichomonas intestinalis, s. hominis; Davaine, 1854) и в полости рта (Trichomonas elongata; Steinberg; 1962).
В настоящее время окончательно доказано* что возбудитель мочеполового трихомоноза способен, паразитировать в различных отделах мочеполовых органов не только женщин (влагалище, цервикальный канал, уретра, барто-линовы железы, скеновы железы, мочевой пузырь it Др*)г но и мужчин (уретра, предстательная железа, купе|юш железы и др.). В связи с этим термин «влагалищная трихомонада» (Trichomonas vaginalls) как устаревший и не характеризующий основных свойств возбудителя трихомоноза следует заменить другим. Мы полностью разделяем мнение тех авторовг которые предложили называть данного протиста урогенитальной трихомонадой (Trichomonas urogenitalls), а вызываемое им заболевание — мочеполовым, или урогенитальным, трихомонозом
ГЛАВА II
СОВРЕМЕННОЕ ПРЕДСТАВЛЕНИЕ О БИОЛОГИИ ВОЗБУДИТЕЛЯ МОЧЕПОЛОВОГО ТРИХОМОНОЗА
СИСТЕМАТИКА УРОГЕНИТАЛЬНЫХ ТРИХОМОНАД И ВИДОВАЯ ИХ САМОСТОЯТЕЛЬНОСТЬ
Урогенитальная трихомонада (Trichomonas urogenitalis, s. vaginalis) — одноклеточный паразит животной природы, приспособившийся в процессе эволюции к обитанию в мочеполовых органах человека. По зоологической номенклатуре она относится к типу простейших (Protozoa), классу жгутиковых (Flagellata или Mastigophora), отряду Protomonadidae, роду Trichomonas.
Выше уже отмечалось, что трихомонады паразитируют в организме человека не только в мочеполовых путях (Trichomonas urogenitalis), но также в кишечном тракте (Trichomonas intestinalis, s. hominis) и в полости рта (Trichomonas elongata, s. tenax, biicalis). Вопрос о самостоятельности каждого из упомянутых видов трихомонад (рис. 1) уже многие годы привлекает пристальное внимание как отечественных, так и зарубежных исследователей. Для его решения применялись самые разнообразные методы. Например, Wenrich (1947) и Jirovec (1960, 1962) использовали для этой цели метод морфологического сравнения трихомонад, обнаруживаемых в различных органах и системах (мочеполовые органы, кишечник, полость рта), а также обобщили литературные сведения о степени распространения этих протистов среди различных групп населения. При этом было констатировано, что каждый из описанных видов трихомонад наряду с общими признаками имеет ряд характерных особенностей, сохраняющихся, как указывает Jirovec (1962), /даже в случае культивирования паразитов на питательных средах. На этом основании авторы считают, что в организме чело-века паразитируют по меньшей мере три самостоятельных вида трихомонад.
Ссылаясь на различную чувствительность трихомонад, обитающих в мочеполовых органах и кишечнике человека, в желудочном соке и дуоденальном содержимом, Г. М. Тка ченко (1955) также относит этих протистов к различным зоологическим видам.
В настоящее время считается установленным, что трихомонады трех локализаций отличаются друг от друга по своей биохимической активности (Б. А. Теохаров, 1959, 1962; Ю. X. Терас, 1961, и др.), лекарственной чувствительности и патогенности.
ротовых трихомонад- во влагалище женщин. В подобных экспериментах, совершенно недопустимых в условиях нашего социалистического общества, участвовало 100 женщин-«волонтерок». В результате этих исследований стало известно, что кишечные трихомонады во влагалище некоторых женщин не обнаруживались уже спустя 30 минут после их введения. Лишь у одной женщины они оставались жизнеспособными во влагалище в течение 24 часов. При введении в вагину женщинам трихомонад, выделенных из полости рта, у 5 женщин протистов обнаруживали в вагинальном секрете в течение 48 суток, а у 3 женщин — 72—168 суток. Ни у одной из этих женщин не наблюдалось клинически выраженного заболевания, напоминающего трихомонадный кольпит. Не обнаружив у трихфмонад способности к изменению присущей им локализации, авторы считают, что в различных органах и системах человеческого организма обитает не менее трех зоологических видов трихомонад.
Следует констатировать, что точку зрения о существовании трех видов трихомонад — паразитов человека — не разделяют такие авторы, как Б. С. Пойзнер и А. А. Кулакова (1952), Э. И. Слапак (1955), В. А. Сапожкова (1956), Gro-let, Montauge (1955) и др. В частности, они считают, что между трихомонадами, паразитирующими в различных органах и системах человеческого организма, не имеется существенных различий и что при определенных условиях эти протесты могут не только менять присущую им локализацию, но и адаптироваться к изменившимся условиям среды обитания.
Весьма оригинальный путь избрала-для доказательства данного положения В. А. Сапожкова (1955, 1956), Она исследовала содержимое полости рта, кишечника и мочеполовых "органов новорожденных первых 2—3 дней жизни, родившихся от матерей, страдавших трихомонадными кольпитами. В этом случае 2,3 %! обследованных новорожденных девочек оказались пораженными кишечными трихомонадами. Паразиты были обнаружены в содержимом кишечника. Так как некоторые новорожденные-^ыли извлечены при кесаревом сечении, то возможность заглатывания ими отделяемого родовых путей, содержащих трихомонады, полностью исключалась. Отвергается автором также возможность занесения новорожденным трихомонад и во время ухода за ними лиц из числа обслуживающего персонала, ибо последние оказались свободными от кишечных
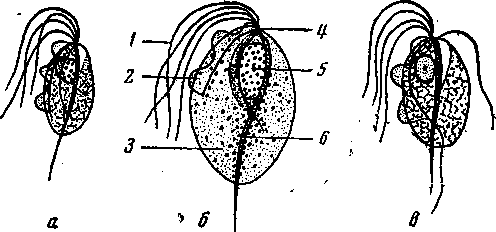
Рис. 1. Схематическое изображение трихомонад, паразитирующих у человека (по Хегнеру с соавторами). а — ротовая трихомонада (Trichomonas elongata, s. te-nax); б — урогенитальная трихомонада.(Trichomonas urogenitalis, s. vaginalis); в — кишечная трихомонада (Trichomonas intestinalis); / — жгутик; 2 — ундулирую-щая мембрана; 3 — цитоплазма; 4 — блефаропласт, 5 — ядро; 6 — аксостиль.
Результаты серологического типирования трихомонад, полученные при помощи реакций агломерации, связывания комплемента и агглютинации, свидетельствуют о том, что урогенитальные трихомонады имеют существенные отличия в антигенном отношении от кишечных и ротовых (О. И. Олейник, 1961; Ю. X. Терас, 1961, и др.). ИсхЬдя из этих данных, указанные авторы считают, что урогенитальные трихомонады являются самостоятельным видом.
Такую же позицию занимают в этом вопросе Л. Е. Чалая (1959), Б. А. Теохаров (1959), Cerva, Cervova (1961) и другие авторы на том основании, что им не удалось найти частого совпадения одновременного паразитирова-ния трихомонад в полости рта, кишечнике и мочеполовых органах обследованных женщин.
Вопрос о самостоятельности видов трихомонад, имеющих различную локализацию в организме человека некоторые авторы (Stabler, Feo, Rakoff, 1941; Stabler, Feo, 1942), пытались разрешить путем перевивки кишечных и ротовых трихомонад во влагалище женщин. В подобных экспериментах, совершенно недопустимых в условиях нашего социалистического общества, участвовало 100 женщин-«волонтерок». В результате этих исследований стало известно, что кишечные трихом он ады во влагалище некоторых женщин не обнаруживались уже спустя 30 минут после их введения. Лишь у одной женщины они оставались жизнеспособными во влагалище в течение 24 часов. При введении в вагину женщинам трихомонад, выделенных из полости рта, у 5 женщин протестов обнаруживали в вагинальном секрете в течение 48 суток, а у 3 женщин — 72—168 суток. Ни у одной из этих женщин не наблюдалось клинически выраженного заболевания, напоминающего трихомонадный кольпит. Не обнаружив у трихсщонад способности к изменению присущей им локализации, авторы считают, что в различных органах и системах человеческого организма обитает не менее трех зоологических видов трихомонад.
Следует констатировать, что точку зрения о существовании трех видов трихомонад — паразитов человека —не разделяют такие авторы, как Б. С. Пойзнер и А. А. Кулакова (1952), Э. И. Слапак (1955), В. А. Сапожкова (1956), Gro-let, Montauge (1955) и др. В частности, они считают, что между трихомонадами, паразитирующими в различных органах и системах человеческого организма, не имеется существенных различий и что при определенных условиях эти протисты могут не только менять присущую им локализацию, но и адаптироваться к изменившимся условиям среды обитания.
Весьма оригинальный путь избрала для доказательства данного положения В. А, Сапожкова (1955, 1956). Она исследовала содержимое полости рта, кишечника и мочеполовых органов новорожденных первых 2—3 дней жизни, родившихся от матерей, страдавших трихомонадными кольпитами. В этом случае 2,3%! обследованных новорожденных девочек оказались пораженными кишечными трихомонадами. Паразиты были обнаружены в содержимом кишечника. Так как некоторые новорожденные-были извлечены при кесаревом сечении, то возможность заглатывания ими отделяемого родовых путей, содержащих трихомона-ды, полностью исключалась. Отвергается автором также возможность занесения новорожденным трихомонад и во время ухода за ними лиц из числа обслуживающего персонала, ибо последние оказались свободными от кишечных и урогенитальных трихомонад. Следует подчеркнуть* что у 4 новорожденных трихомонады были обнаружены" не только в кишечнике, но и в околоплодной жидкости. Учитывая возможность интроамниального проникновения про-тистов через неповрежденный плодный пузырь, а^тшщё факт заглатывания плодом во второй половине бйреш^ ности околоплодных вод, автор полагает, что от длительного и многократного проникновения урогенитальных три* хомонад в кишечнике плода происходит адаптация протестов к необычным для них условиям обитания. По нашему мнению, сам акт проникновения трихомонад в кишечник плода — это свидетельство исторически выработавшейся способности паразита к существованию, результат борьбы за сохранение вида в природе.
Изучая устойчивость урогенитальных трихомонад в воде, молоке, сметане и желудочном соке, В. И. Карелина (1959) пришла к выводу, что они способны проходить через желудок с указанными пищевыми продуктами и водой также у взрослых лиц, особенно при пониженной кислотности содержимого желудка.
-. На возможность проникновения трихомонад из кишечного тракта во влагалище указывают М. Ф. Елистратова (1937), Г. Р. Робачевский (1954), Э. И. Слапак (1955) и др. Трихомонад, паразитирующих^ в полости рта, кишечнике и мочеполовых органах, следует отнести, по мнению этих авторов, к одному зоологическому виду. Аналогичную позицию занимают в данном вопросе также Б. С. Пойзнер, А. А. Кулакова \1952), В. А. Сапожкова (1956) и др,
В литературе имеются и такие взгляды, согласно кота-рым ротовые трихомонады по своим биологическим свойствам находятся значительно ближе к урогенйтальным, чем к кишечным (Mandoul, Pestre, Domenech, Laeroix, 1957). Основанием для подобного утверждения является различная способность, имеющая место у этих протестов,-к приживлению в половых органах подопытных животных (белых крыс), предварительно подвергшихся кастрации, а затем воздействию бензоатэстрадиола.
Таким образом, результаты изучения и анализ доступной нам отечественной и зарубежной литературы свще-тельствуют о том, что вопрос о видовой самостоятельности трихомонад, имеющих различную локализацию в человеческом организме, остается до сих пор фактически не решенным. Решить его методом морфологического сравнения, как это делают Wenrich (1947), Jirovec (I960, 1962) и др.., по нашему мнению, также не представляется возмож-шм из-за большого сходства, существующего между уро-дренитальными, кишечными и ротовыми трихомонадами. Р110тлнчйть их друг от друга удается лишь по размерам, >^лдлйне ундулирующей мембраны и количеству жгутиков. ?'~Т§ частности, длина-тела урогенитальной трихомонады со-: ^тавляет, по Wenrich (1947), 13 мк, кишечной —7—8 мк, ротовой — 6,5 мк. Кишечная трихомонада в противоположность урогенитальной имеет не 4, а 3—5 жгутиков и достигающую заднего конца тела ундулирующую мембрану, заканчивающуюся свободным рулевым жгутом (см. рис. 1). В остальном же эти три протиста морфологически почти -т не различимы. Мы допускаем, что имеющиеся морфологические различия между трихомонадами, паразитирующими в полости рта, кишечнике и мочеполовых органах, зависят от того, насколько каждая из упомянутых сред обитания способна удовлетворять пищевые потребности этих протестов. Судить поэтому о единстве видов трихомонад, паразитирующих у человека, на основании указанных выше признаков, очевидно, не имеется достаточных оснований. В ШШ z отсутствием окончательного ответа по данному вопросу возникают затруднения при решении некоторых противоэпидемических мероприятий, имеющих чисто практическое значение. В частности, практическим работникам органов здравоохранения пока не представляется возможности организовывать при мочеполовом трихомонозе профилактические мероприятия таким образом, чтобы они одновременно были направлены против ротового и кишечного трихомоноза. Мы считаем, что необходимо учитывать при разработке организационных мер борьбы с мочеполовым трихомонозом человека наличие трихомоноза других ло* кализаций.
МОРФОЛОГИЯ, ФИЗИОЛОГИЯ, ЦИКЛ РАЗВИТИЯ И РАЗМНОЖЕНИЯ УРОГЕНИТАЛЬНЫХ ТРИХОМОНАД
Еще И. П. Лазаревич (1870) писал, что трихомонады могут иметь грушевидную, яйцевидную, ланцетовидную и другие формы. В настоящее время считается установленным, что форма этих протистЬв непостоянна (см. рис. 2, 3). В вагинальных выделениях наиболее часто обнаруживают трихомонад, имеющих грушевидную, овальную и круглую формы, реже — веретенообразную или почковидную. Напротив, в содержимом мужской уретры и во влагалище де вочек преобладают круглые протесты (Б. П. Метальников с соавторами, 1959; М. С. Месинев, 1960; А. Ф. Аникин, 1964; В. И. Рыбников, 1964, и др.). Методом получения культур из круглых трихомонад была доказана возможность перехода указанных форм в грушевидную (Б. Ф. Пе-черский, 1951; Ю. X. Терас, 1959, и др.). Округлую форму протесты приобретают, по Ю. X. Терасу (1954), в неблагоприятных условиях. По мнению В. А. Лосицкой (1926), Н. Е. Сидорова, А. М. Корчемкина, А. П. Колесова (1959) и др., форма трихомонад зависит от фазы менструального цикла. Перед менструацией протесты, как правило, имеют грушевидную форму, после нее — круглую, а спустя еще неделю они снова становятся грушевидной формы. В литературе имеются также указания, согласно которым наблюдаемая у трихомонад изменчивость формы может быть связана с колебаниями рН среды, воздействием антибиотиков и т. д. (И. И. Ильин, 1962; Kupferberg, Johnson, Sprin-се, 1948). Согласно нашим данным, полученным при микроскопическом исследовании нативных мазков и препаратов, окрашенных азур-эозином по Романовскому, основной биологической формой существования урогенитальных трихомонад является грушевидная, тогда как другие разновидности форм паразитов появляются лишь на определенной стадии развития. В частности, вытянутые, веретенообразные и т. п. формы протистов образуются в процессе их размножения, а именно на стадии деления материнской клетки на дочерние особи. Становится очевидный, что полиморфизм паразитов возникает в результате появления среди них делящихся особей, т. е. в процессе размножения трихомонад. Последний может быть активизирован, как показали наши наблюдения (И. К. Падченко, 1964), многими агентами (воздействие химическими веществами и мочой, изменение температурного режима и др.)» в том числе и антибиотиками. Исходя из этого, имеются основания считать, что наблюдавшийся некоторыми авторами полиморфизм трихомонад при колебаниях кислотности среды и воздействии на них антибиотиков объясняется именно ускорением процесса размножения, что способствует увеличению возможности обнаружения в вагинальном секрете протистов, имеющих различную форму. Вполне возможно, что и при менструальном цикле также происходит активизация процесса размножения трихомонад.
Размеры урогенитальных трихомонад (рис. 2), согласно данным отечественных и зарубежных авторов (Г. М. Тка-
енко, 1955; Б. В. Вершинский, 1958; И. А. Лаан, 1961; Wenrich, 1947; Jirovec, 1960, и др.), колеблются от 7 до il5—60 мк. Средние размеры протестов составляют 10— 2 мк. Мелкие формы трихомонад |(8—ПХ4—6 мк) чаще обнаруживают при остром трихомонозе, а крупные (35—45 мк) и средние х (15—35 мк)—при хроническом i (Jirovec, 1960). Таким образом, ^становится очевидным, что возможность обитания в мочеполовых органах человека трихомонад различных размеров является в настоящее время установленным фактом.
Аналогичное явление наблюдается также при культивировании протистов на питательных средах. Следует, однако, отметить, что причина появления трихомонад различных размеров (в том числе карликовых и крупных, до 40— 60 мк) как в выделениях больных, страдающих трихомонозом, так и на питательных средах остается пока окончательно не распознанной. Между тем результаты наших исследований (1960, 1964) свидетельствуют о том, что крупные трихомонады представляют собой промежуточную, далеко не постоянную стадию в цикле развития данного протеста. В культурах они обычно появляются, как будет показано ниже, при неблагоприятных условиях окружающей среды. По своей сущности крупные паразиты — это особи, находящиеся в состоянии множественного деления (см. рис. 7, 8, 9). Мелкие подвижные, или так называемые карликовые, трихомонады являются юными формами протеста, которые также образуются в процессе размножения. Однако возникают они вследствие неравномерного распределения протоплазмы материнской клетки между дочерними особями. Тело урогенитальных трихомонад состоит из оболочки( пелликулы), протоплазмы, блефаропласта, ризопласта, ядра, аксостиля, жгутиков, ундулирующей мембраны, под-(краевой) и парабазальной фибрилл (см. рис. 1,2,3,4).
Оболочка у живых паразитов незаметна. Она становится видимой лишь на препаратах, окрашенных трипанблау или сафранином после фиксации осьмиевой кислотой (Г. И. Пе-рекропов и Е. А. Галкина, 1930; А. М. Корчемкин, 1959, и др.). Протоплазма содержит пищевые вакуоли и хромати-новые зерна, лежащие по сторонам аксостиля. Иногда в протоплазме обнаруживают заглоченные бактерии, лейкоциты и сперматозоиды (А. П. Колесов, 1950; Н. Е. Сидоров с соавторами, 1959, и др.). Следует, однако, заметить, что жгутиконосцы, взятые из содержимого влагалища, лишь в исключительных случаях содержат пищеварительные вакуоли (Jirovec, 1960) и заглоченные бактерии, хотя в культурах подобное заглатывание бактерий наблюдается часто. В передней трети тела паразита расположено продолговато-овальной формы ядро (см. рис. 2, 4), заключенное в нежную оболочку и содержащее одну кариозому, видимую лишь на влажно фиксированных препаратах, мелкие равномерно рассеянные гранулы хроматина (Wenrich, 1947; Jirovec, 1960). Впереди ядра, на переднем конце тела паразита, располагается блефаропласт, являющийся центром аппарата. От блефаропласта берут свое на-жгутики, ундулирующая мембрана, аксостиль, подбирающая и парабазальная фибриллы. Блефаропласт соединяется с ядром посредством короткого, стержневидной формы ризопласта.
Урогенитальная трихомонада, как правило, имеет четыре передних жгутика, состоящих из двух частей — аксонемы, идущей от блефаропласта до переднего конца тела паразита, и собственно жгутика, находящегося вне его тела и покрытого перипластом. В отдельных случаях паразит снабжен пятью жгутиками (3. С. Ярмухамедова, 1959). Длина жгутика составляет 7—8 мк. Иногда встречаются особи с одним жгутиком или вовсе без жгутиков (Ю. X. Те-рас, 1954; Г. М. Ткаченко, 1955, и др.). Объясняется это тем, что жгутики могут отламываться в процессе фиксации и окраски препаратов.
Волнообразная или ундулирующая мембрана, представляющая собою дупликатуру оболочки, простирается от переднего конца тела паразита к заднему на протяжении Двух третей его длины. На границе между телом протиста и волнообразной мембраной проходит подпирающая фиб-
C волнообразной мембраной аппарат паразита.
Парабазальная фибрилла идет от блефаропласта к аксо-стилю внутри тела протеста, огибая на своем пути ядро.
опорную функцию у паразита, иврсвое начало от блефаропласта и заканчивается на задаём конце теЛа протеста. Он ^Представляет собой тонкую сидерофильную нить. Задний конец аксостиля ригидный.
А. Евнин, Н. А. Дапидус, 1923, и др.) о существовании у три-хомонады ротового отверстия (цитостома) не нашли своего подтверждения в отечественной и зарубежной литературе^
Изучая сверхтонкие срезы урогенитальной трихомонады методам электронной микроскопии, Inoki Shozo и соавторы (195§i 1Щ)) установили, что ядро протиета окружено двухслойной мембраной, имеющей поры. Цитоплазма гйрйфе-рин б^рйзует стенку клетки — перипласт. В цитоплазме находятся гранулы величиной 5—12 мк, различных размеров вакуоли и эндоплазматической ретикулум. Ясно видны волокна с поперечной полосатостью, поднимающиеся от основания жНущка и достигающие поверхности клетки.
На продольном срезе предполагаемого аксостиля^ окру-женнсй^^ййбраной, видны гранулы и полосы. Жгутик содержит центральную аксонему, покрытую оболочкой, В ак-еонеме имеется 10 пар фибрилл, из которых 9 расположены йя| периферии и одна в центре.
<jr Урогеиитальные трихомонады обладают выраженной подвижностью. Движения они довершают вокруг продольной оси тела, которые могут быть поступательными и вращательными; иногда трихомонады бывают малоподвижными. Однако в этих случаях у них сохраняется деятельность жгутиков и отчетливо видны колебания волнообразной перепонки. Некоторые авторы (Г. М. Ткаченко, 1955; Н. Е. Сидоров с соавторами, 1959; Wenrich, 1947, и др.) отмечают у трихомонад наличие амебоидных движений. По данным В. И. Рыбникова (1964), трихомонады могут существовать в подвижной и неподвижной формах. По его мнению, неподвижные трихомонады — это адаптивные формы, появляющиеся при изменении условий, обычных для данного протиета в естественмй среде его обитания. Появлению неподвижных трихомонад способствуют повышение реактивных сил макроорганизма, накопление в сре де обитания продуктов жизнедеятельности паразита, а так же проводимое лечение. В частности, оrруглые неподвижные
формы трихомонад автор наблюдал при назначе-нии больным различных методов.лечения: рентгено- и радиотерапии, химиотерапии кислотами,.щелочами и солями тяжелых металлов. Эти формы протистов якобы бблее устойчивы к различным препаратам (в условиях макроорганизма). Автор считает, что наиболее часто неподвижные трихомонады обнаруживаются у девочек и мужчин. Неподвижных или малоподвижных трихомонад обнаруживали у мужчин и другие авторы (Б. Ф. Печерский, 1951; В. П. Ме-тальников с соавторами, 1959, и др.)- Хотя эти протисты в мужской уретре оказались менее подвижными, чем во влагалище, А. Ф. Аникин (1964), однако, считает, что совершенно неподвижных трихомонад не бывает ни в выделениях, ни в культурах. С этим положением согласуются и "результаты наших наблюдений.
Некоторые авторы (А. А, Васильев-Чеботарев, 1927; С. Г. Юрьевский, 1931; И. А. Евнин, Н. А. Лапидус, 1939, и др.) указывают, что присутствие крови (даже в неболь шо количестве) во влагадащном секрете значительно по вышает активность трихомонад. В противоположность это му активность паразитов при низких температурах, сог ласно литературным данным и нашим наблюдениям (1960, 1964, 1965), резко понижается.
Питаются трихомонады путем всасывания через поверх ность тела питательных веществ из окружающей среды, т. е. эндосмотически. Данное положение подтверждается тем, что в естественных условиях в протоплазме протистов лишь в исключительных случаях имеются пищеварительные вакуоли (Jirovec, 1960). Некоторые авторы (В. Г. По-ножина, 1923; Б. С. Пойзнер и А. А. Кулакова, 1952, 1959, и др.) допускают возможность заглатывания трихомонада-ми оформленных частиц (распавшихся клеток, бактерий, эритроцитов, грибков и т. п.) через рот, наличие которого у них, как указывалось выше, не доказано. На факт проникновения в цитоплазму протиста различных включений (гонококки, сперматозоиды и т. д.) указывают также А. П. Колесов (1950), П. В. Журавский (1953), А. М. Кор-чемкин (1956, 1960) и др. Проникновение форменных элементов (клеточный детрит, бактерии и т. п.) в протоплазму трихомонады, однако, происходит, по их мнению, по типу фагоцитоза, а не путем заглатывания через рот. Согласно данным Schmidt-Gross (1958), пользовавшегося методом фазово-контрастной микроскопии, место прсщйкновещя бактерий в протоплазму паразита находится у ясноващш прикрепления жгутиков. В дальнейшем благодаря образному интенсивному движению ундулирующей браны бактерии продвигаются в глубь тела где и распадаются на зернышки. Во второй фазе пищеварения в задней части тела появляются полущаршиЛШе выпячивания протоплазмы, содержащие много вакуолей, наполненных зернышками. По^ле выделения зерныщек урогенитальная трихомонада снова приобретает присущую ей продолговатую форму.
Между тем далёко не все авторы (Jirovec, 1958;И. И. Ильин, 1962, и др.) считают трихомонад способными
фагоцитировать сперматозоиды. Некоторыми из них, на
пример, было подмечено, что при трихомонозе имеются яв
ления токсикоза сперматозоидов, а не их фагоцитоза (Ну-
nie с соавторами, 1960). Подобно этому Б. А. Теохаров
(1957, 1960) также наблюдал совершенно иные взаимоот
ношения между указанными компонентами^ В частности,
сперматозоиды, смешанные in vitro с трихомонадными ли-
затами, уже в первые часы становились малоактивными, а
спустя 5—12 часоЕп— совсем неподвижными. Снижение ак
тивности сперматозоидов наступает, очевидно*вследствие
обеднения спермы фруктозой, так как количество послед
ней, по данным Keutel (1958), заметно уменьшается в
случае обнаружения в сперме большого числа три-
хомонад. ч
Детальное изучение пищевой потребности урогениталь-ных трихомонад стало возможным благодаря исследованиям Johnson (1940), Trussell (1940—194*) и Щ. зХ. Тераса (1955), которым удалось получить этих простейших в без-бактёриальных культурах. Использование метода культур позволило установить, что для обеспечения роста и нормальной жизнедеятельности урогенитальных трихомонад крайне важное значение имеют полимеры глюкозы (с конфигурацией глюкозо-4,1-р-глюкозид), белки сыворотки, пантотенат кальция и некоторые другие вещества (Кцр-ferberg с соавторами, 1948; Read, 1957; Carneri, 1956; О. Ировец, 1960, и др.). За один час урогенитальная трихомонада потребляет 160*10~8 мл кислорода, а 100 млн. этих протистов за указанное время выделяют 85 мл углекислоты (Kupferberg с соавторами, 1953). Кроме того, паразит in vitro выделяет молочную и в небольшом количестве пировиноградную кислоты, изменяя рН среды. При
22
урогенитальных трихомонад на питательных средах они образуют кислоту из всех Сахаров. А. Лаан, 1961). Интенсивность кислотообразующей функции зависит в этом случае от популяции простейших и вида сахара. Наибольшее количество кислоты выделяет на питательных средах, содержащих мальтозу и крах-. В первый день роста культуры кислота почти не обра-|г^уется, на зто^ые сутки количество ее резко увеличивается Достигает максимума во время самого интенсивного роста протистов. С уменьшением числа подвижных трихомонад падает и интенсивность образования кислоты. Полное прекращение ее образования наступает с прекращением движения трихомонад. Результаты наших исследований также показали, что с возрастом культур реакция среды становится кислой (рН смещается от 6,3 до 4,0). Имеются основания полагать, что в естественной среде обитания трихомонады образуют кислоту за счет гликогена слизистой влагалища.
В питательных средах урогенитальные трихомонады разлагают с образованием кислоты и газа мальтозу,,глюкозу, крахмал и слабо разлагают лактозу, галактозу и другие углеводы (Б. А. Теохаров, 1957, 1959; Ю. X. Терас, 1961, и др.), т. ё. являются биохимически активными микроорганизмами. Если отдельные штаммы трихомонад почти не отличаются друг от друга по степени выраженности у них кислотообразующей функции, то по интенсивности газообразования их можно разделить на три группы (Ю. X. Терас, 1961).
Первая группа штаммов протистов способна продуцировать газ почти из всех Сахаров (арабинозы, ксилозы, рам-нозы, глюкозы, галактозы, мальтозы, лактозы, сахарозы, рафинозы, крахмала, глицерина, маннита, дульцита, инозита). Вторая группа разлагает с образованием газа только мальтозу и маннит. Среди штаммов трихомонад третьей группы подобной закономерности в отношении ферментации Сахаров не наблюдается.
Из протеолитических ферментов во внешнюю среду паразит выделяет в небольшом количестве гиалуронидазу (Н. И. Севастьянова, 1961; Boni, Orsi, 1958; Filadoro, Orsi, 1960).
Примерно 3*109 простейших способны образовать 160 вискозиметрических единиц гиалуронидазы. В незначительной степени урогенитальные трихомонады обладают также амилазной (Е. Л. Рыбалко, 1957), каталазной, фосфатаз-
23
ной и другими видами активности (Н. И. Севастьянова, 1961; Sharma, Bourne, 1964, и др.).
Количество белкового азота в культурах^ мало изменяющееся в первые 2—3 дня сс момента посева трнхомонад^ т. е. в период интенсивного их размножения, заметно^ уменьшается в последующие дни роста паразитов. Наоборот, процент содержания остаточного азота в культуре постепенно увеличивается. Содержание аминокислот и аммиака также в первые 2—3 дня увеличивается немного, а в последующем периоде роста — значительно. Более активный распад белков, происходящий в поздней фазе культивирования трихомонад, зависит, вероятно, от ферментов, освобождающихся при гибели паразитов (Jyori, 1957).
Урогенитальная трихомонада — факультативный анаэроб. Лучше, однако, паразит растет на питательных средах в атмосфере азота (Ivey, 1961).
Данные литературы и результаты наших исследований (И. К- Падченко, 1961) свидетельствуют о том, что рН среды от 3,4 до 8,3 в культурах трихомонад не является препятствием для их развития. Между тем оптимум роста паразитов „находится при рН от 4,8 до 7,6 (Б. А. Теохаров, 1958; О. Ировец, 1960, и др.).
В естественной среде обитания трихомонад обнаруживают обычно при рН влагалищного секрета от 4,5 до 8,5. Иногда их находили у беременных при рН 3,6—4,7 (Nieo-lettr, ~1961). Однако максимально благоприятные условия для них создаются в выделениях, имеющих кислотность от 5,6 до 7,0 (А. Г. Кан и О. С. Таргонская, 1938; Е. Л. Рыбалко, 1956; В. А. Сапожкова, 1956, 1960; Б. А. Терхаров, 1959; Roth, 1944, и др.).
В естественных условиях трихомонады питаются, как это считают некоторые авторы (Е. И. Кватер, 1935; Б. В. Вер-шинский, 1958, и др.), гликогеном, содержащимся в мочеполовых органах человека (влагалище, предстательная железа и др.).
Вопросы, касающиеся цикла развития и размножения урогенитальных трихомонад, освещены в отечественной и зарубежной литературе противоречиво. В частности, по мнению В. Г. Поножиной (1923), С. Л. Козина (1957, 1961) и др., в цикле развития урогенитальной трихомонады следует различать вегетативную форму и стадию цисты. Исходят при этом авторы из того факта, что в половой сфере, особенно у мужчин, нередко встречаются крупные, круглой формы, лишенные жгутиков образования, якобы напомина-
24
ющие по своей толщине и устойчивости стадию цисты. Подобные образования Г. М. Ткаченко (1955) считал самостоятельной стадией в жизненном цикле трихомонады и назвал ее цистоидной формой. По его мнению, трихомона-да существует в трех формах: вегетативной, амебоидной и цистоидной.
При культивировании урогенитальных трихомонад на питательных средах, содержавших в различных концентрациях осарсол, Ю. X. Терас (1954, 1958) наблюдал единичные крупные неподвижные образования, лишенные жгутиков. Подобно этому наблюдал он их и в серозном экссудате, взятом из брюшной полости экспериментально' зараженных урогенитальными трихомонадами мышей. Автор допускает, что это были более «стойкие» формы развития трихомонад, так как они всегда давали на питательных средах рост культуры.
В противоположность этим взглядам некоторые авторы (Б. А. Теохаров, 1959; Л. Е. Чалая, 1959; О. Ировец, 1958, 1960; Р. Петер, 1955, 1959, и др.) категорически отрицают наличие стадии цисты в жизненном цикле урогенитальной трихомонады. По мнению этих авторов, обнаруживаемые в содержимом половых органов и на питательных средах круглые, неподвижные, безжгутиковые образования, не превышающие по своим размерам грушевидных трихомонад, являются дегенерирующими формами паразита. Подтверждается это тем, что указанные образования не дают роста на питательных средах, а их оболочка ничего общего не имеет с цистной оболочкой простейших. Проводившееся нами в течение ряда лет (1957—1965) изучение биологических свойств трихомонад в различных условиях внешней среды позволило установить, что такие протисты появляются в культурах в процессе их старения. В частности, с возрастом культур все трихомонады постепенно округляются, теряют жгутики, многие из них несколько уменьшаются в размерах и оседают на дно пробирки. Протоплазма протистов становится вакуолисто-зернистой. Заканчивается этот процесс обычно лизисом трихомонадных клеток.
Некоторые авторы (И. Лазаревич, 1870; П. В. Толстов, 1950) в прошлом допускали, что размножение урогенитальной трихомонады происходит половым путем. В настоящее время, однако, считается установленным, что этот протист размножается исключительно путем деления, которое может быть равномерным и неравномерным. Неравномерное деление трихомопады, или так называемое почко-
25
ванне, было описано И. П. Лазаревичем (1870), П. В. Жу-равским (1956), Л. Г. Богачевой (1958) и др. Отделяемая при почковании дочерняя особь всегда значительно меньше по своим размерам материнской клетки.
В случае равномерного деления паразита на две особи процесс размножения начинается с последовательного продольного разделения блефаропластов, ядра, жгутиков и волнообразной перепонки, количество которых удваивается, и заканчивается поперечной перешнуровкой протоплазмы (К. А. Карпова, 1941; Е. II. Павловский, 1946; С. Л. Козин, 1959, 1961). После разделения протоплазмы дочерние особи отделяются друг от друга. Полный цикл деления продолжается в среднем от 30 минут до 1*/2—2 часов.
Наряду с простым делением протиста на две особи некоторые авторы описали также множественный способ его размножения. Согласно данным С. Л. Козина (1959, 1961), в случае такого размножения трихомонада делится на 4— 8 особей. Крупные паразиты диаметром до 40—60 мк, содержавшие от 3 до 7 блефаропластов, из которых исходили интенсивно двигавшиеся жгутики, были выявлены в культурах и описаны И. А. Лааном (1961). При множественном размножении трихомонад первоначально происходит деление ядра, блефаропласта, аксостиля и жгутиков. В меньшей мере это касается цитоплазмы, где возникает только перетяжка. Разделение цитоплазмы происходит в более позднем периоде развития протиста и лишь после этого образуются самостоятельные особи (С. Л. Козин, 1959, 1961; Jimenez, 1962). На возможность множественного деления трихомонад, кроме того, указывают В. Г. Поножина (1923), В. А. Догель (1939), И. А. Евнин и Н. А. Лапидус (1939) и др. Хотя известный паразитолог О. Ировец (1948, 1958) категорически и не отвергает возможность множественного размножения у трихомонад, однако он пишет, что ему никогда не удавалось наблюдать более чем трехъядер-ные формы паразита.
Результаты наших исследований (1960, 1964) показали, что урогенитальные трихомонады размножаются на свежей, свободной от дополнительных примесей питательной среде ТУ (Ю. X. Терас, 1955) обычно путем продольного деления клетки на две особи (рис. 5, 6). Лишь иногда (при загрязнении пробирок) в первые 2—3 суток роста культуры встречаются отдельные паразиты, делящиеся на 4—8 особей. Трихомонады с множественным типом деления округлые, крупные (диаметр 18—40 мк), вяло подвижные (рис. 7, 8,
27
9). По своему характеру эти движения чаще не поступз тельные, а вращательно-качательные. Следует что даже в случае кажущейся абсолютной
У таких протистов обычно наблюдается более или мрнрр активная деятельность жгутиков. Имеются поэтому основа ния согласиться с мнением А. Ф. Аникина (1964?У что То вершенно неподвижных жизнеспособных трихомонад ^ бывает ни в выделениях из половых органов ни в культу рах. Разделение крупной трихомонады на дочернее особи может произойти одномоментно или последовательно Пои последовательном способе деления протиста протоплазмя материнской клетки распределяется между дочерними осо 1ми чаще неравномерно. Это обстоятельство позволили hp которым авторам (И Лазаревич, 1870; П В ЖурТскиГ 1956, Л. Г. Богачева, 1958, и др.) предположить и описать
в жизненном цикле урогенитальной трихомонады особый способ размножения, названный ими «почкованием».
Нам стало также известно, что появление в культурах значительного числа трихомонад с множественным типом деления происходит в случае воздействия на них некоторыми химическими и лекарственными веществами. Такая стимуляция множественного размножения протистов связана не с произвольным разведением того или другого вещества в питательной среде, а со строго определенными для каждого из них концентрациями, соответствующими сублетальным дозам в отношении этих паразитов (табл. 1). Активизируется множественное размножение протистов также в случае воздействия на них человеческой мочой, высокими
и низкими температурами.
Таблица 1
Уровень сублетальной концентрации некоторых веществ, обусловливающий множественное размножение урогенитальных
трихомонад
Название вещества
Концентрация вещества
в питательной среле
(в %)
рН питательной среды при наличии в ней сублетальной концентрации вещества
Винный спирт
Борная кислота
Хлорная известь
Осарсол
Формалин
Уротропин
Нашатырный спирт
Бензойная кислота
Едкий натр
Лизол
Соляная кислота
9,0—10,0
0,5—1,0
0,9—1,0
0,09—0,1
0,05—ОД
0,05—0,1
0,09—0,1
0,09—0,1
0,09—0,1
0,05—0,1
0,05—0,1
5,44
4,96
10,65
3,55
5,51—5,54 7,73—8,87
10,78 2,87
10,72
9,38—9,95 1,0
До настоящего времени природа фактора многоделения, как предполагаем мы его называть, стимулирующего множественное размножение трихомонад, остается пока не выясненной. Можно лишь предполагать, что своим происхождением он обязан массовой гибели паразитов, вызываемой многими агентами внешней среды, в том числе сублетальными дозами химических веществ. Возможно, что именно погибшие паразиты являются источником возникновения фактора многоделения. Зависимость же его происхождения от концентрации водородных ионов исключается, подтверждением чему являются довольно широкие пределы колебаний рН среды (от 1,0 до 10,7), при которых в культурах по-
30
являлись паразиты с множественным типом деления (см. табл. 1).
Мы считаем, что по своей биологической сущности процесс множественного размножения урогенитальных трихо-монад, очевидно, является одной из форм защиты и сохранения данного паразита как биологического вида от вредных воздействий окружающей среды. Убеждают нас в этом не только результаты собственных исследований, но и наблюдения В. И. Рыбникова (1964), согласно которым малоподвижные трихомонады появлялись также в естественной среде обитания (мочеполовые органы) в случае назначения больным радио-, рентгено- и химиотерапии, т. е. при воздействии на трихомонад явно неблаготворных факторов.
При многократном тщательном исследовании крупных трихомонад с множественным типом деления как в натив-ных мазках, так и на постоянных препаратах, окрашенных азур-эозином по Романовскому или гематоксилином по Гей-денгайну, нам никогда не приходилось наблюдать у них оболочки, хотя бы в какой-то мере напоминающей оболочку цисты амеб, лямблий или других простейших, имеющих в своем жизненном цикле эту стадию. Следовательно, результаты наших исследований согласуются с данными тех авторов (Б. А. Теохаров, 1959; Л. Е. Чалая, 1959; О. Иро-вец, 1948, 1958, 1960; Р. Петер, 1958, 1959, и др.), которые отрицают наличие стадии цисты в цикле развития урогени-тальной трихомонады. Возможно, что именно крупные трихомонады с множественным типом деления некоторые авторы принимали за цисты (В. Г. Поножина, 1923; С. Л. Козин, 1957, 1960, 1961, и др.), цистоидные формы (Г. М. Тка-ченко, 1955; или за так называемые устойчивые формы (Ю. X. Терас, 1954, 1959).
Такое допущение окажется тем более правомерным, если учесть, что крупные, круглые безжгутиковые, малоподвижные образования, т. е. формы паразитов, принимавшиеся этими авторами за стадию цисты или «цистоидную форму», обычно обнаруживаются, как было показано выше, у муж-чип. Более частые находки подобных образований в выделениях из мужской уретры, по нашему мнению, объясняются тем, что урогенителыше трихомонады в этом случае постоянно омываются мочой, а следовательно, систематически подвергаются воздействию фактора многоделения, стимулирующего множественное размножение трихомонад. Подобно этому протесты оказываются, как это следует из результатов наших наблюдений (см. табл. 1), под влиянием
31
фактора многоделения также в случае воздействия на них субтоксическими дозами осарсола. Поэтому мы считаем, что именно данный факт был положен в основу появившегося в литературе утверждения, согласно которому в цикле развития урогенитальных трихомонад якобы существуют так называемые устойчивые формы.
Итак, приведенные данные позволяют заключить, что урогенитальные трихомонады существуют лишь в вегетативной стадии. Основной биологической формой существования протеста является грушевидная, тогда как другие разновидности его форм являются промежуточной стадией развития, появляющейся только в процессе размножения. Промежуточную и далеко не постоянную стадию в жизненном цикле паразита представляют также крупные (до 40— 60 мк) и карликовые трихомонады. Те и другие появляются лишь в процессе размножения. Карликовые возникают вследствие неравномерного распределения протоплазмы между дочерними особями. Крупные трихомонады представляют собой паразитов, находящихся в стадии множественного деления, возникающего только в неблагоприятных условиях (при воздействии сублетальными дозами химических веществ, лучами радия, рентгеновыми лучами и т. д.). Трихомонады обладают подвижностью, питаются эндосмо-тически и путем фагоцитоза, во внешнюю среду они выделяют кислоту, газ и протеолитические ферменты (гиалуро-нидаза, амилаза и каталаза). В биохимическом отношении эти протесты являются активными микроорганизмами.
Не приходится сомневаться, что биологические свойства урогенитальных трихомонад, определяющие некоторые морфологические, физиологические и биохимические их особенности, должны учитываться прежде всего при решении вопросов, связанных с диагностикой трихомонадной инвазии. Становится также очевидным, что в связи с отсутствием в жизненном цикле урогенитальной трихомонады стадии цисты единственным возбудителем при данной инфекции может быть лишь вегетативная форма протиста. В этом свете ее устойчивость в условиях внешней среды приобретает особое значение.
УСТОЙЧИВОСТЬ ТРИХОМОНАД В УСЛОВИЯХ
ВНЕШНЕЙ СРЕДЫ
Исследованиями последних лет доказано, что обязательным условием жизнедеятельности урогенитальных трихомонад является наличие влаги. Различные факторы внеш-
32
ней среды, способствующие быстрому высыханию материала, содержащего трихомонады (повышение температуры, снижение влажности и т. д.), губительно действуют на паразитов. Например, в условиях 50% влажности трихомонады выживают в течение 12—20 часов (Е. Ин, 1956). Подобно большинству патогенных бактерий трихомонады хуже переносят высокие температуры. Так, если при комнатной температуре протесты оставались жизнеспособными в течение 10—60 минут (Б. А. Теохаров, 1958, 1959; Е. Ин, 1956; Preisler, 1960) и даже до 5 суток (И. А. Евнин, Н. А. Лапидус, 1939), то при более высоких температурах они погибают почти моментально. Напротив, к низким температурам трихомонады менее чувствительны. Оказалось, например, что при —10° они оставались жизнеспособными в течение 7 часов (Е. Ин, 1956). В случае замораживания протистов некоторые авторы (Lindgren, Ivey, 1964) сохраняли их при —43° в течение 8 недель, а при —79° — даже в течение 26 месяцев (McEntergart, 1959). В замороженной культуре тканей возможно хранение культуры трихомонад до 1 месяца. Количество выживающих в этом случае паразитов колеблется от 50 до 100%! (Christian, 1963). Через 1 год в таких условиях живыми оставался 1% паразитов. После оттаивания замороженного материала был отмечен. рост трихомонад, которые морфологически ничем не отличались от исходных, т. е. не подвергавшихся замораживанию при столь низкой температуре. В том же случае, когда протистов трижды подвергали замораживанию при —70° и оттаиванию при 37°, обычно наступало их полное разрушение (Mandel, Honigberg, 1964).
Прямые солнечные лучи убивают урогенитальных трихомонад через 45—60 минут (О. Ировец, 1960).
В пресной воде различных водоемов (реки, озера, пруды, бассейны и др.) и водоисточников (водопроводная) урогенитальные трихомонады выживали, по данным некоторых авторов (Wu, 1938; С. А. Паевский, 1956; В. И. Карелина, 1958, 1959; Л. К. Зерчанинов, 1961), в среднем в течение 3—12 часов. В отечественной и зарубежной литературе имеются и другие показатели устойчивости протистов в воде. Так, согласно данным Г. М. Ткаченко (1955), Б. А. Теохарова (1958, 1959, 1960), Ю. X. Тераса (1961, 1964), О. Ировца (1948, 1958, 1960) и Preisler (1960), трихомонады оставались жизнеспособными в пресной воде не свыше 9—60 минут. В комочках же вагинальной слизи они выживали в течение 12, а иногда даже 24 часов (Л. Е. Ча лая, 1959). В дистиллированной воде протисты были жизнеспособными всего лишь 6—10 минут (Б. А. Теохаров, 1959). Согласно нашим данным (1961, 1962, 1964), в речной и водопроводной воде (Днепр, Лыбедь) урогениталь-ные трихомонады в среднем выживали до 2—5 часов, а в прудовой (окрестности Киева, с. Глеваха, ст. Боярка)—в некоторых случаях даже до 2—8 часов. Следует, однако, отметить, что в ряде опытов в прудовой, речной и водопроводной воде паразиты оставались жизнеспособными не бо лее 2 часов. Трихомонады, внесенные в экспериментальных условиях в воду, полностью лишенную солей или с низким их содержанием (гипотоническая среда), уже в первые минуты обычно разбухают, а их протоплазма ва-куолизируется (рис. 10). Не более 2 часов протисты выживали в речной воде, также по данным Morlunghi, Ricciardi (1961).
В ходе проведения экспериментальных исследований нам стало известно, что сроки выживаемости урогенитальных трихомонад в воде зависят от ее химического состава (табл. 2). Наши исследования показывают, что из всех солей, входящих в состав минеральных вод, применяемых для ванн в условиях курортов Западных областей УССР,
33
29
| Вода | Количество основных химических элементов в неразведенной воде (В Г/ Л) | Общее содержание солей в исследуемой воде (в"%) | Концентрация некоторых минералов в исследуемой воде | Температурные условия опыта (в °С) | Сроки выживаемости урогенитальных трихомонад | |
| хлориды (в %) | сульфаты (в %) | |||||
| Минеральная из | Na 1J !,7 | 20—24 | 72 часа | |||
| источника Л° 7' | К 1,7 | 3 | 1,76 | 0,06 | 36—37 | 96 часов |
| (Трускавец) | Са 1,2 | |||||
| Mg 0,9 | 5 | 2,93 | 0,1 | 20—24 | 72 часа | |
| CI 175,2 | 36—37 | 96 часов | ||||
| \ | НС03 0,6 | |||||
| S04 5,8 | ||||||
| H2S 0,046 | ||||||
| Минеральная из | ||||||
| источника Ла I2 | Na 118,7 | |||||
| (Моршин) | К 4,6 | |||||
| Са 0,4 | 3 | 1,61 | 0,22 | 20—24 | 25!/2 часов | |
| Mg 2,4 | 5 | 2,69 | 0,38 | 36—37 | 48 » | |
| CI 177,0 | 7 | 20—24 | 8 » | |||
| НСО3 0,09 | 36—37 | \~f 8 » | ||||
| SO4 24,6 | \s | |||||
| Минеральная из | Na 0,293 | |||||
| источника № 8 | К 0,002 | |||||
| (Трускавец) | Са 0,610 | 0,32 | 0,05 | 0,13 | 20—24 | 8 часов |
| 36—37 | л часа | |||||
| Mg 0,079 | а 1 С* V- СД | |||||
| CI 0,584 | ||||||
| НС03 0,354 | ||||||